
铜铁互作效应对方格星虫稚虫生长、存活、体成分及相关酶活性的影响
2023-03-31 07:47曾梦清董兰芳许明珠刘海娟游出超龙子扬林立蔚李世才彭慧婧
西南农业学报 2023年1期
曾梦清,董兰芳,许明珠,刘海娟,游出超,龙子扬,林立蔚,李世才,彭慧婧
(1.广西海洋研究所有限责任公司/广西海水养殖新品种繁育工程技术研究中心,广西 北海 536000;2.广西科学院,南宁 530007)
【研究意义】裸体方格星虫(Sipunculusnudus)俗称沙虫,又称沙肠子、方格星虫,主要栖息在沙质底的沿海滩涂中,在我国广东、广西、海南、福建和山东等沿海地区均有分布,尤其以北部湾海域资源最丰富[1-2]。据报道,方格星虫含有丰富的氨基酸、脂肪酸、多糖及多种矿物质元素,同时含有多种活性物质,具有提高免疫力、抗疲劳和抗血栓等医学功效[3-6]。近年来,野生方格星虫数量锐减,广东和广西两地顺应市场需求,积极开展方格星虫滩涂养殖。为摆脱苗种限制,扩大养殖规模,广西海洋研究所开发了规模化人工育苗技术,经过室内繁育和室外池塘中培,生产了大量苗种。但目前市场上尚无高效的方格星虫专用配合饲料,养殖户多使用各种鱼料或虾料进行池塘中培,不仅成本较高,苗种存活率也受到影响。因此,研发高效的专用人工配合饲料已成为推动方格星虫苗种产业健康发展的重要内容。【前人研究进展】广西海洋研究所研发团队在三大营养元素(糖、蛋白和脂肪)[7-9]及单一微量元素(铜和铁)[10-11]的最适量研究中取得一定进展,已明确方格星虫专用饲料中铜的最适添加量为11.2 mg/kg及铁的最适添加量为115.6 mg/kg时,方格星虫的增重率最大。但各微量元素间存在复杂的相互作用,当饲料中同时添加多种微量元素时,微量元素间可能产生拮抗或促进作用,因此饲料中单一微量元素的最适添加量不能作为多元素饲料中微量元素的最适添加量,需进行多因素互作效应下的最适量探讨。铜和铁均属于生物体内的必须微量元素[12-13],分析其在饲料中互作效应下方格星虫稚虫生长和生理机能的变化情况,对推动方格星虫稚虫人工配合饲料的研发进程具有重要意义。铜与生物体的抗氧化功能紧密相关,是细胞色素氧化酶(CO)、超氧化物歧化酶(SOD)和赖氨酸氧化酶(LOX)等的主要构成成分,通过影响上述氧化酶、拮抗铁和锌等微量元素或有害物质的毒性而参与机体免疫和代谢活动[14-16]。已有研究发现,适量铜对水生动物的生长具有明显促进作用,郭志勋等[17]在基础饲料中添加0~45.0 mg/kg铜饲喂凡纳滨白对虾(Litopenaeusvannamei),结果表明铜添加水平为30.0 mg/kg的对虾生长速度显著高于其他添加水平。但过量铜会抑制动物生长,甚至会使机体产生中毒症状,导致死亡[18]。叶超霞等[19]对斜带石斑鱼(Epinepheluscoioides)的研究发现,饲料铜含量升高到1127.51 mg/kg时,斜带石斑鱼的增重率、饲料效率显著降低。Lee等[20]研究发现,饲料铜含量超过80.0 mg/kg时,斑节对虾(Penaeusmonodon)的存活率显著降低,当饲料铜含量为160.0 mg/kg时,存活率降低至22.22%。铜与铁的转运与吸收关系密切[12],因为铁被吸收后以二价铁离子形式进入血浆,而血浆铜蓝蛋白参与铁的氧化还原过程,因此机体缺铜时血浆铜蓝蛋白不足,可引起对铁元素的吸收和利用障碍。铁在方格星虫体腔液中含量较高[13],是细胞色素酶(CYP)、过氧化氢酶(CAT)、过氧化物酶(POD)的重要组成部分,在氧化还原反应中发挥传递氢的作用,主要参与体内氧的转换、变换和组织呼吸过程,维持生物体的生命活动[21]。由于铁是一种造血元素,因此缺铁时动物体的血红蛋白数量减少,造成各器官供氧减少,导致机体出现缺氧症状[22],如河鲶(Silurusasotus)在缺铁时呈现血球容积比、血红素和红细胞比例下降的现象[23]。不同生物对各种微量元素的需求量不同,且微量元素间存在拮抗或促进的相互作用[12]。刘宝娟[24]研究了铁和铜对皱纹盘鲍(HaliotisdiscushannaiIno)生长的影响,结果表明,在铁含量为1300.0 mg/kg和铜含量为80.0 mg/kg时皱纹盘鲍的生长率最高,铜和铁含量对皱纹盘鲍的增重率具有显著交互作用。【本研究切入点】铜和铁元素属于微量元素中占比较高的元素,且对动物的生理生化功能具有重要作用,但目前关于饲料多因素效应对水生动物影响的研究报道仅见于鱼类和贝类[24-27],有关多种微量元素互作效应对方格星虫生长和生理机能影响的研究较少。【拟解决的关键问题】在方格星虫稚虫饲料的铜元素最适添加量为11.2 mg/kg[10]、铁元素最适添加量为115.6 mg/kg[11]基础上以低量、适量、过量的铜和铁饲料饲喂方格星虫稚虫,探究铜与铁互作效应对方格星虫稚虫生长、存活、体成分及相关酶活性的影响,确定饲料中有机铜和铁的适宜添加量,以期为开发出经济、环保、高效的方格星虫人工配合饲料提供理论依据。
1 材料与方法
1.1 试验材料
根据方格星虫铜和铁单因素试验结果[10-11],分别选择7.8(低量)、13.2(适量)和23.5 mg/kg(过量)铜含量梯度及61.5(低量)、120.6(适量)和243.7 mg/kg(过量)铁含量梯度制作方格星虫稚虫饲料并开展养殖试验。预先制作基础饲料,再用蛋氨酸铜和蛋氨酸铁调节试验饲料中的铜和铁元素含量。基础饲料配方参考许明珠等[28]的方法进行配制,主要由酪蛋白(40%)、糊精(20%)、微晶纤维素(14%)、明胶(10%)、鱼油(8%)、褐藻酸钠(3%)、卵磷脂(2%)组成,再添加1.5%维生素预混料和1.5%复合矿物盐。采用3×3完全随机试验设计,将基础饲料分为9组,3个水平(7.8、13.2和23.5 mg/kg)铜(以蛋氨酸铜形式)和3个水平(61.5、120.6和243.7 mg/kg)铁(以蛋氨酸铁形式)随机组合后添加于基础饲料中,制成9种饲料饲喂方格星虫稚虫。详细添加量如表1所示。试验用方格星虫稚虫为广西海洋研究所有限责任公司自主繁育的苗种。
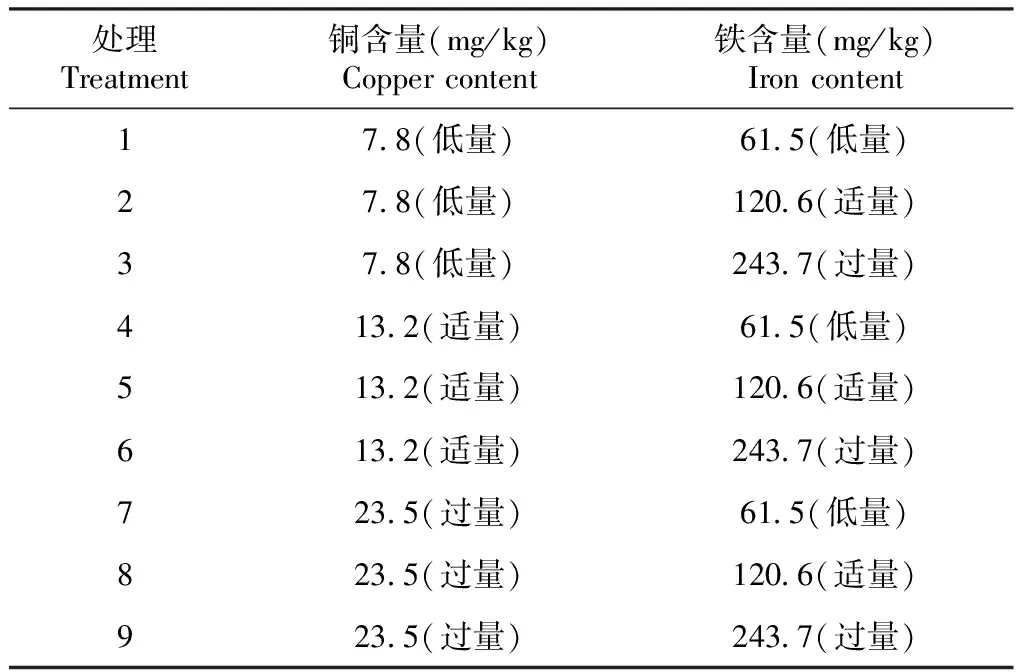
表1 各处理饲料的铜和铁含量
饲料制作方法:先将主要原料粉碎后过100目筛网,然后按照配方逐步称重,使用混合机将原料混合均匀,再与鱼油和水充分混匀,用机器制成过100目筛网的微颗粒饲料并烘干至恒重,冷却后装入密封袋标号冷藏备用。
1.2 养殖方法
养殖试验于2018年8—10月在广西海洋研究所有限责任公司海水增养殖基地进行,为期8周。
挑选自然状态下体长1.3~1.8 cm、平均体质量(16.12±0.26)mg/条的同批次健康稚虫8100条,以300条为1个重复,3个重复为1组,随机分成9组,分别饲喂9种不同水平铜和铁元素的人工微颗粒饲料(处理1~9)。每个重复均使用65 cm×55 cm×45 cm的长方体整理箱作为方格星虫稚虫栖息地,整理箱底部铺设一层5 cm的细砂,加满砂滤海水。每天8:00和17:00换水1/3后投喂饲料,开始时按照放苗总体质量的10%投喂,之后根据饲料残留情况调整,以稍过量投喂为宜。养殖整理箱内布置1个气石,24 h连续充气,保持溶解氧在5.0 mg/L以上。水温保持在27~30 ℃,盐度控制在18‰~25‰,白天光照在3000~8000 lx。
1.3 测定项目及方法
养殖试验结束后,用砂滤海水将砂层中的稚虫翻洗出来,转移至装有砂滤海水的小整理箱中停食排沙24 h,期间连续充气。待稚虫消化道内的沙子完全排出后,称量各处理稚虫总重,同时清点并记录稚虫存活数,用以下公式计算各处理方格星虫稚虫的增重率、特定生长率和存活率。
增重率(%)=(Mt-M0)/M0×100
特定生长率(%/d)=(lnMt-lnM0)/t×100
存活率(%)=Nt/N0×100
式中,N0为初始稚虫条数,Nt为终末稚虫条数,M0为初始平均体质量,Mt为终末平均体质量,t为试验天数。
饲料常规成分和稚虫体成分采用AOAC[29]的方法进行测定。使用105 ℃烘箱将稚虫烘干至恒重测定水分含量,以凯氏定氮法(Kjeltec 8400,瑞典)测定粗蛋白含量,以索氏抽提法(Soxtec 2050,瑞士)测定粗脂肪含量,以550 ℃高温灼烧法测定灰分含量。虫体铜和铁元素含量参照GB 13885—2003《动物饲料中钙、铜、铁、镁、锰、钾、钠和锌含量的测定 原子吸收光谱法》的方法进行测定。方格星虫稚虫虫体(包括体壁和体腔液)匀浆液CAT活性(U/mg)按照南京建成生物工程研究所试剂盒说明进行测定。
1.4 统计分析
试验数据采用SPSS 19.0进行统计和方差分析,以Tukey’s进行多重比较,以Excel 2003绘制折线图,图中折线平行表示无互作效应,相交表示存在互作效应[28]。
2 结果与分析
2.1 饲料中铜与铁互作效应对方格星虫稚虫生长的影响
由表2可知,至养殖试验结束时,不同铜和铁含量饲料对各处理稚虫的终末体质量、增重率、特定生长率和存活率均具有显著影响(P<0.05,下同)。其中,处理2(铜含量为7.8 mg/kg、铁含量为120.6 mg/kg)稚虫的增重率和特定增长率最高;当铁含量为适量(120.6 mg/kg,处理2、5和8)时,改变铜含量其特定生长率无显著差异(P>0.05,下同);当铁含量为低量(61.5 mg/kg,处理1、4和7)时,适量铜和过量铜处理(处理4和7)稚虫的增重率和特定生长率均较高,且均高于低量铜处理(处理1)的增重率和特定生长率;当铁含量为过量(243.7 mg/kg,处理3、6和9)时,方格星虫稚虫的增重率和特定生长率均低于适量铁处理(处理2、5和8),低铜处理3的增重率、特定生长率比适量铜处理6高,但差异不显著,处理3稚虫的存活率与处理6差异显著。
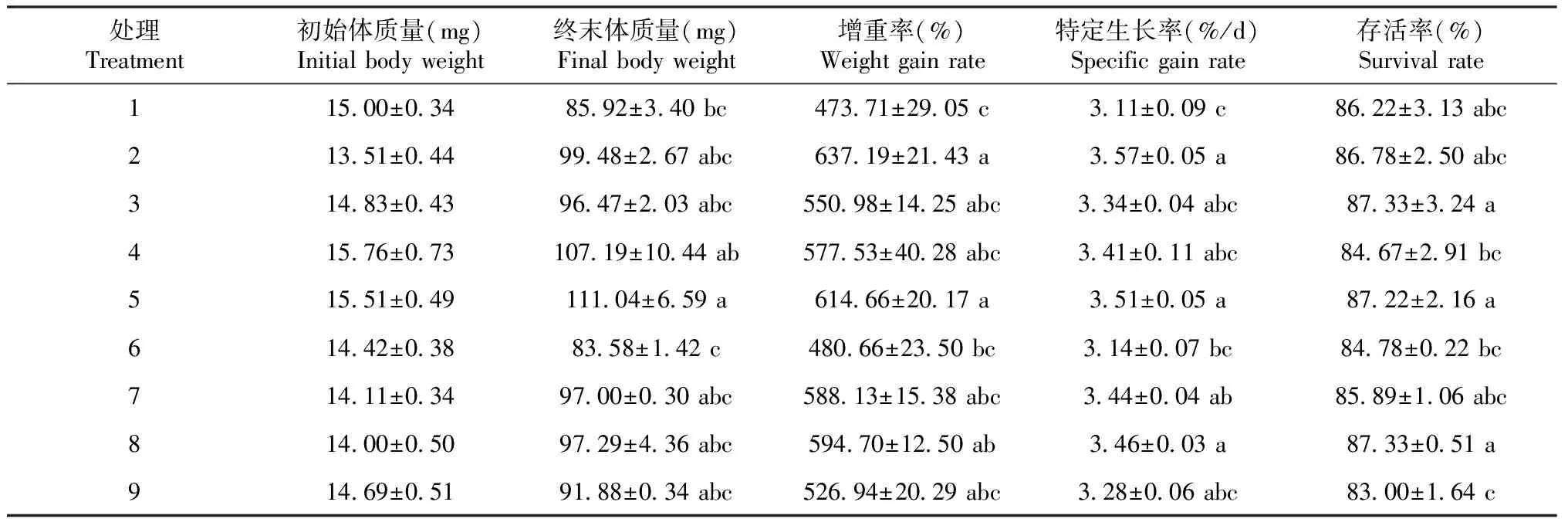
表2 饲料中铜和铁水平对方格星虫稚虫生长的影响
从图1~2可看出,铜含量为7.8 mg/kg、铁含量为120.6 mg/kg时,方格星虫稚虫的增重率和特定生长率最高;当铜含量一定时,增重率和特定生长率随着铁含量的增加呈先升后降的变化趋势。图1~2中折线相交,说明饲料中的铜和铁元素对方格星虫稚虫的增重率和特定生长率呈现明显的互作效应。
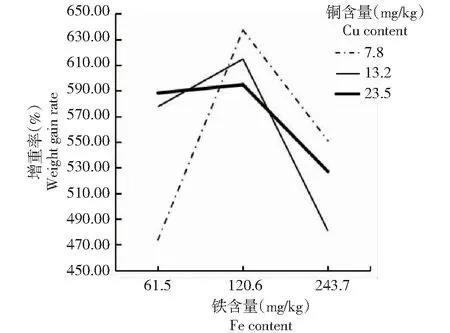
图1 铜与铁互作效应对方格星虫稚虫增重率的影响
图3结果表明,当铜含量为7.8 mg/kg时,方格星虫稚虫的存活率随着铁含量的增加而上升,推测此时存活率主要受铁含量的影响;当铜含量增加到13.2或23.5 mg/kg时,存活率随着铁含量的增加呈先升后降的变化趋势;当铁含量偏低(61.5 mg/kg)时,铜含量13.2 mg/kg处理(处理4)的存活率最低,但当铁含量偏高(243.7 mg/kg)时,铜含量23.5 mg/kg处理(处理9)的存活率最低;图3中折线相交,说明饲料中铜和铁元素对方格星虫稚虫的存活率呈现明显的互作效应。
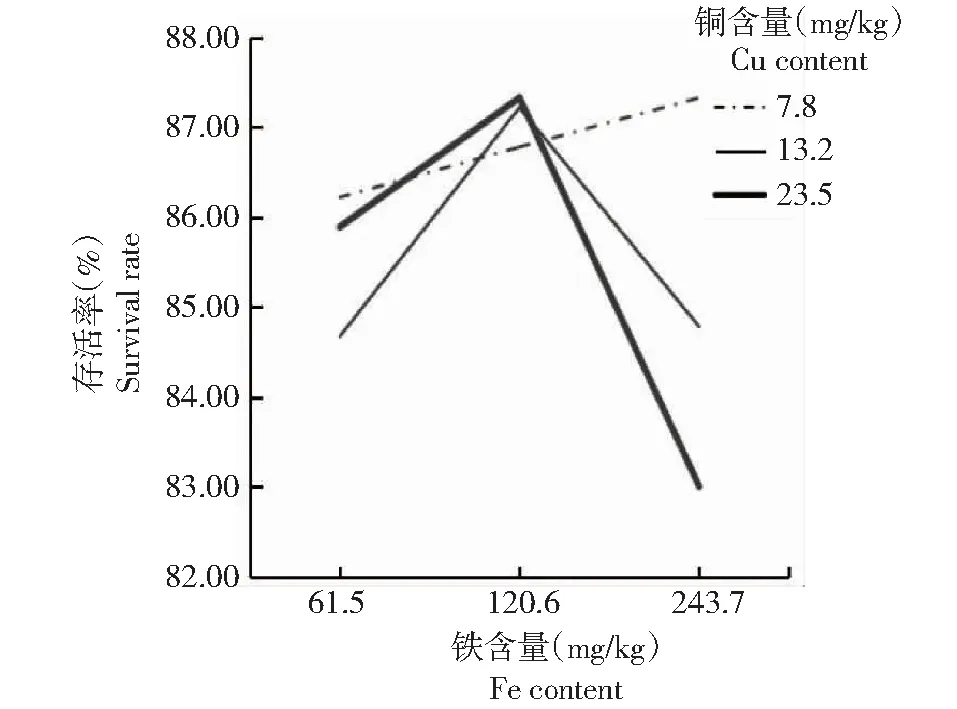
图3 铜与铁互作效应对方格星虫稚虫存活率的影响
2.2 饲料铜和铁水平对方格星虫稚虫体成分的交互影响
由表3可知,各处理方格星虫稚虫的水分含量为81.45%~82.38%,粗蛋白含量为60.40%~66.58%,粗脂肪含量为1.41%~1.70%,灰分含量为21.04%~24.84%。上述指标在各处理间均无显著差异,说明饲料中不同铜和铁含量对方格星虫稚虫体成分指标无明显影响。
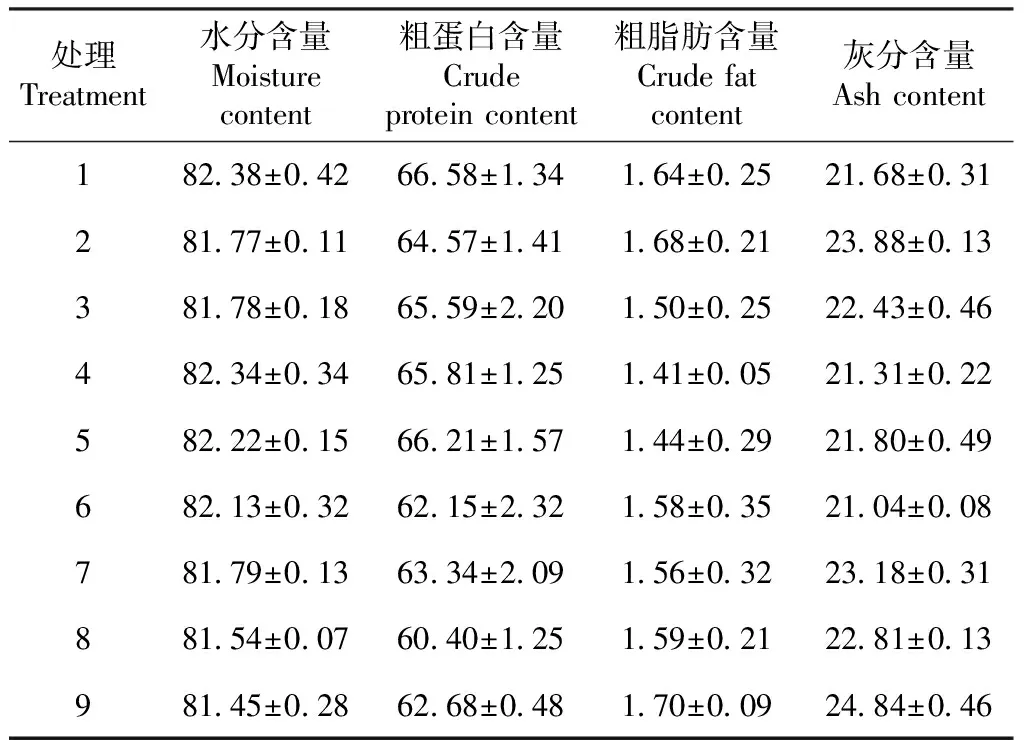
表3 铜铁互作效应下的方格星虫稚虫体成分比较
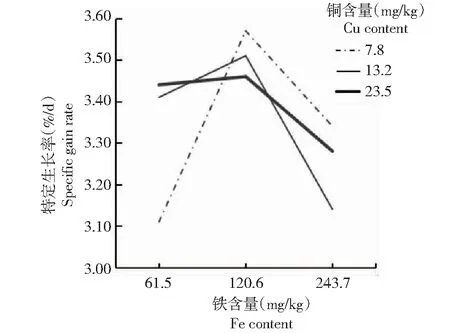
图2 铜与铁互作效应对方格星虫稚虫特定生长率的影响
2.3 饲料中铜和铁水平对方格星虫稚虫CAT活性的交互影响
由表4可知,各处理的方格星虫稚虫CAT活性在2.24~2.81 U/mg,各处理间均无显著差异,表明饲料中不同铜和铁含量对方格星虫稚虫CAT活性无明显影响;当铁含量为61.5 mg/kg(处理1、4和7)时,方格星虫稚虫CAT活性较低;当铁含量增加到120.6 mg/kg以上(处理2、5、8及处理3、6、9)时,方格星虫稚虫CAT活性相对较高但差异不显著,即当铜含量一定时,虫体CAT活性随着铁含量增加呈先增后趋于平稳的变化趋势。可见,方格星虫稚虫CAT活性主要受饲料铁含量影响。从图4可看出,铜含量为7.8 mg/kg的折线与13.2 mg/kg的折线不相交,说明当铜含量在13.2 mg/kg以下时,铜和铁元素含量对虫体CAT活性无明显互作效应;铜含量为23.5 mg/kg的折线与另2条折线均相交,说明当铜元素过量(23.5 mg/kg)时,铜和铁元素含量对虫体CAT活性呈现出一定的互作效应。
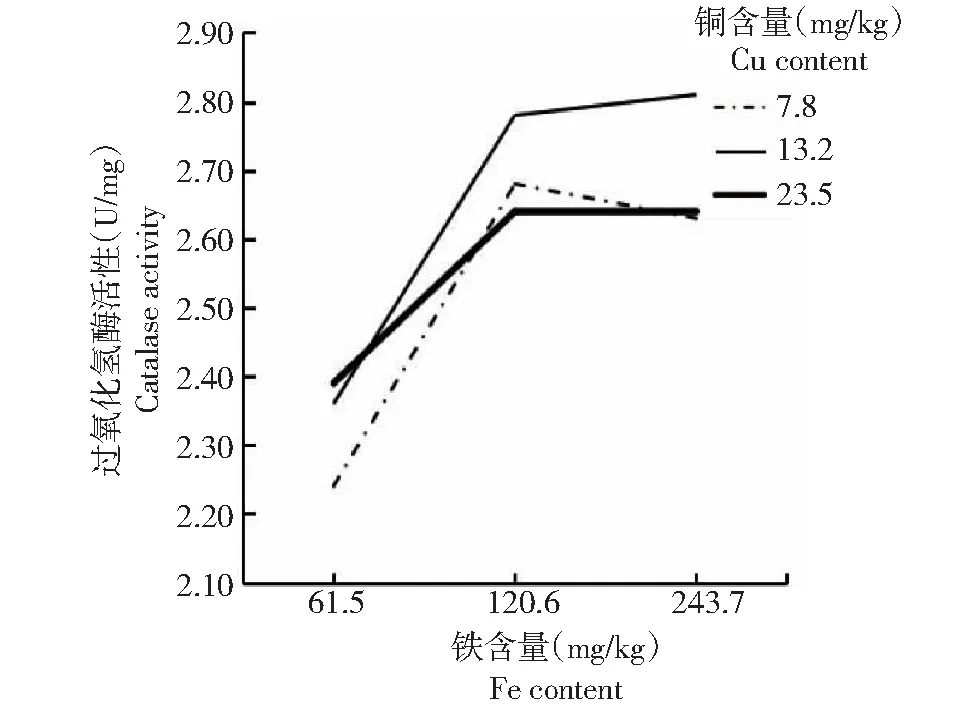
图4 铜与铁互作效应下的方格星虫稚虫CAT活性比较
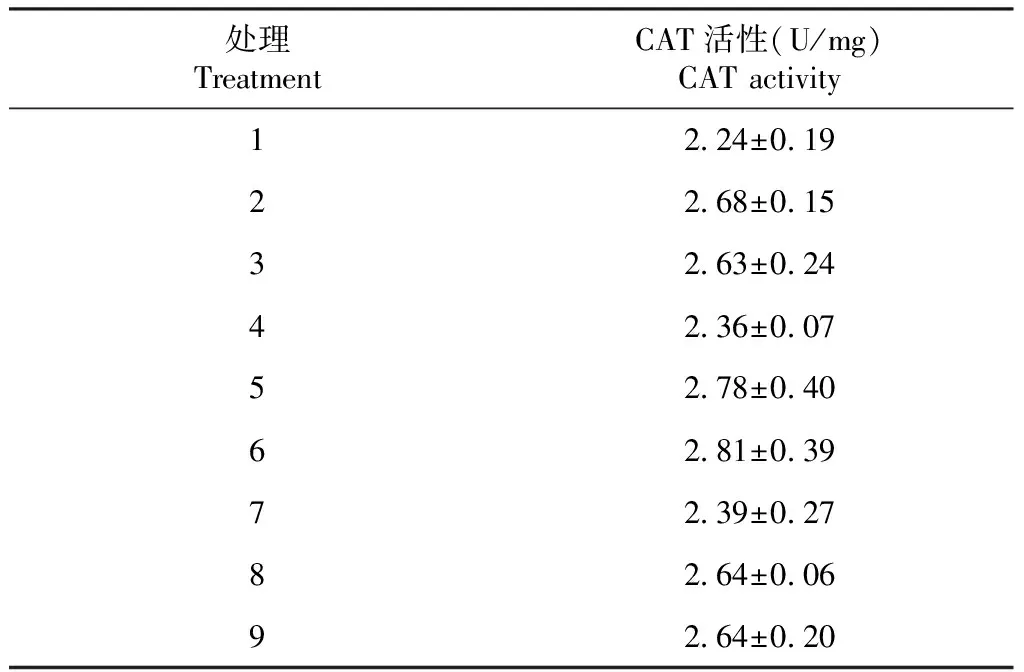
表4 不同铜和铁含量饲料对方格星虫稚虫CAT活性的影响
2.4 饲料中铜和铁水平对方格星虫稚虫虫体铜和铁含量的影响
由表5可知,方格星虫稚虫虫体铜含量随着饲料铜含量的增大而增加。其中,低量铜处理(处理1、2和3)的虫体铜含量为3.14~3.22 μg/g,显著低于适量铜处理(处理4、5和6)和过量铜处理(处理7、8和9);适量铜处理(处理4、5和6)和过量铜含量处理(处理7、8和9)的虫体铜含量为4.75~5.33 μg/g,二者间无显著差异。可见,在铜含量梯度范围内,虫体铜含量与饲料铜含量呈正比关系。从表5还可看出,方格星虫稚虫虫体铁含量在226.71~244.06 μg/g时,各处理间无显著差异,其中,当铜含量偏低(处理1、2和3)和过量(处理7、8和9)时,虫体铁含量随着饲料铁含量的增大而增加,当铜含量为适量(处理4、5和6)时,虫体铁含量随着饲料铁含量增加呈先升后降的变化趋势。

表5 铜与铁互作效应下的方格星虫稚虫虫体铜和铁含量比较
从图5~6折线的相交和平行情况可看出,饲料铜和铁含量对方格星虫稚虫虫体铜含量无明显互作效应,但对虫体铁含量表现出明显的互作效应。
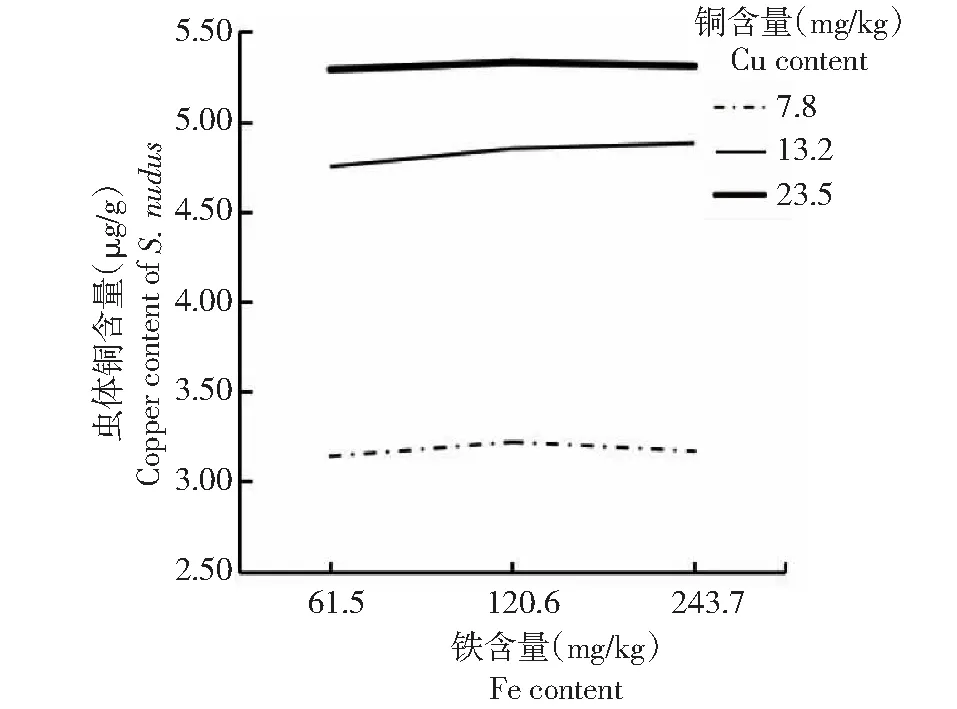
图5 铜与铁互作效应下的方格星虫稚虫虫体铜含量
3 讨 论
矿物质作为多种酶构成成分和活性成分的中心,能调节或激活酶的活性,从消化、吸收和利用等各层次调控能量和蛋白在动物体内的代谢过程,是动物的必需营养素和机体必需成分,具有不可替代的生理生化作用[30-31]。已有研究表明,微量元素的化学形式是影响其利用率的主要因素之一[32-33]。本研究选择利用率较高的氨基酸微量元素螯合物蛋氨酸铜和蛋氨酸铁添加到方格星虫基础饲料中,并根据方格星虫铜和铁单因素试验结果[10-11]选取2种元素的低量、适量和过量3个梯度设计完全随机组合试验,结果表明,不同铜和铁含量饲料对方格星虫稚虫生长和存活均具有显著影响。其中,铜含量为7.8 mg/kg、铁含量为120.6 mg/kg处理方格星虫稚虫的增重率和特定增长率最高;当铁含量为适量(120.6 mg/kg)时,改变铜含量的方格星虫其特定生长率间无显著差异,因此推测方格星虫稚虫的生长率主要受铁元素影响,或铁元素的作用大于铜元素,与卢彩霞[34]对花鱼骨(HemibarbusmaculatusBleeker)饲料的研究结果一致。
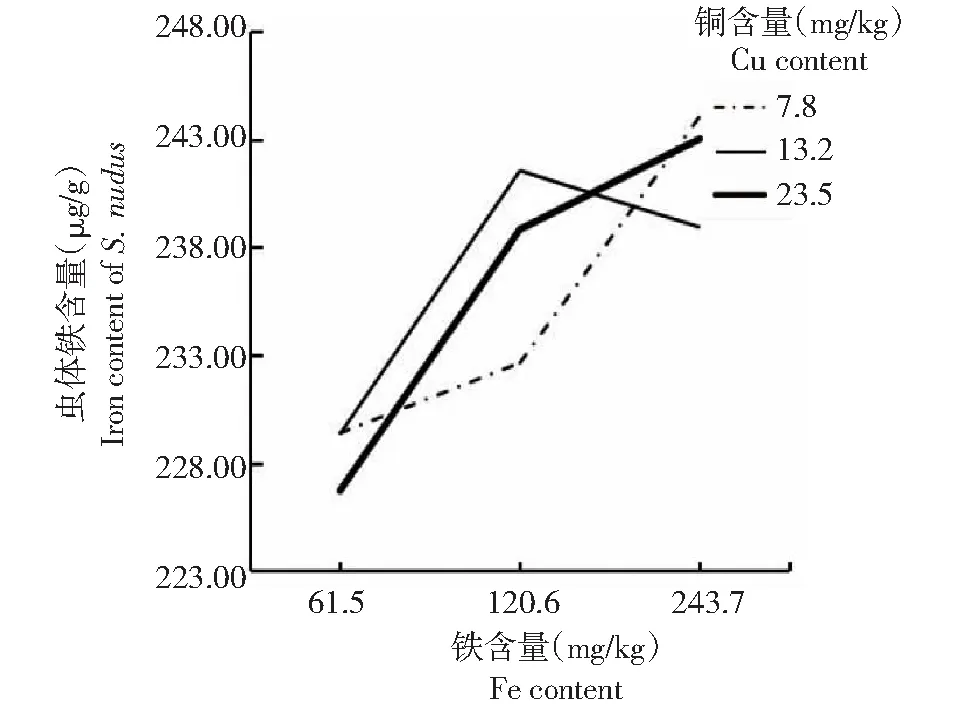
图6 铜与铁互作效应下的方格星虫稚虫虫体铁含量
已有研究表明,铜和铁元素在机体内具有协同作用[35],铜作为造血反应的促媒催化铁合成血红蛋白和生成红细胞,同时参与铁的转运。但也有研究指出,过量的铜可抑制铁的吸收[36]。可见,铜与铁的交互作用复杂多变,且可能与生物体所处生长阶段及使用的微量元素是否过量有一定关系。本研究中,当铁含量适量时,铜含量高低对方格星虫稚虫的生长和成活无显著影响,而当铁含量为低量或过量时,铜与铁产生互作效应,使方格星虫稚虫的生长和成活指标随着铜含量的变化而变化。
本研究结果显示,饲料中不同铜和铁添加量对方格星虫稚虫体成分无显著影响,与已报道的单因素铜或单因素铁对方格星虫稚虫体成分影响的研究结果一致[10-11],也与单因素铜或铁对草鱼(Ctenopharyngodonidellus)[37]、罗非鱼(Oreochromisniloticus)[38]和黄鳝(Monopterusalbus)[39]体成分无显著影响的研究结果一致,究其原因可能是生物体自身物质合成是复杂的过程,微量元素虽然是生物体代谢过程中不可或缺的因素,但多数起着辅助促进作用[29],会影响体内大分子物质的代谢速度,但对体组织成分质量的影响较小。
CAT可清除自由基,是以铁卟啉为辅基的结合酶,1分子酶蛋白中含有4个铁原子,因而铁对CAT活性起着不可或缺的作用[30]。单因素铁营养试验结果表明,当铁水平达115.6 mg/kg时,CAT活性达最大值(3.62 U/mg),4组高铁水平饲料组(61.3、l15.6、283.7和482.1 mg/kg)间的CAT活性无显著差异[11]。本研究中,饲料铁含量梯度为61.5、120.6和243.7 mg/kg,各处理间的CAT活性无显著差异,与许明珠等[11]的研究结果一致,但CAT活性最大值为2.81 U/mg,略低于单因素铁营养试验中的方格星虫虫体CAT活性最大值,可能是受铜铁互作效应影响;当铜含量在13.2 mg/kg以下时,铜和铁元素对方格星虫虫体CAT活性无明显互作效应;当铜含量过量(23.5 mg/kg)时,铜和铁对方格星虫虫体CAT活性呈现一定的互作效应,可能是CAT活性本身主要受铁元素影响,在铜含量低时,铜与铁互作效应不明显,而当铜含量过量时,铜元素对CAT活性具有抑制作用[40],同时与铁元素产生互作效应,使CAT活性出现波动。刘宝娟[41]研究指出,当铜含量一定时,皱纹盘鲍肝胰脏的CAT活性随着铁含量的升高而下降,从而推测饲料中过高的铁含量也会对CAT活性产生抑制作用,本研究结果与其相似。
李俊伟等[13]研究发现,方格星虫虫体内钒、镉和铜等元素受环境沉积物影响较明显,而沉积物中微量元素通过方格星虫的摄食行为进入体内循环并发挥生理生化功能。本研究中,虫体铜含量随着饲料铜含量的增大而增加,与李俊伟等[13]的研究结果相似;虫体铁含量则相对稳定,且各处理间无显著差异,究其原因可能是:方格星虫体腔液中的蚯蚓血红蛋白结合着大量铁离子[42],能维持稳定的铁含量,而虫体铁含量需要维持在一个相对恒定的范围才能维持虫体的正常生命活动。仲晓丽[43]研究发现,饲料中的锌含量对皱纹盘鲍肝胰脏中铁元素的积累无显著影响。本研究中,饲料铜和铁含量对方格星虫稚虫虫体铁含量也无显著影响,与仲晓丽[43]的研究结果相似。本研究还发现,各处理方格星虫虫体的铁含量间差异不显著,但会受到饲料铜与铁含量交互作用的影响而产生波动,与仲晓丽[43]研究发现皱纹盘鲍肌肉中的铁含量随着饲料中锌含量的升高而显著增加的结果不一致,推测是其肌肉组织内锌和铁元素间产生交互效应的结果。
4 结 论
饲料中铜和铁的交互作用可显著影响方格星虫稚虫生长、存活及虫体铜含量,对方格星虫稚虫体成分、CAT活性和虫体铁含量无显著影响。饲料铁元素对方格星虫稚虫生长的影响程度大于饲料铜元素。以增重率为评价标准,方格星虫稚虫饲料中最适铜和铁含量分别为7.8和120.6 mg/kg。
猜你喜欢
云南化工(2021年10期)2021-12-21
防护林科技(2020年9期)2020-11-09
中南林业调查规划(2018年3期)2018-11-24
家庭医学(2017年9期)2017-10-24
中南林业调查规划(2015年3期)2015-12-20
生态学报(2014年8期)2014-05-05
亲子(2014年3期)2014-04-18
郑州大学学报(理学版)(2014年4期)2014-03-01
故事作文·高年级(2009年11期)2009-12-11
故事作文·高年级(2009年9期)2009-12-11
