
转录组学分析揭示高粱对麦二叉蚜抗性的响应机制
2023-05-06 02:54:10王坚强李慧明王林芳王绘艳王磊闫素仙杨慧勇
山西农业大学学报(自然科学版) 2023年1期
王坚强,李慧明,王林芳,王绘艳,王磊,闫素仙,杨慧勇
(山西农业大学 高粱研究所/高粱育种与种质创新山西省重点实验室,山西 晋中 030600)
麦二叉蚜是高粱生产过程中引起蚜害的主要优势种群之一,在高粱拔节孕穗期达到危害高峰,直接从植株体内吸食营养、分泌大量蜜露污染作物,影响植物的光合作用,进而影响高粱的产量和品质,同时还是多种病毒病的传播媒体[1]。目前,生产上主要通过药剂进行防治,但高粱对大多数药剂(有机磷类、烟碱类)较为敏感,容易产生药害,因此选育抗蚜品种是防治该虫害的有益措施。然而,育种工作中抗蚜种质资源较少且抗蚜机理缺乏深入系统的研究[2-5],使得高粱抗蚜种质资源创制进展缓慢,严重阻碍现代育种技术在高粱抗蚜品种选育过程中的应用。研究表明,当植物受病菌侵害、动物取食等胁迫时,会改变体内基因的表达和酶类的活性等来完成这些信号的感应、传递以及生物学效应的实现[6]。从植物转录组数据着手,获得转录组与生物胁迫之间的关系,进一步揭示基因信息、发掘新的基因及其功能,能够为植物抗逆性育种提供理论指导[7]。植物一般通过趋避性、抗生性或耐害性3种途径抵抗昆虫取食造成的危害,有些植物抗源兼具多种抗虫途径,而不同的抗虫途径由不同的抗性机理调控。因此,解析高粱的抗蚜机制对高粱抗蚜育种工作具有重要的指导意义。刘国庆等研究表明,高粱对蚜虫的抗性遗传比较复杂,不同抗源材料控制抗性的基因位点数不同,且抗性遗传机制由单显性、隐性基因控制或由少数主效基因控制[5]。Zhou等研究结果显示,昆虫取食通常会导致代谢加速并产生生物碱、萜烯和硫代葡萄糖苷等防御代谢产物,表达模式因植物种类而不同[8]。此外,NBS-LRR等抗性基因也常参与到对昆虫取食的抗性反应中[9-11]。杨慧勇等人对293份高粱种质资源进行抗蚜鉴定,发现PI550607在苗期和成株期均对麦二叉蚜表现出一定的抗性,抗性表现稳定[12]。本研究通过对PI550607接蚜前后0 h和12 h样本进行转录组测序,筛选差异基因,并对差异基因的基因功能和代谢途径进行分析,以期明确PI550607的抗蚜机制。旨在从转录组水平对PI550607的抗蚜机理进行研究,揭示高粱抗麦二叉蚜的调控方式,为高粱抗蚜聚合育种提供理论依据。
1 材料和方法
1.1 植株接蚜后表型调查
抗蚜表型鉴定采取苗期鉴定的方法,将高粱品种PI550607和冀蚜2号播种于50穴的育苗盘内,每穴播种5棵,室温21~25 ℃。出苗后2周,从大田易感植株上采集麦二叉蚜,用毛刷轻轻刷到植物叶片上,共接蚜100头。分别于接蚜后7、10、14 d调查每穴材料叶片的受害百分比,以叶片出现红点为麦二叉蚜受害性状表现。
1.2 测序样品准备及测序
对PI550607进行接蚜处理,处理方式如前文所示,以不接蚜PI550607植株作为对照,分别对不接蚜0、12 h以及接蚜后12 h处理组的地上部分进行采样,采样量不低于1 g,每个样品设置3次生物学重复。样品编号及处理方法见表1。采用Trizol(Invirogen)试剂提取叶片总RNA,并通过DNaseI(TaKaRa)消化样品中的DNA。1.2%琼脂糖凝胶电泳检测RNA样品条带,NanoDrop检测RNA样品纯度及质量,由北京百迈客公司利用Illumina Hiseq2500平台进行转录组测序。
表1 RNA⁃seq样品编号及对应处理Table 1 Sample number and corresponding treatment of RNA⁃seq samples
1.3 转录组数据分析
对原始测序数据进行过滤,去除接头序列及低质量的reads,得到clean reads,保证后续数据分析的可靠性。同时对测序数据中的数据量、Q30、GC含量进行统计。使用HISAT2[13]软件将上述过滤后得到的clean reads比对到高粱参考基因组v3.1,比对标准为 SNP≤1。使用 StringTie[14]软件进行转录本的拼装与表达量分析。通过FPKM值反映基因的表达丰度。使用Deseq2_EBseq软件进行差异基因分析,选取FDR<0.01,且Fold Change≥2作为衡量差异表达基因的标准。得到的差异表达基因,通过BLAST软件与nr、Swis⁃sProt、GO、COG进行功能注释与分类,利用GO数据库对差异表达基因进行功能分类,利用KEGG数据库分析差异表达基因的代谢途径。
1.4 实时荧光PCR验证
采用与转录组测序相同的样品材料,随机选取差异表达基因进行qRT-PCR分析,验证转录组测序分析数据的可靠性。通过Primer 5.0软件设计引物,然后由上海生工生物工程股份有限公司进行引物合成,并通过电泳检测引物特异性。验证基因表达相关引物见表2。使用2-ΔΔCT法计算基因的相对表达并转换成log2FC。
表2 基因表达的相关引物Table 2 The related primers of gene expression
1.5 统计分析方法
采用Excel整理数据、进行统计分析并绘制图表,2样本平均值之间差异显著性通过t-test进行分析。
2 结果
2.1 接种蚜虫后植株表型
分别于接种麦二叉蚜后7、10和14 d调查PI550607以及冀蚜 2号表型,接蚜 7、10、14 d PI550607均对麦二叉蚜表现免疫,受害程度为0%,而冀蚜2号对麦二叉蚜表现高感,在接蚜7d后出现蚜虫侵害症状,10 d后与PI550607相比出现明显的受害症状,接蚜14 d完全死亡(表3,图1)。PI550607抗麦二叉蚜抗性表现较好。

图1 蚜虫取食10 d后PI550607和冀蚜2号表型Fig.1 Phenotype of PI550607 and Jiya2 10 days after greenbug in⁃oculation

表3 植株接蚜后表型调查结果Table 3 Phenotype of plants after greenbug inoculation
2.2 测序数据的质量分析
测序结果显示,共得到4 831.68万~5 872.19万条clean reads,测序总数据量均高于6 GB,Q30比例为93.96%~94.64%,GC含量为55.68%~59.62%,各样品均有超过88%的unique reads进入后续分析(表4)。

表4 转录组测序数据质量统计Table 4 Quality statistics of sequencing data
2.3 基因表达数量及比例分析
采用FPKM标准化方法对测序深度及基因长度进行归一化处理,对9个样品的基因表达量进行统计,不同样本基因表达分布规律一致,FPKM值大于0.1的基因数量均达到87.80%以上,不同样本中基因表达量在3~15范围内的基因数量最多,其次为表达量在0.1~1的基因,基因表达量极高(>60)或极低(<0.1)的基因占比较少(表5)。
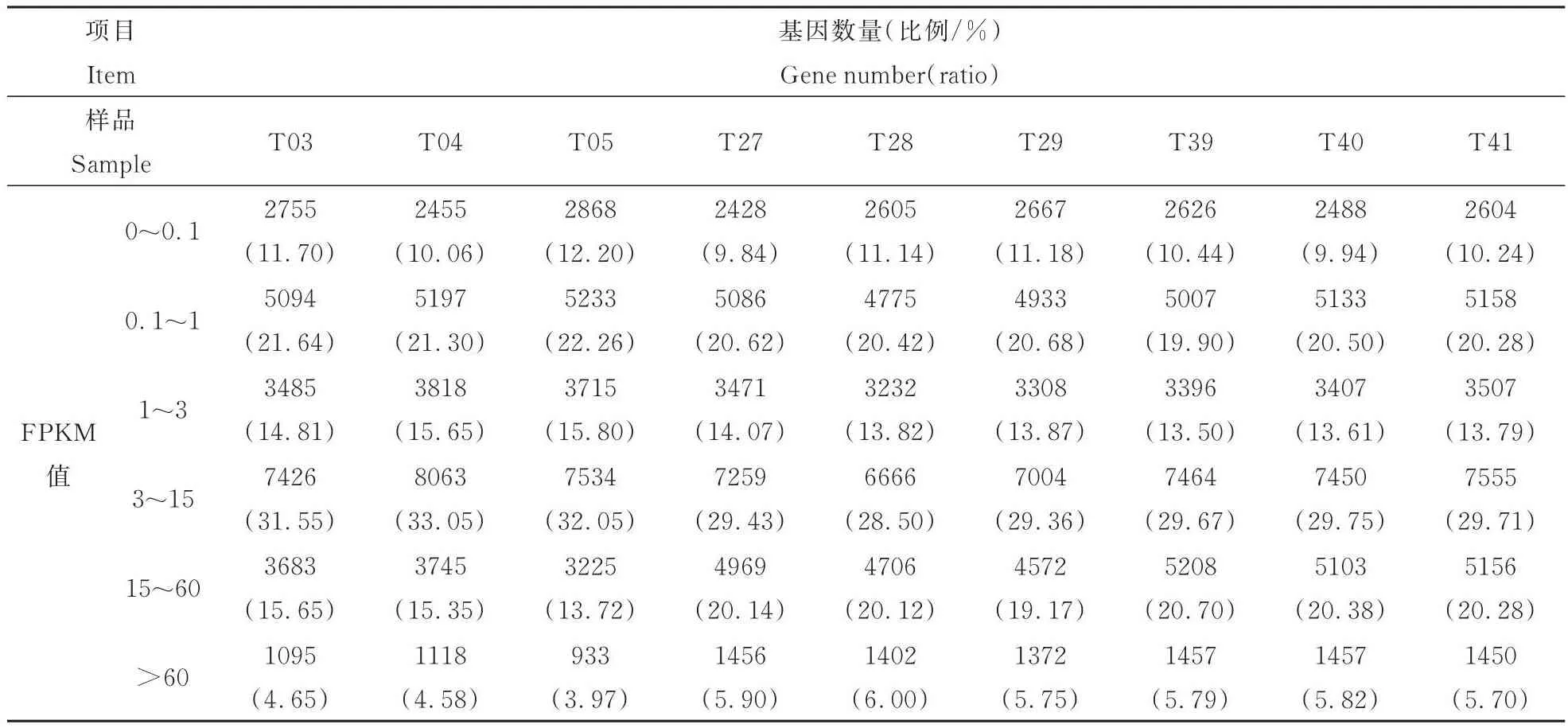
表5 不同表达水平间的基因数量及比例统计Table 5 Statistics of the number and proportion of genes in different expression levels
2.4 差异表达基因的筛选
与接蚜0 h时材料相比,PI550607在接蚜12 h后共有受诱导基因2938个,其中接麦二叉蚜12 h和不接蚜12 h之间表达有差异的基因共222个,排除掉不接蚜12 h与0 h之间差异表达的基因22个,接蚜后12 h特异的受诱导表达基因共200个(图2)。其中上调基因188个,下调基因12个。上调基因数目显著多于下调基因数目,说明高粱品种PI550607中有大量的基因通过增强表达参与到对麦二叉蚜取食的防卫反应中。对差异表达基因进行功能注释,差异表达基因中,参与植物激素代谢途径的有TIFY蛋白基因、茉莉酸氨基合酶基因、丝氨酸/苏氨酸蛋白激酶基因,参与苯丙烷类生物合成的有羟基肉桂酰转移酶基因、肉桂醇脱氢酶基因、过氧化物酶基因、肉桂酰辅酶A还原酶基因、阳离子过氧化物酶基因,参与类黄酮生物合成的基因有查尔酮合酶基因,参与谷胱甘肽代谢的基因有谷胱甘肽-S-转移酶基因,其他数量较多的基因还有WRKY转录因子,细胞色素P450等。

图2 蚜虫取食和对照情况下高粱品种差异基因表达韦恩图Fig.2 Venn diagram of differential gene expression in sorghum cul⁃tivar under greenbug and control treatment
2.5 麦二叉蚜抗性相关差异表达基因的GO分析
对特异性差异表达基因进行GO富集分析发现,在蚜虫取食12 h后处理组与对照间的188个上调表达基因中有178个基因获得注释,分布于35个注释条目中,在生物学过程、细胞组分和分子功能 3个方面所占比例分别为 40.00%(14)、31.43%(11)和28.57%(10)。生物学过程中,参与代谢过程、细胞过程、单个组织过程、生物调节以及对刺激的响应等过程中所包含的基因最多。在细胞组分中,细胞、细胞部件、膜、膜部件以及细胞器功能组拥有的基因数最多,而在分子功能中,参与催化活性和结合的基因占比最多(图3)。这些差异表达基因参与了多个生物过程。
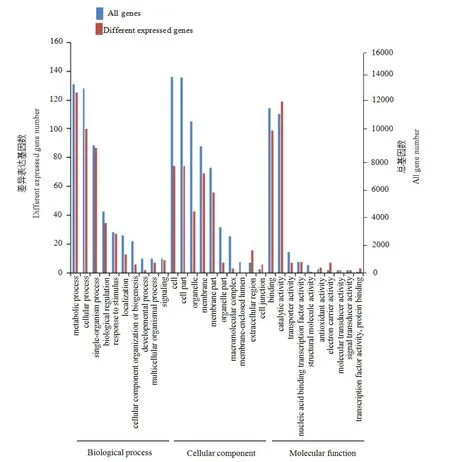
图3 蚜虫取食和对照情况下高粱差异表达基因GO分析柱状图Fig.3 GO histogram of differential gene expression in sorghum cultivar with or without greenbug damage
对富集的GO条目进行筛选(表6),结果显示,差异表达基因在参与细胞壁合成途径的苯丙烷类生物合成过程,响应生物胁迫以及非生物胁迫的创伤反应过程、防卫反应过程、茉莉酸介导的信号途径、对茉莉酸的反应以及催化活性调节、花粉识别、核苷酸为模板的转录活动的负调控、细胞外成分、引导立体定向合成活性、配基查尔酮合成酶活性以及血红素结合等过程中富集程度较高。
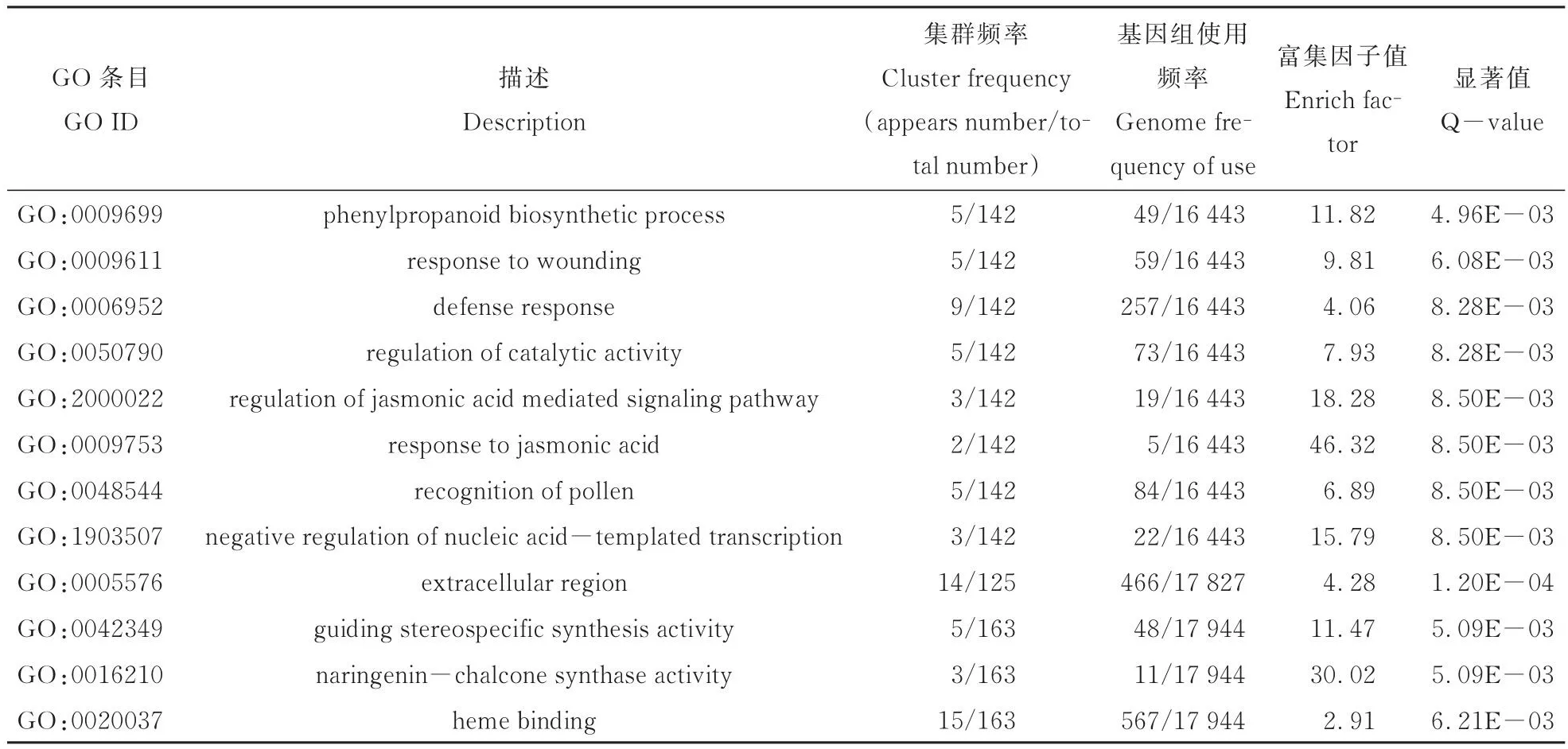
表6 接麦二叉蚜与对照情况下高粱样品间的差异表达基因的GO富集分析Table 6 GO enrichment of differentially expressed genes between sorghum samples with or without greenbug damage
2.6 麦二叉蚜抗性相关差异基因KEGG富集及代谢途径分析
KEGG富集分析显示(图4),特异性差异表达基因都大量富集于苯丙烷类生物合成途径、植物激素信号传导途径以及谷胱甘肽代谢途径、类黄酮代谢合成途径、α-亚麻酸代谢途径、氨基酸生物合成以及植物昼夜节律调控等代谢途径中。

图4 蚜虫取食和对照情况下高粱差异表达基因KEGG富集分析Fig.4 KEGG enrichment analysis of differential gene expression in sorghum cultivar with or without greenbug damage
本研究中,参与α-亚麻酸代谢途径的So⁃bic.001G077400、Sobic.001G230100、Sobic.001G 329800和Sobic.006G095600表达均出现上调,而α-亚麻酸经由十六烷或十八烷途径合成茉莉酸[15]。下游茉莉酸信号途径中的Sobic.001G482700、So⁃bic.002G214800、Sobic.003G224100和Sobic.006G056400的表达也出现上调,说明茉莉酸信号途径受蚜虫取食诱导。其次,苯丙烷类代谢途径中的Sobic.002G 391700、Sobic.002G416700、Sobic.003G050300、So⁃bic.004G065600、Sobic.004G141200、Sobic.005G011500、Sobic.006G014700、Sobic.007G076000和Sobic.010G066700表达均出现上调,其下游参与类黄酮代谢途径的Sobic.005G136300、Sobic.005G137000和Sobic.005G137100的表达也出现上调,说明类黄酮途径也参与到对蚜虫取食的防卫反应中。
2.7 应用qRT⁃PCR对测序结果的验证
为了验证RNA-seq数据的可靠性,随机选择了6个表达差异较大的基因:高粱亚油酸9S脂氧合 酶(Sobic.001G125700)、WRKY 转 录 因 子WRKY76(Sobic.002G202700)、酸性内切酶(So⁃bic.003G244600)、锌指蛋白 ZAT11(Sobic.005G2 26100)、辛 奎 卡 因 -2(Sobic.006G007100)以 及Bowman-Birk型胰蛋白酶抑制剂(Sobic.005G215 900)进行qRT-PCR验证。以未经蚜虫处理0h的对照材料FPKM值为1做基准进行比较,qRTPCR结果表明6个基因在蚜虫取食12 h后表达均出现显著上调,通过转录组测序结果和qRT-PCR分析结果比较表明这6个基因的转录组测序结果与qRT-PCR结果的表达趋势一致(图5),表达倍数上略有差异,转录组测序结果准确可信。
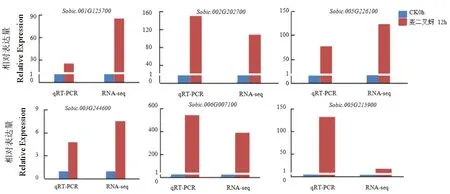
图5 部分基因的qRT-PCR验证Fig.5 qRT-PCR validation of partially selected gene expression
3 讨论
高粱抗蚜反应是一个复杂的过程,本研究对抗蚜材料PI550607在蚜虫取食以及对照条件下的差异表达基因进行研究。在PI550607中,大量的基因参与到对蚜虫取食的抗性反应中,主要通过上调表达这些基因以增强自身抗性,这一结果与Zhang等[11]的研究结果一致,说明在抗虫材料PI550607中主要通过提高基因表达,增强主动的防御措施来抵御蚜虫的取食。蚜虫取食会抑制植物的代谢活动[16-17],能够在昆虫攻击下保持代谢正常的植物往往表现出更强的抵抗力[18]。
在昆虫取食后,参与植物激素代谢与信号途径、创伤反应相关、控制蛋白质和脂质结合、谷胱甘肽代谢和细胞分解代谢过程的基因常参与到植物的防卫反应中[19-20]。抗虫基因型材料通过改变酶功能、信号传导和基因表达以应对蚜虫取食[21-22]。植物激素茉莉酸作为信号分子在对生物和非生物胁迫的抗性中发挥着关键作用。茉莉酸及其衍生物茉莉酸甲酯是植物防御昆虫取食的重要植物激素[23]。本研究中α-亚麻酸代谢途径基因表达出现上调,进而影响到其下游茉莉酸信号途径基因的表达。在特异性差异表达基因中,TIFY蛋白基因、茉莉酸氨基合酶基因、丝氨酸/苏氨酸蛋白激酶基因等基因的表达发生变化均会对植物中茉莉酸信号通路产生影响。这一研究结果说明茉莉酸信号通路在PI550607的抗蚜反应中发挥了重要的作用。茉莉酸和茉莉酸甲酯会诱导植物产生烟碱、多酚氧化酶、蛋白酶抑制素以及大量挥发性有机化合物,对蚜虫产生驱避作用或者引诱蚜虫的寄生性或捕食性天敌[24]。茉莉酸信号通路在抗蚜虫辣椒品种“猪大肠”中特异表达[25],而在易感大豆植株中,蚜虫会抑制植株的茉莉酸信号途径基因表达[26]。蚜虫取食早期茉莉酸的瞬时增加可以阻止蚜虫的在高粱上的定殖,提高对蚜虫取食的抗性[27]。
此外,本研究中参与苯丙酸代谢途径的基因表达出现上调,苯丙氨酸代谢途径是植物体次生代谢途径中的一个通用途径,可生成反式肉桂酸、香豆酸等中间产物并最终可以形成木质素、黄酮、类黄酮等次生代谢产物,参与到水杨酸信号通路以及木质素合成途径中[28]。其中类黄酮代谢途径的基因在PI550607中出现上调,说明在PI550607中苯丙酸代谢途径基因影响了类黄酮的合成继而影响植物对蚜虫取食的抗性。类黄酮类化合物作为植物-微生物间的信号物质、植物着色物质、植物防御素在抗氧化、清除自由基等方面发挥着重要的作用[29-30]。在昆虫取食时,类黄酮作为植物次生代谢产物还会使昆虫体内部分酶活性发生变化,影响昆虫生长发育,使植物对植食昆虫产生防御作用[31]。类黄酮还是合成单宁的前体物质,而单宁也可以保护植物免受伤害,提高植物适应逆境的能力[32]。
4 结论
综上所述,本研究中PI550607苗期对蚜虫的抗性表现为趋避性和抗生性的混合抗性[33]。蚜虫取食后茉莉酸信号途径的上调表达可能对蚜虫产生趋避性,而类黄酮代谢途径、谷胱甘肽代谢途径可能参与到对蚜虫抗生性抗性中。
猜你喜欢
快乐作文(1.2年级)(2023年9期)2023-05-12 11:31:34
古今农业(2022年2期)2022-08-15 01:39:52
小天使·一年级语数英综合(2022年5期)2022-05-25 16:36:27
青年文学家(2022年1期)2022-03-11 12:27:39
发明与创新·小学生(2021年5期)2021-08-23 02:24:00
发明与创新(2021年19期)2021-05-07 07:53:00
文苑(2020年12期)2020-04-13 00:55:10
意林·全彩Color(2019年11期)2019-12-30 06:09:06
意林彩版(2019年11期)2019-11-22 11:49:05
小学生导刊(2018年31期)2018-12-06 08:36:50
