
棉花陆海渐渗杂交群体纤维品质性状的QTL定位
2023-04-06 03:01杨延龙师维军汪鹏龙郑巨云郭仁松胡文冉
新疆农业科学 2023年1期
马 君,杨延龙,师维军,汪鹏龙,郑巨云,郭仁松,胡文冉,杨 洋
(1.新疆农业科学院经济作物研究所, 乌鲁木齐 830091;2.新疆农业科学院库车陆地棉试验站,新疆库车 842000;3.新疆农业科学院核技术生物技术研究所,乌鲁木齐 830091)
0 引 言
【研究意义】陆地棉适应性广且产量高,而海岛棉高抗黄萎病且纤维品质优良,但是产量偏低、生育期长[1]。利用海岛棉优良的纤维品质性状,将其优良基因导入陆地棉,培育出适应性广、产量高且纤维品质优良的陆海渐渗系,是对陆地棉进行优良性状改良的有效方法[2]。在陆地棉高产的基础上对其进行纤维品质的改良也成为当前棉花育种的重要研究方向[3]。棉花的纤维品质是由多基因控制的数量性状,受环境因素影响较大,普遍存在基因型与环境的互作,且与产量构成因素存在负相关等,遗传机制较复杂[4]。常规遗传育种方法在提高陆地棉产量、改良纤维品质的过程中起到了重要作用,培育了大面积推广的优良品种。种内杂交选育中骨干亲本的集中使用和长期的人工选择会导致陆地棉遗传基础狭窄[5-6]。有效发掘利用海岛棉优异性状基因,对拓宽陆地棉栽培种遗传基础有重要意义。【前人研究进展】分子标记技术的快速发展为标记辅助选择和QTL聚合育种提供了条件,可以从目标性状的分子水平上进行选择,有利于提高表型性状选择的准确性,也是提高作物遗传改良及育种进程的有效途径[7]。通过构建连锁遗传图谱,与棉花纤维品质性状相关的QTL(Quantitative trait locus)可以在染色体组上准确定位,通过分子标记辅助选择育种减少常规育种选择的盲目性[2]。目前对陆地棉与海岛棉优势结合进行棉花纤维品质性状QTL定位做了许多研究,构建了连锁遗传图谱,并检测到大量与纤维品质相关的QTL位点[2,8-14]。【本研究切入点】优良陆海渐渗系群体是基因聚合、QTL定位等研究的理想材料,需研究利用与纤维品质紧密连锁的分子标记进行辅助选择育种。【拟解决的关键问题】选用早中熟陆地棉品种新陆中60号为母本与优质抗病海岛棉品种新海41号为父本进行杂交,并以新陆中60号为轮回亲本构建回交群体BC1F2。利用SSR分子标记,对陆海渐渗杂交BC1F2分离群体进行基因型分型,构建陆海杂交的遗传连锁图谱,种植BC1F2家系群体,测定纤维品质性状,利用区间作图法对纤维品质性状进行QTL定位,检测与棉花优质纤维品质性状相关的稳定QTL位点,为海岛棉优良基因定位和利用奠定研究基础。
1 材料与方法
1.1 材 料
2014年在库车陆地棉试验站,利用高纯系陆地棉品种新陆中60号为母本与优质海岛棉品种新海41号为父本杂交,获得F1种子,2015年再以新陆中60号为轮回亲本与F1回交获得BC1F1群体种子。
2018年在库车陆地棉试验站种植新陆中60号、新海41号及BC1F2分离群体,在棉花现蕾期取幼嫩叶片采样后立即用液氮速冻,亲本材料随机各选5株采样,BC1F2群体选取190株采样。获得151份前期已采样的BC1F2群体。
1.2 方 法
1.2.1 纤维相关性状的调查
于2018年10月田间取样,在BC1F2群体中选取已标记过的151个单株收花,轧花后取15~20 g纤维样品送农业农村部棉花品质监督测试中心进行品质检测,检测指标包括纤维长度、纤维强度、马克隆值、整齐度和伸长率。
1.2.2 SSR分子标记检测与PCR 扩增
棉花基因组DNA的提取参照宋国立等[15]改良的CTAB法,其中纯化 DNA的操作步骤略作改进,用紫外分光光度计测定DNA浓度。SSR的分析流程参照李立群等[16]方法,优化体系参照许玉兰等[17]的方法,确定最佳反应体系为15 μL体系,模板DNA(5 ng/μL)1 μL,正反引物各1 μL(浓度为10 μmol/mL),2×PCR mix 7.5 μL(加拿大应用生物材料有限公司(abm Inc.)的2×PCR mix),dd H2O 4.5 μL。反应程序:94℃ 5 min;95℃ 15 min;Tm 15 s;72℃15 min;30个循环;72℃10 min;退火温度(Tm)根据引物进行调节。
1.3 数据处理
统计SSR多态性标记,在BC1F2群体中与母本新陆中60号相同的条带记为A,与父本新海41号相同的条带记为B,具有双亲性状的条带记为H,BC1F2代中有而双亲中都没有的条带记为D,缺失数据记为-。使用Join Map 4.0软件构建SSR标记遗传连锁图谱,连锁的最低LOD值为3.0,最大遗传距离为50 cM,作图函数采用Kosambi函数。采用QTL分析软件Windows QTL Cartographer 2.5,选用复合区间作图(composite interval mapping,CIM)方法,进行纤维相关性状的QTL定位和效应检测,QTL命名方法按照 QTL(q)+性状(英文缩写)+染色体(chr)+QTL个数(数字)表示。采用单标记分析(Ici Mapping)方法分析纤维品质性状相关的定位。
2 结果与分析
2.1 遗传图谱的构建与纤维品质性状的QTL位点
研究表明,共获得105对差异明显的多态性引物,其中有23对引物不能检测出BC1F2代的基因分离,多态性频率为42.17%。构建了一个包含52个标记、14个连锁群的遗传图谱,该图谱总长824 cM,覆盖棉花基因组的18.5%,最长的连锁群为150.3 cM,包含6个标记,最短的为0.3 cM,包含2个标记。图1
2.2 亲本及BC1F2家系纤维品质性状
研究表明,两亲本间各性状差异明显、范围适中,海陆渐渗系性状值(纤维长度, 断裂比强度和伸长率)均高于亲本新陆中60号,而马克隆值和纤维整齐度相对偏低。各项纤维指标在BC1F2家系群体中均呈连续分布,这些性状都受多基因控制的数量性状,其中,纤维长度略超亲本,存在超亲分离。纤维品质各项指标除伸长率外,偏度都小于1,符合典型的数量性状正态分布特征。表1

图1 遗传图谱与纤维品质性状的 QTL 位点Fig.1 Linkage map and QTL location of fiber quality traits

表1 亲本及BC1F2家系纤维品质性状Table 1 Analysis of fiber quality in parent and BC1F2 population
2.3 BC1F2群体纤维品质性状的相关性
研究表明,BC1F2群体纤维品质各性状间存在着复杂的相关性,纤维长度与纤维整齐度呈极显著正相关,相关性系数为0.552;纤维长度与马克隆值呈极显著负相关,相关性系数为-0.534;断裂比强度与纤维整齐度和伸长率呈极显著正相关,相关性系数分别为0.872与0.983;纤维整齐度和伸长率呈极显著正相关,相关性系数为0.908。存在正相关的各纤维品质之间可相互促进提高,存在负相关的各纤维品质之间相互制约不可能同步提高。表2
2.4 纤维品质性状的QTL定位
研究表明,检测到与纤维品质性状相关的QTL位点1个,该位点与纤维长度相关,位于Chr14的32.31 cM处,标记区间位于BNL1905-BNL3502,LOD值为2.0,加性效应-0.96,显性效应0,解释表型变异8.59%;检测到与马克隆值相关的单标记位点1个,引物名称为NAU1119,位于Chr26上,LOD值为3.0,加性效应-0.15,显性效应3.7,解释表型变异8.88%。表2
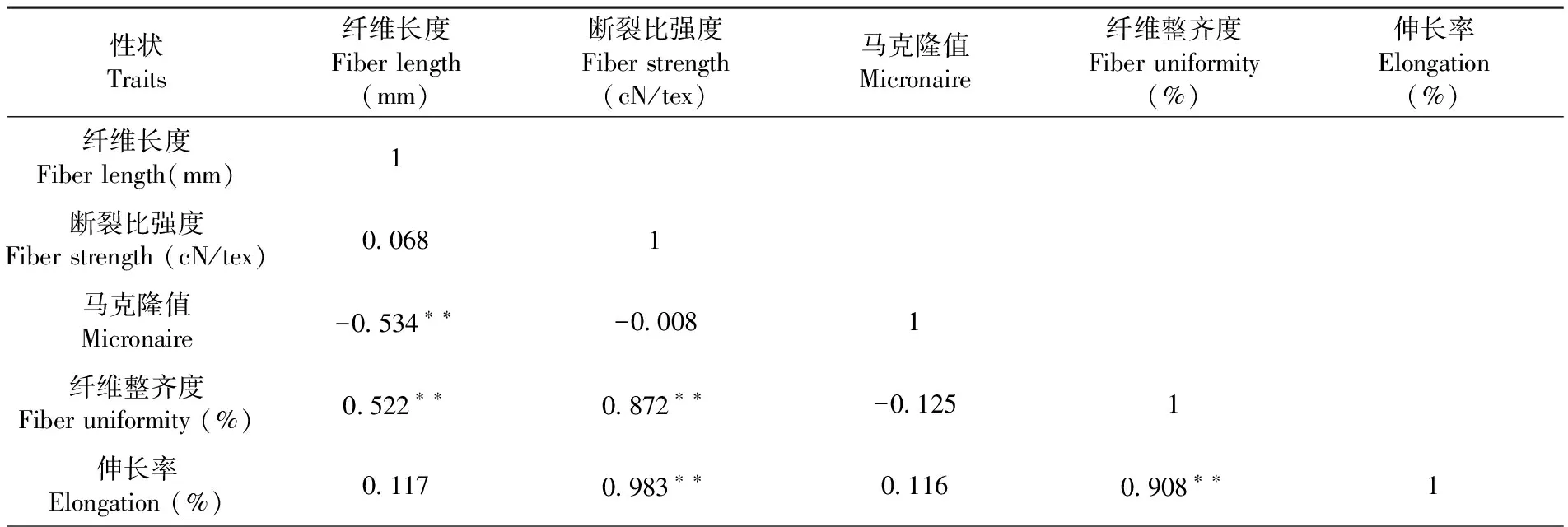
表2 BC1F2群体纤维品质性状相关性Table 2 Analysis of correlation among fiber quality in BC1F2 population
3 讨 论
海岛棉和陆地棉是2个主要的异源四倍体栽培种,在产量、纤维品质和生态适应性等方面存在可互补性[18]。研究选用高纯系主推的陆地棉品种新陆中60号为母本与优质海岛棉品种新海41号为父本进行杂交构建作图群体,将二者优良特性优势结合,实现产量和纤维品质的同步提高。选取249对分布于棉花全基因组的SSR分子标记进行多态性筛选,共获得105对差异明显的多态性引物。当前开发的棉花分子标记引物多来自海陆群体,遗传连锁图谱的加密主要依赖于开发的引物数量和亲本遗传基础的差异大小。棉花海陆群体QTL定位研究趋于饱和。张正圣等[19]利用陆地棉种间杂交(渝棉1号×T586)F2群体,构建了一张包括173个标记位点(159个SSR,11个AFLP和3个形态标记)的遗传连锁图,覆盖1 360 cM,约占棉花基因组的3.06%。Ullao等[20]利用陆地棉种内杂交(MD5678ne-×Prema)F2群体构建的连锁图,包括17个连锁群,81个RFLP 位点,标记间的平均距离8.7 cM,覆盖700.7 cM,约占棉花基因组的15.8%。
因多效现象在自然界生物进化遗传中普遍存在,在多环境多群体中获得的 QTL位点往往有较高的稳定性和精确度。所获得的1个位点在相关文献中已有报道,qFL-14-1与qFM-14-1[21-22]、q FS-14-1[22-24]及q FU-14-1[2]在同一位点,效应不同,该位点对应的等位基因有可能连锁遗传,或者是一因多效[24]。在不同群体不同环境下定位到的QTL,能更准确地反映控制棉花纤维品质性状的等位基因的位置,环境因素对数量性状位点的检测影响很大。
4 结 论
共获得105对差异明显的多态性引物。筛选的与优质性状相关SSR标记可应用于棉花优质分子标记辅助选择上。检测到与纤维长度相关的QTL位点1个,位于Chr14的32.31 cM处,解释表型变异8.59%。检测到与马克隆值相关的位点1个,位于Chr26上,解释表型变异8.88%。
猜你喜欢
区域治理(2022年40期)2022-11-27
少儿科学周刊·儿童版(2022年10期)2022-06-27
小读者(2020年4期)2020-06-16
动漫界·幼教365(小班)(2019年10期)2019-10-28
动漫界·幼教365(大班)(2019年10期)2019-10-28
动漫界·幼教365(中班)(2019年10期)2019-10-28
江苏农业科学(2017年22期)2018-01-06
江苏农业科学(2016年5期)2016-07-23
中国种业(2016年3期)2016-04-14
山东农业科学(2014年1期)2015-03-09
