
油菜不同还田量对新疆盐碱土壤线虫群落的影响
2023-03-29 06:14张凤华
干旱地区农业研究 2023年1期
杨 涛,陈 虹,张凤华
(石河子大学,新疆生产建设兵团绿洲生态农业重点实验室,新疆 石河子 832003)
土壤线虫是土壤生态系统的重要组成部分,能够通过调控物质的分解和转化影响土壤环境[1-2]。除此之外,线虫还能通过影响生长素和赤霉素等其他物质的含量以调节土壤环境[3]。因此土壤线虫常被看作是评价土壤环境和植物条件的指示生物[4]。在农田管理措施中,耕作、灌溉以及施肥等都能改变土壤肥力及土壤微生物特性,从而影响线虫群落[5]。已有研究表明,油菜等秸秆还田可显著提高紫色土中线虫、蚯蚓等土壤动物数量[6]。
据调查,新疆盐碱地面积占可耕地总面积的32.07%[7],因此,盐碱改良是我国目前要解决的重大难题。近年来,绿肥还田作为一项新兴的生物改良技术,能够提升土壤肥力、改善土壤环境、提高后茬作物产量[8]。有研究表明,油菜和紫云英还田能够提高土壤有机质和有效氮、磷的含量以及水稻产量。油菜作为十字花科芸薹属作物,其抗逆性强、适应性广[9]。油菜作绿肥可以提高土壤肥力,还能降低土壤盐分含量[10];此外,油菜所含的硫甙类物质能够有效抑制杂草及病虫害,根系的分泌物可增强土壤微生物活性[11]。
目前对油菜作绿肥的研究主要集中在养分释放及种植技术等方面[12],而针对油菜还田后线虫群落变化特征的研究相对较少。因此本试验采用高通量测序技术对新疆盐碱土壤油菜作绿肥还田后的土壤线虫群落进行研究,阐明油菜还田对土壤线虫群落结构的影响,揭示影响土壤线虫群落结构变化的主要环境因子,为新疆盐碱土壤生物改良提供科学的理论依据。
1 材料与方法
1.1 研究区概况
研究区位于新疆生产建设兵团第八师石总场(44°27′47″N,86°0′54″E),光照充足,雨量稀少,属于典型的温带大陆性气候。该区年平均气温6.6℃,年平均降水量180 mm,年平均蒸发量1 600 mm,年平均日照时数2 784 h,年平均无霜期166 d,≥0℃积温3 918℃。该区域地下水位较高,强烈的蒸发使得土壤盐分表聚,耕层土壤盐渍化程度较重。2020年10月测定研究区土壤理化性质如表1所示。

表1 研究区土壤理化性质(2020年)Table 1 Physical and chemical properties of soil in the study area (2020)
1.2 试验设计
试验于2020年7月15日进行油菜播种。油菜品种为‘华油杂62号’(甘蓝型油菜,由华中农业大学提供),播种量为15 kg·hm-2,滴灌栽培。播种时施用重过磷酸钙300 kg·hm-2,出苗后30 d开始随水追施尿素,全生育期浇水6~7次,其他田间管理与实际生产一致。于2020年10月17日进行油菜打杆,10月27日进行油菜粉碎翻压还田,油菜地上部鲜重为84 000 kg·hm-2。试验设置油菜不还田(CK)、还田量21 000 kg·hm-2(T1)、还田量42 000 kg·hm-2(T2)、还田量84 000 kg·hm-2(T3)4个处理。每个处理设置3次重复,共计12个小区,小区面积为5 m×5 m=25 m2,采取单因素完全随机区组设计,每两小区之间均设置2 m保护行。
1.3 样品采集
于2021年7月7日进行土壤样品采集,取样深度为0~20 cm耕层土壤。采用五点混合取样法进行取样,各处理的3个重复小区分别随机选取5个样点,将5个样点土样去除动植物残体及石块等杂物后均匀混合(3次重复进行平均),置于透明无菌自封袋内,放入冰盒冷藏,并及时带回实验室。土壤样品过1 mm土筛后分为2份,一份储存在-80℃冰箱中用于线虫群落的分析,另一份自然风干后用于土壤理化性质的测定。
1.4 测定指标与方法
1.4.1 土壤理化性质测定 土壤容重采用环刀法进行测定;土壤含水率采用烘干法进行测定;土壤pH采用哈希HQ411d台式酸度计进行测定(蒸馏水浸提,水土比为5∶1);土壤电导率采用雷磁DDSJ-308F型电导率仪进行测定(蒸馏水浸提,水土比为5∶1);土壤总盐含量采用残渣烘干-质量法测定;有机质采用重铬酸钾容量法-外加热法测定:碱解氮采用碱解扩散法测定;速效磷采用NaHCO3浸提-钼锑抗比色法测定;速效钾采用NH4OAc浸提-火焰光度计法测定[13]。
1.4.2 土壤DNA提取、PCR扩增及高通量测序 采用土壤DNA提取试剂盒(MOBIO UltraCleanTM)从0.5 g土壤样品中提取总DNA,保存于-20℃冰箱中备用。利用凝胶电泳检测基因组DNA,使用酶标仪测定其浓度和纯度。对样品进行序列检测扩增,扩增区域为NF1F_18Sr2bR,扩增片段长度为350 bp。PCR产物利用QuantiFluorTM-ST蓝色荧光定量系统(Promega 公司,美国)进行定量,混合后进行Miseq文库构建、拼接质控、OTU聚类及生信分析。采用Miseq PE300平台进行序列的测序及分析(上海美吉生物医药科技有限公司)。
1.4.3 土壤线虫群落分析 根据土壤线虫的特征结构及捕食习性将其分为4个营养类群:食细菌类线虫(Bacterivores)、食真菌类线虫(Fungivores)、植物寄生类线虫(Plant-parasites)和捕食杂食类线虫(Omnivores-predators);并根据线虫的食性和生活策略划分不同的生活史c-p值。每100 g土壤中食细菌类线虫、食真菌类线虫、植物寄生类线虫和捕食杂食类线虫占线虫总数比例为各营养类群的相对丰度;根据土壤线虫的分类结果计算其生态指数,即:
(1)线虫通道指数(nematode channel ratio, NCR):NCR=B/(B+F)。式中,B为食细菌类线虫所占比例;F为食真菌类线虫所占比例。
(2)自由生活线虫成熟度指数(maturity index of free-living nematodes, MI);植物寄生类线虫成熟度指数(plant parasite index, PPI):MI(PPI)=∑vi×fi。其中,vi为自由生活线虫(或植物寄生类线虫)被赋予的c-p 值,fi为第i种线虫的数量占群落总数的比例。
(3)富集指数(EI):EI=100×e/(b+e);结构指数(SI):SI=100×s/(b+s);式中,b代表食物网基础成分,指Ba2和Fu2这两个类群;e代表食物网富集成分,指Ba1和Fu2这两个类群;s代表食物网结构成分,包括Ba3-Ba5、Fu3-Fu5、Om2-Om5类群(其中Ba2代表食细菌类线虫中c-p值为2的线虫数量,Fu2代表食真菌类线虫中c-p值为2的线虫数量,Om2代表捕食杂食类线虫中c-p值为2的线虫数量)。
1.5 数据处理与分析
利用Microsoft Excel 2021对土壤理化结果进行整理,采用SPSS 26进行方差分析和相关性分析,采用Duncan法检验处理间差异显著性(P<0.05)。
高通量测序结果使用Fastp 0.19.6 进行质控,采用Flash 1.2.11进行双端序列拼接获得有效数据。使用QIIME软件包挑选各OTU的代表序列,并将其与数据库进行对比注释。采用Uparse 7.0.1090软件按照97%相似性进行OTU聚类,各OTU中丰度最高的序列为代表序列,所有样本的覆盖率均高于99.7%。采用RDP Classifier 2.11贝斯叶算法对97%相似性的OTU代表序列进行序列分类注释,设置信度阈值为0.7,与NT(Nucleotide Sequence Database)数据库进行数据比对。采用Origin 2021对群落组成作图。采用Mothur 1.30.2对优化序列提取非重复序列,在97%相似水平分析样品多样性指数。采用主坐标分析(PCoA)分析油菜不同还田量处理土壤线虫群落组成差异。利用方差膨胀因子(VIF)筛选环境因子,对土壤理化性质及土壤线虫群落结构进行冗余分析(RDA)。
2 结果与分析
2.1 油菜不同还田量土壤理化性质
从表2可以看出,与CK相比,油菜各还田处理均显著降低了土壤容重、pH、EC及总盐含量(P<0.05)。土壤含水率随着还田量的增加逐渐上升,T3处理最高,较CK增加了23.66%。与CK相比,油菜各还田处理均显著增加了土壤有机质含量,T2处理最高,较CK增加了27.42%。T3处理土壤碱解氮与速效钾含量均显著高于其他处理(P<0.05),与CK相比分别增加了35.46%和11.12%;而土壤速效磷含量在T2处理时最高,与CK相比增加了56.48%,且与其他处理之间差异显著(P<0.05)。
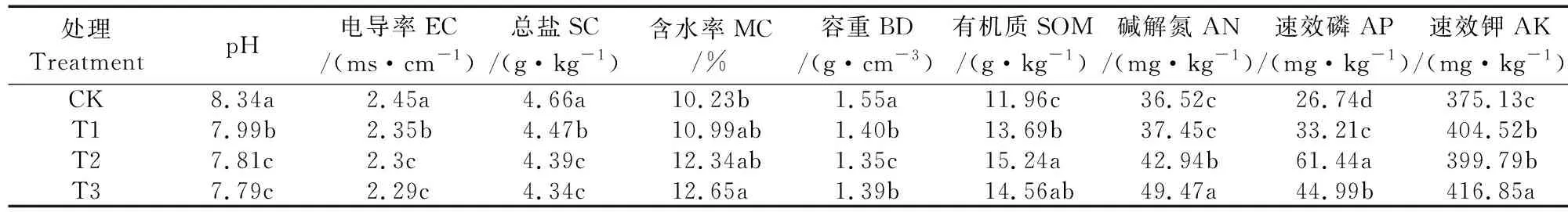
表2 油菜不同还田量土壤理化性质Table 2 Soil physiochemical properties in different rapeseed returning amounts
2.2 油菜不同还田量土壤线虫群落
对各样本土壤线虫进行高通量测序,保留了217 864条有效序列,有效序列长度为220~434 bp,平均序列长度为320 bp。基于97%的序列相似性进行聚类,所有样品中共检测出40个OTU,被划分为3纲3目13科19属24种。其中食细菌类线虫5属,食真菌类线虫4属,植物寄生类线虫3属,捕食杂食类线虫4属,未分类线虫3属。
从图1a可以看出,各处理中相对丰度≥1%的线虫科共有4个,分别为:头叶科(Cephalobidae)、孔咽科(Aporcelaimidae)、盆咽科(Panagrolaimidae)和异皮科(Heteroderidae)。各处理土壤线虫的主要线虫科均是头叶科,在T1处理相对丰度最高,达89.98%;在T3处理最低,为81.97%。随着还田量的增加,孔咽科的相对丰度呈下降趋势,异皮科则相反。粒科(Anguinidae)与刺科(Belonolaimidae)仅在油菜还田处理中被检测出。
属水平土壤线虫群落组成如图1b,平均相对丰度≥1%的线虫属共有5个,分别为:拟丽突属(Acrobeloides)、丽突属(Acrobeles)、无环属(Aporcella)、盆咽属(Panagrolaimus)和刻点胞囊属(Punctodera)。其中,拟丽突属在所有处理中均为优势属(平均丰度>75.00%),在T1处理中相对丰度最高,达87.75%。无环属的相对丰度随还田量的增加逐渐下降,T3处理最低,较CK降低了99.82%。而刻点胞囊属的相对丰度则随着还田量的增加而升高。米卡垫刃属(Miculenchus)与刺属(Belonolaimus)仅在还田处理中被检测出。
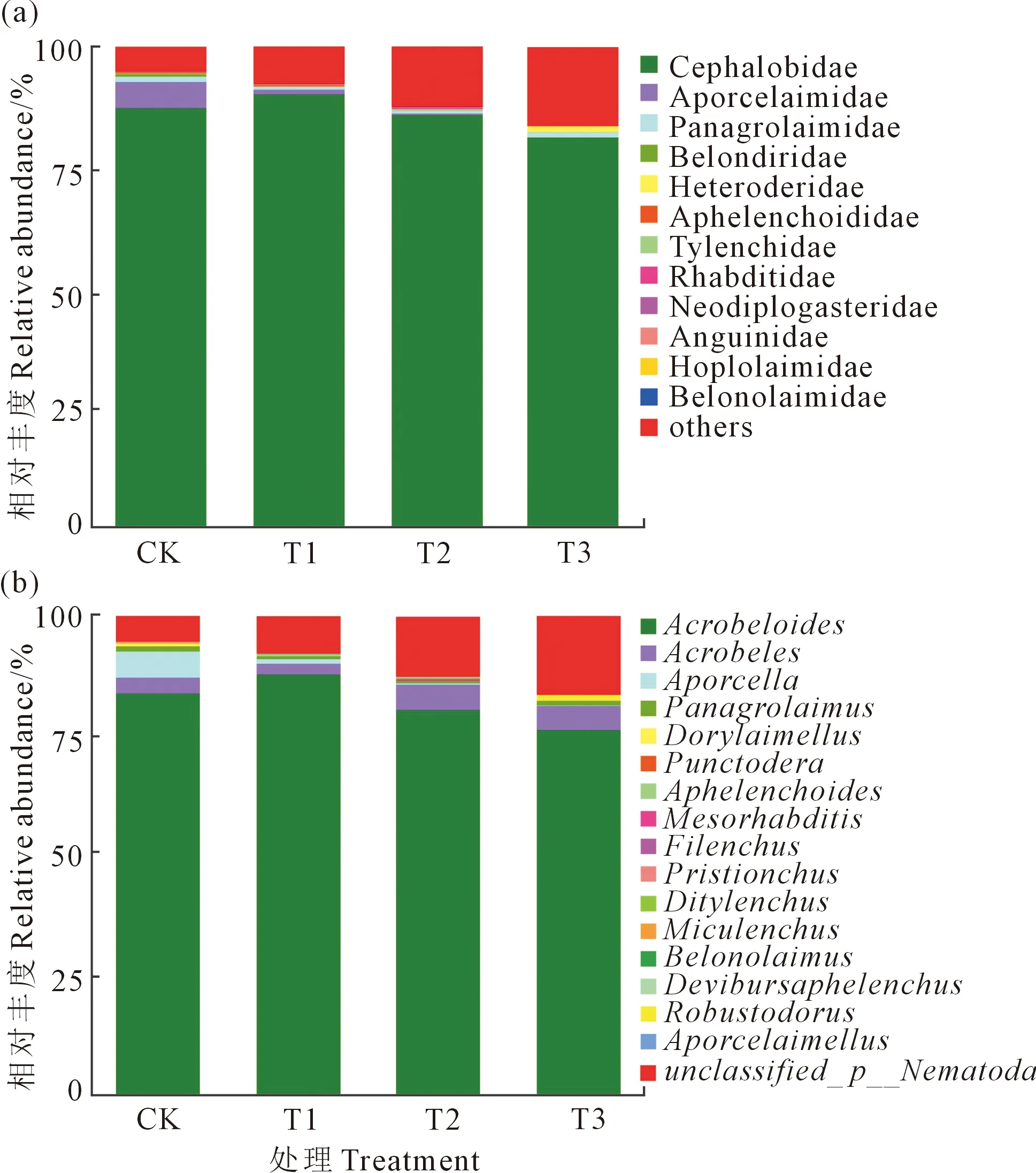
图1 不同还田量下线虫科(a)和属(b)的相对丰度Fig.1 Relative abundance of nematode family (a) and genus (b) in different returning amounts
不同还田量处理土壤线虫各营养类群的相对丰度如图2所示。所有处理优势营养类群均为食细菌类线虫,T2处理下其相对丰度最高,随后呈下降趋势;捕食杂食类线虫的相对丰度随还田量的增加呈下降趋势,而食真菌类线虫与植物寄生类线虫的相对丰度逐渐升高。

图2 不同还田量下土壤线虫各营养类群相对丰度Fig.2 Relative abundance of soil nematode in different returning amounts
通过分析不同还田量处理线虫群落的丰富度指数、多样性指数、线虫通道指数、成熟度指数等发现,所有处理覆盖度均大于0.9970(表3),这说明测序数据合理,能够较真实地反映油菜还田后土壤样本的线虫群落特征。如图3所示,Chao1指数与ACE指数均随着还田量的增加而降低,CK处理最高,T3处理最低;T3处理Chao1指数和ACE指数较CK分别降低41.07%和35.44%。Shannon指数在T2处理最高,T1处理最低,而Simpson指数则相反。从表3可以看出,自由生活线虫成熟度指数MI随着还田量的增加呈下降趋势,CK处理最高,T3处理最低,T3较CK降低了16.80%;与CK相比,油菜还田处理植物寄生线虫成熟度指数PPI均有所增加,处理间差异不显著(P>0.05);油菜还田处理富集指数EI均低于CK但处理间差异不显著,而结构指数SI较CK均显著降低(P<0.05)。
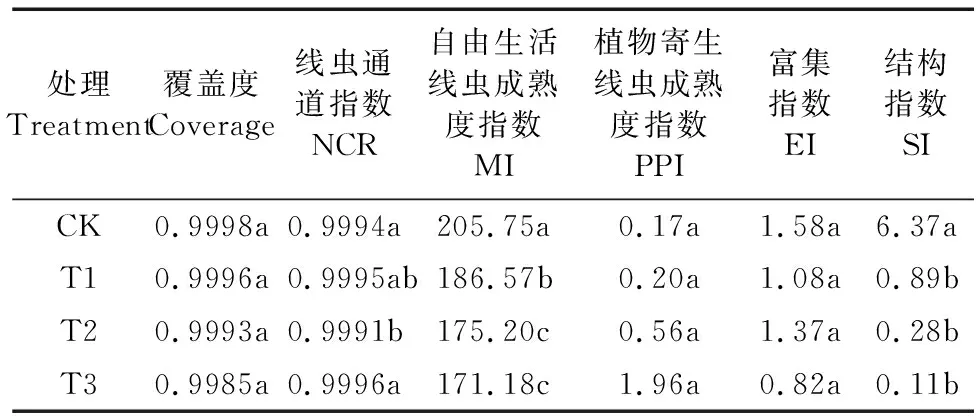
表3 油菜不同还田量下土壤线虫生态指数Table 3 Ecological index of soil nematodes in different returning amounts
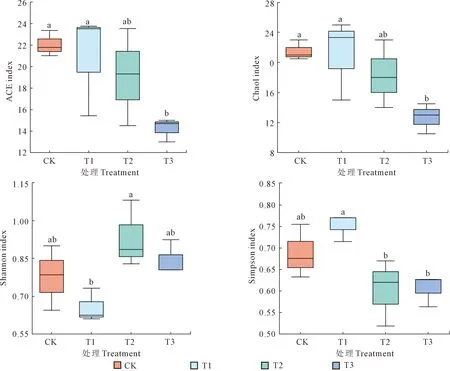
图3 不同还田量下土壤线虫多样性指数Fig.3 Diversity index of soil nematodes in different returning amounts
如图4所示,在OTU水平上进行PCoA分析,2个主成分坐标轴共解释了总变异的91.73%,PCoA1和PCoA2分别解释了总变异的77.24%和14.49%。说明不同处理之间土壤线虫群落存在差异。
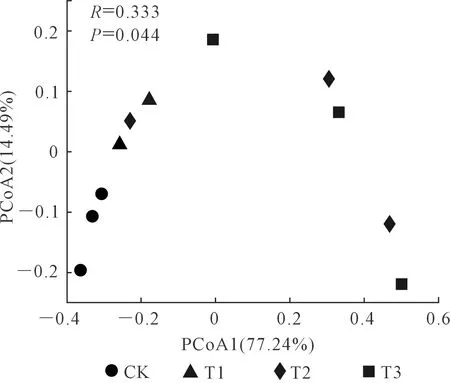
图4 不同还田量土壤线虫群落组成主坐标(PCoA)分析Fig.4 Principal co-ordinates analysis (PCoA) of soil nematode community composition in the treatments with different returning amounts
2.3 土壤理化性质对线虫群落的影响
利用方差膨胀因子VIF分析筛选,保留了土壤有机质、碱解氮、速效磷、速效钾和含水率等环境因子。如图5为土壤理化性质与土壤线虫群落结构间的冗余分析(RDA)。RDA1和RDA2共同解释了线虫群落总变异的97.94%,分别解释了总变异的93.74%和0.24%。土壤有机质(R2=0.666,P=0.005)和速效钾(R2=0.845,P=0.001)是影响土壤线虫群落结构的主要土壤环境因子(图5,表4)。
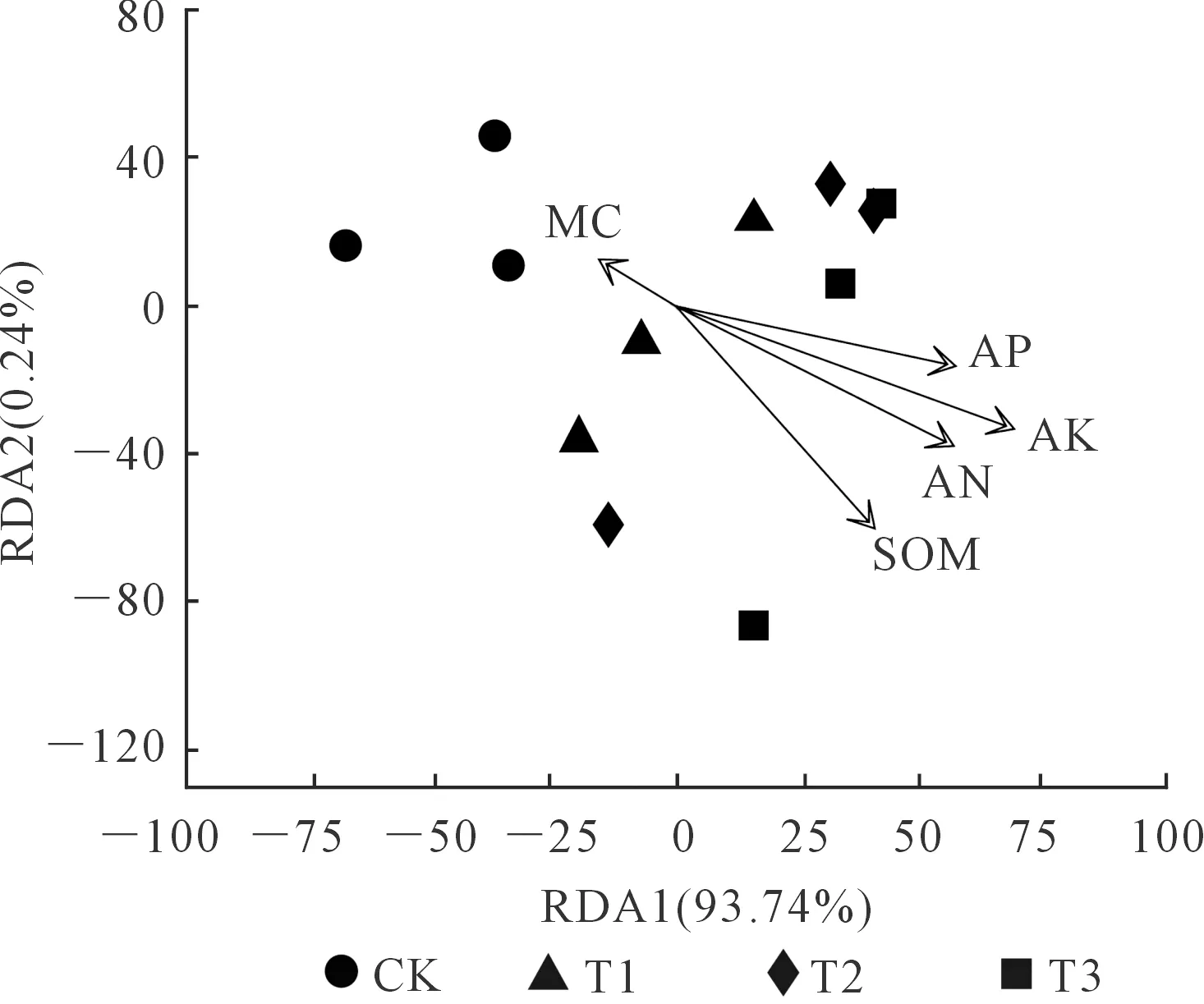
注:SOM:有机质;AN:碱解氮;AP:速效磷;AK:速效钾;MC:含水率。注:SOM:Soil organic matter; AN:Alkali hydrolyzed nitrogen; AP:Available phosphorus; AK:Available potassium; MC:Moisture content.图5 土壤理化性质与线虫群落间的冗余分析(RDA)Fig.5 Redundancy analysis (RDA) of soil physical and chemical properties and soil nematode communities
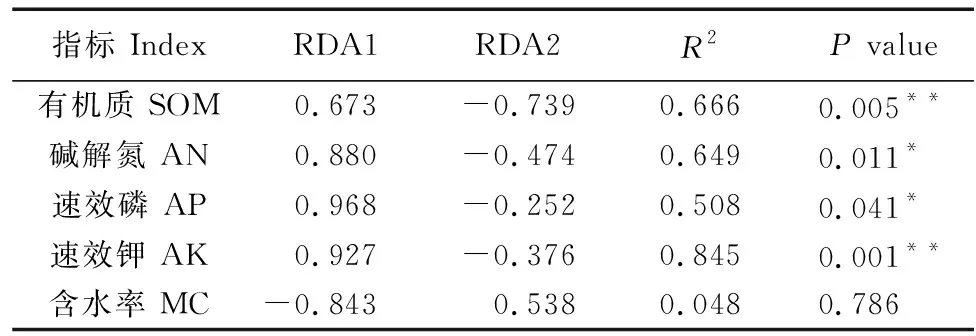
表4 Envfit环境因子表Table 4 Envfit environmental factor table
表5为土壤理化性质与土壤线虫群落生态指数的相关性。Chao1指数和Simpson指数与土壤EC呈显著正相关,与土壤碱解氮显著负相关;自由生活线虫成熟度指数MI和结构指数SI与土壤pH、EC及总盐含量呈极显著正相关,与土壤碱解氮呈极显著负相关。
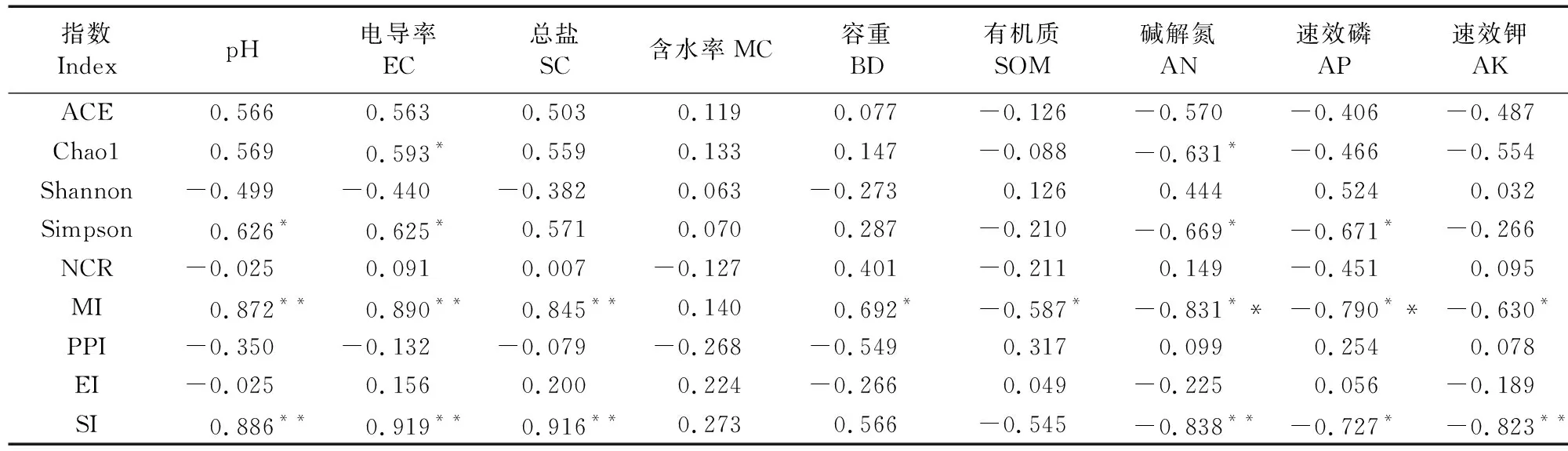
表5 土壤理化性质与土壤线虫生态指数的相关系数Table 5 Correlation coefficients between soil physiochemical properties and soil nematode ecological index
3 讨论与结论
3.1 油菜不同还田量对土壤理化性质的影响
本研究结果表明,油菜还田显著降低了土壤容重、pH、EC和总盐含量,且随着油菜还田量的增加,土壤含水率呈上升趋势。已有研究表明油菜能够通过自身代谢合成大量有机酸,从而调节自身的渗透平衡。当油菜翻压还田后,其体内的有机酸进入土壤中,加之微生物对油菜植株的分解也会产生部分有机酸,从而降低了土壤pH[14]。还有研究表明绿肥还田能够增加土壤孔隙度,从而疏松土壤,降低土壤容重[15]。绿肥还田后还能与土壤颗粒胶结形成稳定的团粒结构,从而增强土壤透气性和透水性[16]。土壤盐分含量下降可能是由于油菜对土壤Na+的吸收能力显著高于其他作物,种植并翻压油菜可以显著降低土壤含盐量和Na+含量[10]。
本研究还表明油菜还田能提高土壤养分含量,这与邓力超等[17]的研究结果一致。油菜还田后能产生大量可溶性有机物,并将自身的多种元素归还到土壤当中,提供多种有效养分,促进土壤养分的循环[18]。与CK相比,不同还田量处理速效磷含量均显著提高,这是因为油菜还田能活化土壤中的难溶性磷,使得土壤有效磷含量增加[19]。除此之外,油菜还田还能通过提高土壤酸性磷酸酶的活性来提高土壤有效磷含量[20]。
3.2 油菜不同还田量对土壤线虫群落的影响
相对于形态学鉴定,高通量测序技术的发展及应用为土壤生物群落研究提供了新途径[21]。本研究通过高通量测序对土壤线虫进行了鉴定,研究结果表明,食细菌类线虫与捕食杂食类线虫的相对丰度随着还田量的增加呈下降趋势,而食真菌类线虫与植物寄生类线虫的相对丰度呈上升趋势。有研究表明秸秆还田过程中能产生大量真菌,从而使其相对丰度增加[22]。捕食杂食类线虫对土壤扰动响应极为敏感,油菜还田过程中对土壤的扰动可能会使其相对丰度降低甚至导致一些属消失。叶成龙等[5]研究发现施用有机肥能增加土壤植物寄生类线虫的相对丰度,这可能是还田增加了植物生物量,形成了更多的取食位点,有利于植物寄生类线虫的取食与繁衍,进而增加其相对丰度[23]。
线虫生态指数能全面地反映线虫群落之间的差异。本研究结果表明油菜还田降低了土壤线虫群落的丰富度,提高了线虫群落的多样性,且部分属如米卡垫刃属与刺属仅在还田处理中被检测出,说明油菜还田提高了食物网的复杂性,使生态系统趋于成熟稳定。本研究结果表明所有处理线虫通道指数NCR均大于0.5,说明有机物质分解主要以细菌分解为主。自由生活线虫成熟度指数MI随着还田量的增加而下降,而植物寄生线虫成熟度指数PPI呈上升趋势。这可能是由于油菜还田增加了植物对线虫取食的承载能力,使得植物寄生类线虫成熟度指数PPI增加[24]。富集指数EI与结构指数SI可以指示土壤环境所受到的扰动和食物网的状况。本研究发现富集指数EI与结构指数SI较CK均有所下降,T3处理最低,这可能是由于油菜还田借助机械耕作,对土壤扰动程度较高,使得土壤处于胁迫状态,原本食物网遭到破坏,其抗外界干扰能力变弱。
3.3 土壤理化性质对线虫群落的影响
土壤理化性质的变化会影响土壤线虫群落结构,不同的理化性质对线虫群落有着不同影响。本研究通过RDA冗余分析,表明土壤有机质和土壤速效钾是影响土壤线虫群落变化的主要环境因子。Chao1指数与土壤EC呈显著正相关,与土壤碱解氮呈显著负相关,这与陈丽燕等[25]的研究结果一致。已有研究表明土壤线虫与土壤有机质的分解和土壤物质能量循环密切相关[6],因为油菜还田增加了土壤有机质含量,使得线虫食物来源更加丰富,促进了土壤食物网结构化,从而增加其多样性。卢焱焱等[26]对红壤线虫群落的研究表明,微生物量碳和有效氮更能影响土壤线虫群落组成。这些环境因子的差异可能与试验地土壤及气候条件、试验处理及方法等因素不同有关。
综上所述,油菜还田能降低土壤容重及盐分含量,改善土壤结构,培肥效果显著;油菜还田还降低了食细菌类线虫的相对丰度,提高了食真菌类线虫与植物寄生类线虫的相对丰度,增加了土壤线虫群落的多样性和复杂性,降低了土壤线虫群落的结构指数。影响土壤线虫群落变化的主要环境因子为土壤有机质和速效钾。
猜你喜欢
特种经济动植物(2023年2期)2023-02-17
今日农业(2022年14期)2022-09-15
浙江农业科学(2019年11期)2019-11-09
节水灌溉(2018年11期)2018-11-28
中成药(2018年2期)2018-05-09
现代园艺(2017年21期)2018-01-03
浙江大学学报(农业与生命科学版)(2017年1期)2017-04-17
绿色科技(2016年16期)2016-10-11
天然产物研究与开发(2016年6期)2016-06-05
亚热带资源与环境学报(2015年1期)2015-01-22
