
高寒草地植物多样性与Godron 群落稳定性关系及其环境驱动因素
2023-03-21 09:18:06雷石龙廖李容王杰张路叶振城刘国彬张超
草业学报 2023年3期
雷石龙,廖李容,王杰,张路,叶振城,刘国彬,张超
(1. 西北农林科技大学水土保持研究所,陕西 杨凌 712100;2. 中国科学院水利部水土保持研究所,陕西 杨凌 712100;3. 中国科学院大学,北京 100049)
作为生物多样性重要组成部分,植物多样性在维持群落稳定性和生态系统功能方面起着至关重要的作用[1]。近几十年来,植物多样性与群落稳定性之间关系及驱动机制一直是生态学的研究热点[2]。研究表明,植物多样性高的群落,其群落稳定性也较高[3],也有研究发现群落稳定性与植物多样性在一定环境下呈负相关关系[4],一些学者也在不同生境中证明了两者之间并非简单的相关关系[5-6]。目前植物多样性与群落稳定性之间的关系仍存在争议。因此有必要开展不同环境梯度下植物多样性与群落稳定性相关关系的研究,揭示植物群落稳定性的维持机制。
草地是陆地上分布最广的生态系统,在生物多样性保护、全球变化应对、碳平衡维持,以及食物的供应中发挥着重要作用[7]。作为“地球第三极”,青藏高原拥有全球最大的高寒草地,面积约16.5×105km2,占中国草地面积的42%[8]。高寒草地作为青藏高原主要的植被类型,其生长季短[9]、自我调节能力弱[10]、群落结构简单但类型复杂[11],是我国最脆弱的生态系统类型之一[12]。由于其在气候调节、水源涵养和多样性维持中发挥着重要作用[13],高寒草地近年来受到众多学者的关注。其中,多样性沿环境梯度的空间变化及与群落稳定性之间的关系是植物群落研究的关键问题[14-15]。例如,吴红宝等[16]发现随着海拔的升高,高寒草地Shannon-Wiener 指数和物种丰富度先增加后降低,而Pielou 均匀度指数先降低后增加;武建双等[17]研究发现高寒草地植物多样性随着生长季累积降水呈现指数增加趋势;薛晨阳等[18]研究发现群落稳定性与多样性无显著关系。这些研究阐明了植物多样性和群落稳定性随海拔、降水量等环境梯度的变化趋势,对理解植物多样性和群落稳定性有重要意义。但目前关于生物多样性与群落稳定性的关系尚存在争议,仍需深入研究[19]。本研究以青藏高原的高寒草甸、高寒湿地草甸、高寒草原和高寒荒漠草原为研究对象,探究不同草地类型植物多样性与稳定性之间关系及其对土壤因子的响应,揭示高寒草地植物多样性和群落稳定性关系,为青藏高原高寒草地生态系统可持续发展提供理论依据。
1 材料与方法
1.1 研究区域概况
研究区位于念青唐古拉山和冈底斯山之间,是青藏高原的核心地区,东西横跨1400 km(79.71°-94.54° E),南北纵跨570 km(28.74°-33.72° N),总面积约45.57 万km2。该区域气候属于典型的高寒大陆性季风气候,寒冷干燥,年平均降水为88.9~526.1 mm,年平均气温为4.1~7.6 ℃,海拔3810~5020 m[20]。研究区域内包含高寒湿地草甸、高寒草甸、高寒草原和高寒荒漠草原4 种草地类型,主要物种为藏北蒿草(Kobresia littledalei),高山蒿草(Kobresia pygmaea),紫花针茅(Stipa purpurea)和沙生针茅(Stipa glareosa)。
1.2 植被调查与样品采集
于2020年7 月在研究区域内选取高寒湿地草甸、高寒草甸、高寒草原和高寒荒漠草原4 种草地类型各20 个,共计80 个样点,每两个样点之间间隔20 km。在每个样点选取远离道路、受人为干扰较少的、地势相同的草地,设置100 m×100 m 的样地,每个样地沿对角线设置6 个1 m×1 m 的观测样方,相邻两个样方之间间隔20 m,共480个样方。在每个样方内用直径为7 cm 的土钻采用5 点法取0~20 cm 土层,充分混合,共采集480 个土样。研究区域及样点分布见图1。
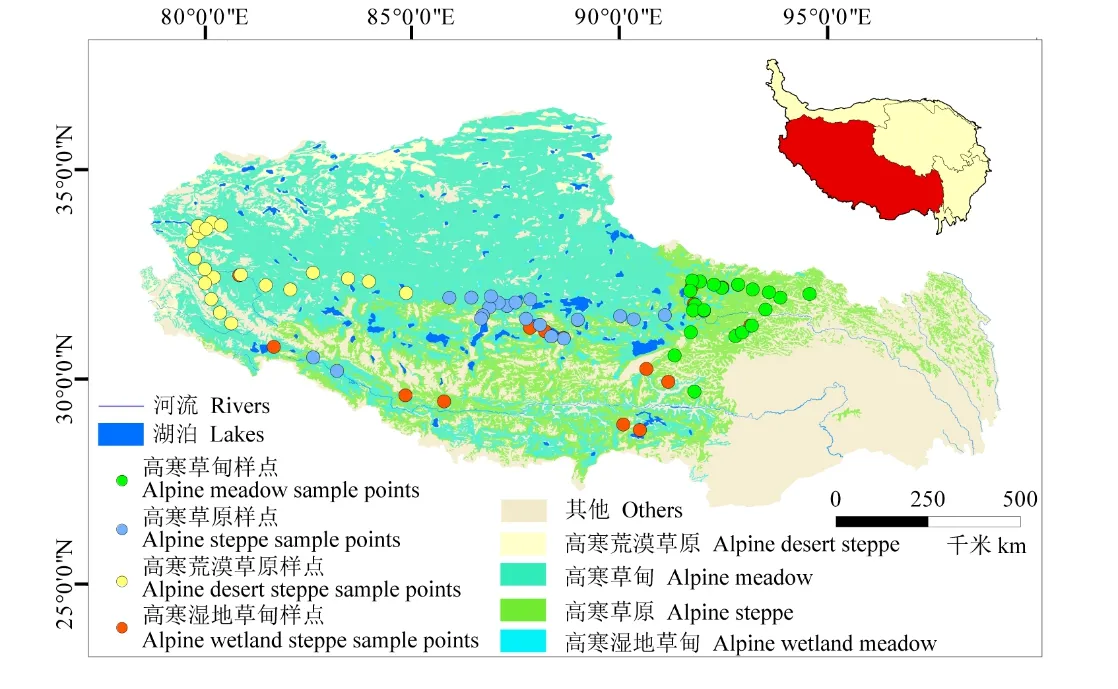
图1 研究区域及样点分布Fig.1 Study area and sample point distribution
采用刈割法将植物按照物种齐根割掉,每割掉一个物种均对刈割前后的样方进行数码相机拍照,将照片图像用CAN-EYE-V6313 软件进行分析,获取样方总盖度和每种植物的分盖度[21-22]。植株高度使用钢卷尺测量。割掉的植物按照物种进行分类标号并装在信封中,放入65 ℃烘箱中烘至恒重,样方内所有物种的生物量相加即为总生物量。以科为单位将每个样方内植物分为莎草、禾草、豆科和杂类草4 个功能群,样方内各科植物的生物量相加即为各功能群的总生物量。pH 采取玻璃电极法测定;土壤有机碳(soil organic carbon,SOC)用重铬酸盐法测定;全磷(total phosphorus,TP)含量采用NaOH 熔融-钼锑抗比色法测定;硝态氮(nitrate nitrogen,NO3--N)用紫外分光光度法测定;速效磷(available phosphorus,AP)和铵态氮(ammonium nitrogen,NH4+-N)用分光光度计法测定[23-25]。
1.3 指标计算
选 取 物 种 丰 富 度 指 数(species richness index,S)、Shannon-Wiener 多 样 性 指 数(Shannon-Wiener biodiversity index,H)和Pielou 均匀度指数(Pielou evenness index,J)来表征植物多样性,计算公式如下:
式中:C为样方内的物种数,Pi为相对重要值。
Godron 稳定性(G)计算方法是目前生态学应用较为广泛的测定群落稳定性的方法[18,26-27]。植物盖度能够表征植物的横向分布、影响土壤浅层水分[28],是表征高寒草地退化程度的重要指标。因此,本研究采用郑元润[29]和王鲜鲜等[30]改进的稳定性计算方法,首先把样方内植物的盖度转换成相对盖度;其次,将植物的盖度与种对应,按照盖度值从大到小排列;然后将群落内相对盖度与物种总数的倒数按顺序分别累加;最后以累积相对盖度为y,以累积物种数为x,建立一条公式为y=ax2+bx+c 的平滑曲线,绘制出y=100-x直线,两条线的交点与稳定点(20,80)之间距离越近则表示群落越稳定,即两点之间欧氏距离与稳定性呈反比。本研究取欧式距离的倒数并乘以100%作为Godron 指数来表征群落稳定状态。
1.4 数据处理
在国家气象科学数据中心(https://data.cma.cn/)下载研究区域2001-2020年的国家气象站数据,在软件Arcmap 10.2 中,运用普通克里金插值[31]的方法对年均降水量(mean annual precipitation,MAP)和年均温度(mean annual temperature,MAT)进行插值分析,通过栅格提取获得采样点的气象数据。采用单因素方差和最小显著差数法(LSD)分析4 种类型草地的植被因子和土壤因子之间的差异以及不同草地类型不同功能群地上生物量和物种数的差异;运用回归分析揭示不同草地类型群落稳定性与多样性之间的线性关系;选用结构方程模型分析气候、土壤、植物多样性与群落稳定性之间的关系,选用比较拟合指数[compare fitted indices(CFI)>0.9]、渐进残差均方和平方根[root mean square error of approximation(RMSEA)<0.08]、标准均方根残差[standardized root mean square residual(SRMR)<0.05]、显著性概率值(P>0.05)等指标对模型优度进行评价,选出最优模型。最终选用均匀度指数、稳定性指数、MAP、pH、NH4+-N、SOC、TP 构建结构方程。结构方程模型在Amos 21.0 中建立,其他的分析与绘图均在R 3.6.2 中进行。
2 结果与分析
2.1 不同草地类型植物群落结构特征
不同草地类型植被群落特征存在显著差异(图2)。地上生物量和群落总盖度由高寒湿地草甸、高寒草甸、高寒草原到高寒荒漠草原显著下降(P<0.05)。高寒荒漠草原物种丰富度指数和Shannon-Wiener 多样性指数均显著小于其他3 种草地类型(P<0.05),而其均匀度指数则相反。高寒草原3 种多样性指标均处在最高水平,但其稳定性指数较低。高寒湿地草甸和高寒草甸3 种多样性指数均没有显著差异。高寒草甸丰富度最高,稳定性显著高于其他3 种草地类型(P<0.05)。

图2 不同草地类型植被群落特征、多样性和稳定性Fig.2 Vegetation community,diversity and stability characteristics of different grassland types不同字母表示不同草地类型间差异显著(P<0.05),下同。Different letters indicate significant differences among different regions(P<0.05),the same below.
由图3 可知,4 种草地类型不同功能群地上生物量和物种数差异显著(P<0.05)。莎草科地上生物量和物种数从高寒湿地草甸、高寒草甸、高寒草原到高寒荒漠草原显著下降(P<0.05);高寒草原禾本科的地上生物量和物种数以及豆科的物种数均显著大于高寒湿地草甸和高寒荒漠草原(P<0.05)。高寒草甸杂类草地上生物量和物种数均显著大于高寒草原和高寒荒漠草原(P<0.05)。由图4 可知,由高寒湿地草甸和高寒草甸到高寒荒漠草原和高寒草原莎草科相对地上生物量和物种数均减少,禾本科和豆科均增加;杂类草相对地上生物量增加,相对物种数大小排序为高寒草甸>高寒草原>高寒荒漠草原>高寒湿地草甸。综上所述,由高寒湿地草甸和高寒草甸到高寒草原和高寒荒漠草原,主要功能群由莎草科逐渐转变为杂类草和禾本科。
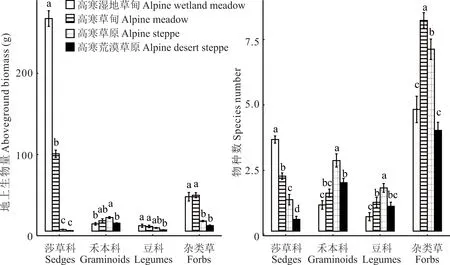
图3 不同草地植被群落功能群地上生物量和物种数Fig.3 Aboveground biomass and species number of functional communities of different grassland vegetation communities

图4 不同草地植被群落功能群地上生物量和物种数相对比例Fig. 4 Relative proportions of aboveground biomass and species number in functional communities of different grassland vegetation communities
2.2 不同草地类型环境因子变化
由图5 可知,各环境因子在4 种草地类型间均有显著差异。MAP 表现为高寒草甸>高寒湿地草甸>高寒草原>高寒荒漠草原(P<0.05),变化范围为88.95~540.22 mm;pH 在4 种草地类型间显著下降(P<0.05),变化范围为5.80~10.00;2 种草甸的NH4+-N、TP 和SOC 均显著低于2 种草原(P<0.05),其变化范围分别为5.64~27.54 mg·kg-1、0.24~8.53 g·kg-1和2.27~92.90 g·kg-1;高寒湿地草甸的AP 和NO3--N 均显著低于2 种草地类型,其变化范围分别为1.26~10.62 mg·kg-1和1.81~34.08 mg·kg-1。

图5 不同草地类型间环境因子的差异Fig.5 Differences of environmental factors among different grassland types·代表异常值Present abnormal value. MAP:年均降水量Mean annual precipitation;NH4+-N:铵态氮Ammonium nitrogen;TP:全磷Total phosphorus;SOC:土壤有机碳Soil organic carbon.下同The same below.
2.3 高寒草地植物多样性与群落稳定性的关系及其影响因素
回归分析结果表明(图6),群落稳定性与Pielou 均匀度指数在4 种草地类型中均呈现显著负相关(P<0.05),拟合优度分别为0.900、0.087、0.859 和0.621;在高寒草甸和高寒荒漠草原中与Shannon-Wiener 多样性指数呈现显著负相关(P<0.05);在高寒湿地草甸和高寒草原中与物种丰富度指数呈显著正相关(P<0.05)。综上所述,不同草地类型植物多样性与群落稳定性关系不同,但大体表现为提高物种丰富度能够增强群落的稳定性,提高物种均匀度和多样性能减弱群落的稳定性。
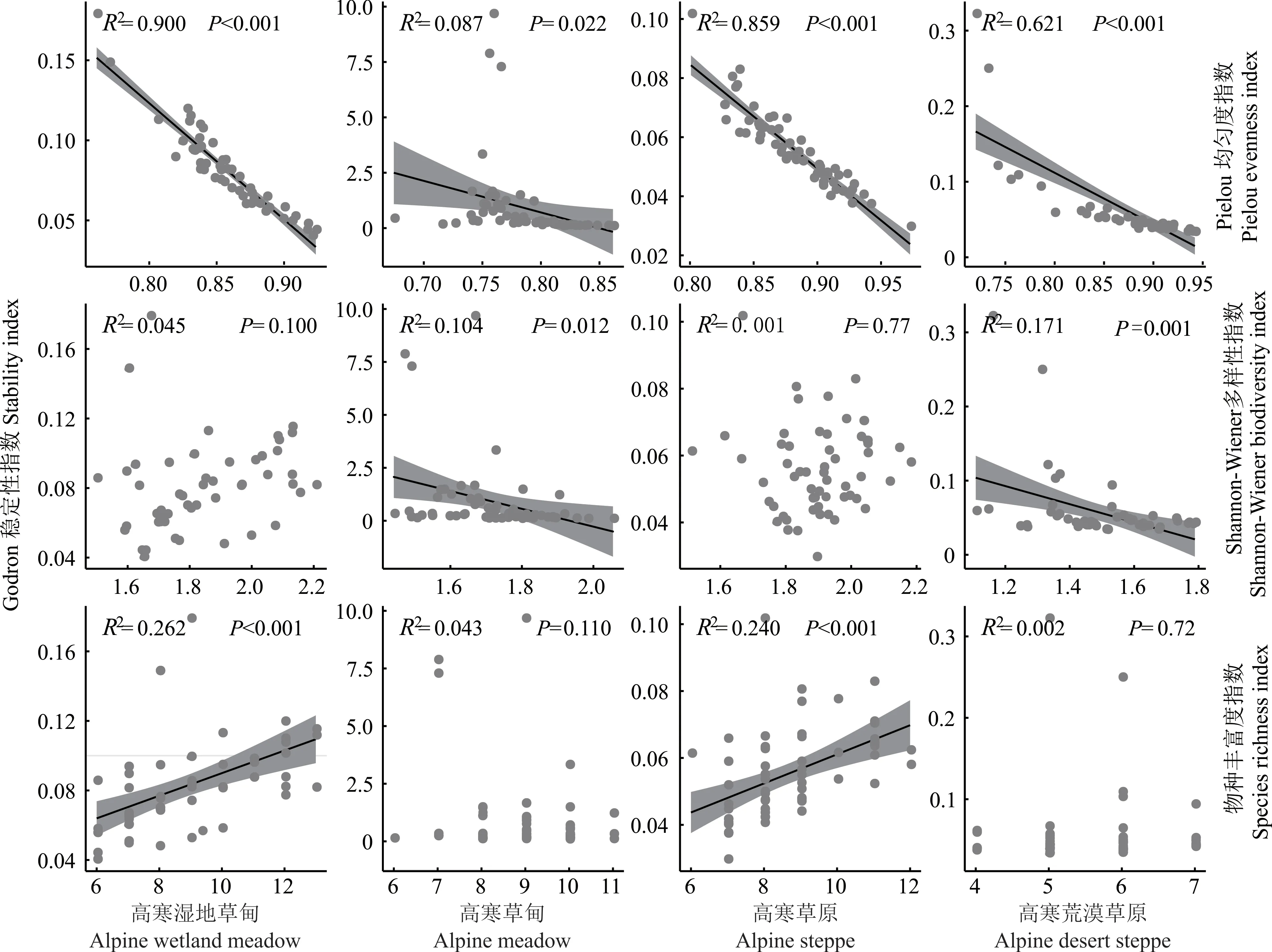
图6 不同草地类型植物多样性与群落稳定性线性回归分析Fig.6 Linear regression analysis of plant diversity and community stability of different grassland types
高寒草地植物多样性与群落稳定性的驱动机制如图7 所示。该模型中,MAP 和NH4+-N 对植物多样性有显著的直接影响,其直接效应分别为-0.457、0.129;pH 和SOC 通过NH4+-N 对植物多样性有显著间接影响。此外,MAP 和Pielou 指数对群落稳定性有显著直接影响,其直接效应为0.317 和-0.262;NH4+-N 和MAP 通 过 植物多样性对群落稳定性有显著的间接影响。综上所述,高寒草地植物多样性的主要影响因子为MAP 和NH4+-N,MAP 和植物多样性是群落稳定性的主要影响因子。
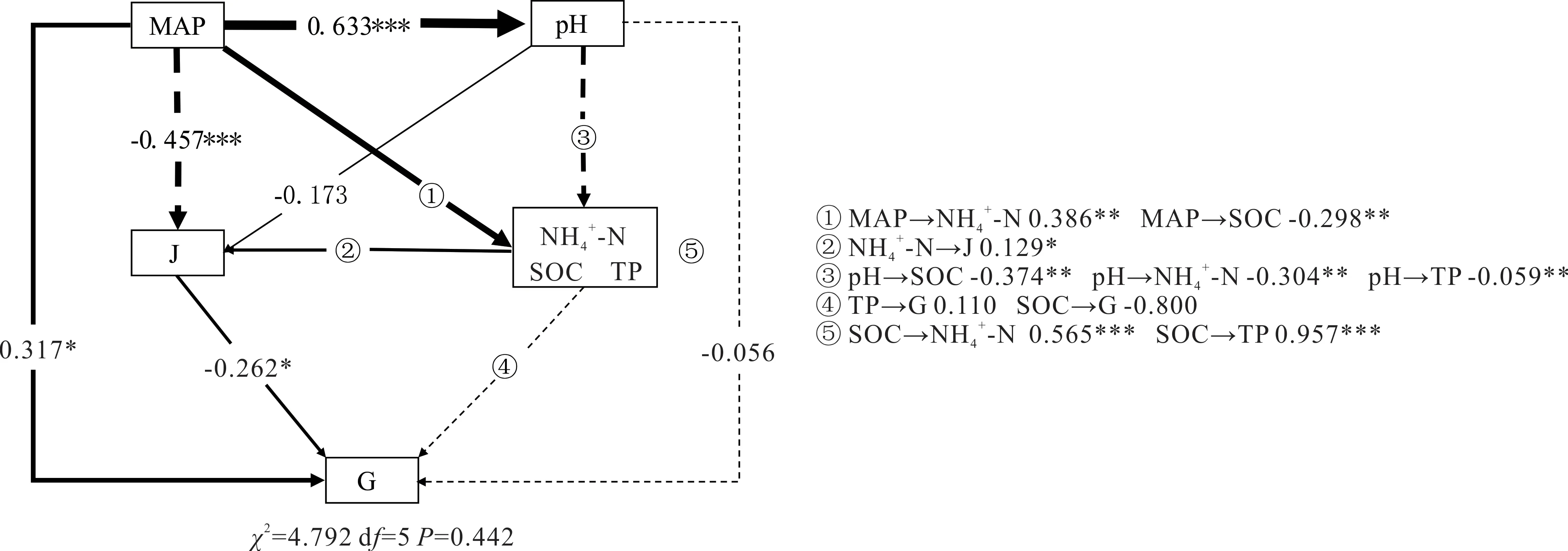
图7 高寒草地植物多样性和群落稳定性驱动因素Fig.7 Driving mechanism of plant diversity and community stability in alpine grassland实线和虚线分别表示正作用和负作用,箭头的宽度与作用强度呈正比,箭头上的数字是标准化路径系数,***,P<0.001;**,P<0.01;*,P<0.05。J:Pielou 均匀度指数Pielou evenness index;G:Godron 稳定性指数Godron stability index.The solid line and the dotted line represent the positive and negative effects respectively. The width of the arrow is proportional to the action intensity. The number on the arrow is the standardized path coefficient.
3 讨论
3.1 不同草地类型下植物群落的差异
有研究表明[32],高寒草地地上生物量呈现出高寒草甸>高寒草原>高寒荒漠草原的趋势。本研究结果与其相符(图2)。然而本研究中高寒湿地草甸年均降水量小于高寒草甸,地上生物量却大于高寒草甸。这可能是由于高寒湿地草甸多处在河流湖泊周围,由于河流湖泊水分的补充,湿地土壤含水量较高,因而高寒湿地草甸植物的可利用水分高于高寒草甸,植被的地上生物量也高于高寒草甸,这说明土壤水分也是影响高寒草地地上生物量的重要的环境因子[28]。
本研究发现,不同草地类型物种丰富度呈现高寒荒漠草原显著小于其他3 种草地类型(图2)。这可能是因为不同类型生境的草地中,生境不同的植物所占比例不同导致的[33]。高寒荒漠草原绝大部分为旱生植物,中生植物少于其他3 种草地类型。此外,本研究发现植物的均匀度指数表现为草甸低于草原,这可能是因为土壤养分对物种均匀度的分布有着重要的影响。土壤作为维持陆地生态系统的重要载体,在转移营养和水分运转、储存、调控与外界气体的交换等方面发挥着重要作用[34-35]。曾思金等[36]研究发现土壤有机碳与Pielou 物种均匀度指数显著正相关,本研究结果与其相符(图7)。本研究中,由于高寒草甸和高寒湿地草甸地上生物量较大,植被茂盛,对土壤营养物质的消耗较多,土壤氮素和有机碳的含量较低,这导致了其物种均匀度低于高寒草原和高寒荒漠草原。
本研究中,高寒草甸群落稳定性大于其他3 种草地(图2),这可能是由于不同草地类型物种的优势度不一样导致的。优势度效应认为优势种的稳定性对群落稳定性有重要作用[37]。群落组成物种的波动性越小,则群落的方差之和就越少,进而降低了群落的波动性,使群落稳定性更高[38]。研究表明优势种比其他物种稳定性更高,优势物种的优势度增加会使群落更加稳定[39]。此外,本研究发现从高寒湿地草甸到高寒荒漠草原,占优势地位的功能群从莎草科逐渐变为杂类草和禾本科。这可能与土壤水分有关。水分含量较高时,莎草科占据了大量的生态位,在对光和养分的竞争中处于优势地位,抑制了杂类草和禾本科植物的生长[40];而水分较少时,耐旱的禾本科和杂类草植株发展壮大。
3.2 植物多样性与群落稳定性关系
本研究中,不同草地类型植物多样性与稳定性关系不同,同一草地类型不同多样性指数对群落稳定性影响也不同,总体表现出均匀度的提高降低了群落稳定性(图6),这可能与群落抗干扰能力有关。Hillebrand 等[41]研究表明均匀度的提高会提高群落的恢复力,降低抗干扰能力。而本研究的样地处于生态环境脆弱、对外界刺激响应敏感的高寒地区,群落演替时间较长,一旦被破坏短时间内难以恢复,群落的抗干扰能力对稳定性的影响更大,因而均匀度与稳定性呈现负相关关系。另外,本研究发现物种丰富度指数与群落稳定性呈正相关关系,这与胡冬等[19]的研究结果一致。由于受到物种间相互作用对环境波动的放大效应和物种对环境波动的敏感性的影响,生物多样性能够增加物种对环境波动响应的多样性,从而使群落的稳定性增加。多样性指数是把物种丰富度与均匀度结合起来的统计量,多样性信息度量、概率度量和几何度量都是以不同的方式表达物种丰富度与均匀度的结合[42]。本研究中Shannon-Wiener 多样性与群落稳定性呈负相关关系,这可能是因为高寒草甸中均匀度与群落稳定性的拟合程度更好,对稳定性的影响高于物种丰富度。
3.3 环境因子对植物多样性和群落稳定性的影响
降水量能够在不同程度上影响植物群落结构、生物多样性和稳定性。高寒草地生态系统对气候变化敏感,尤其是降水的变化会引起植物水分可利用性在时间空间上的差异,导致植物种内、种间关系的变化[43],进而引起群落稳定性的变化。本研究发现降水能够通过直接和间接作用促进高寒草地植物多样性和群落稳定性提高,其原因可能是降水能够促进土壤中种子的萌发、加速土壤有效养分的溶解和流动[44]、提高植物对养分的吸收效率[45],提高物种多样性从而提高群落的稳定性。土壤养分对于植物多样性起着至关重要的作用[11],本研究发现土壤中的NH4
+-N 对植物多样性和群落稳定性有显著影响。这是由于高寒草地氮素含量较高,导致其他营养元素的相对不足,抑制植物丰富度的增加[44],进而降低了群落的稳定性。另外,有研究表明SOC 和pH 对植物多样性有直接影响[46],而本研究发现SOC 和pH 通过影响NH4+-N 间接影响植物多样性。这可能是因为本研究地域跨度大,各草地类型之间植物组成不同,对所处生境资源的需求、能力、方式等均有差异,导致环境因子与植物多样性并未产生直接效应[47]。
4 结论
高寒草地不同草地类型植被群落特征差异显著;高寒草原植物多样性显著高于其他3 种草地类型,高寒草甸群落稳定性最高;植物多样性和群落稳定性的关系并不相同,但大体表现为提高物种丰富度能够增强群落的稳定性,提高物种均匀度和多样性能减弱群落的稳定性。降水量、土壤有机碳、pH 以及铵态氮含量是影响高寒草地植物多样性和群落稳定性的主要因素。在高寒草地生态系统的管理过程中,应关注气候和土壤因子变化,根据不同草地类型制定不同的管理措施。
猜你喜欢
儿童故事画报·自然探秘(2024年5期)2024-05-22 22:26:58
作物研究(2023年2期)2023-05-28 13:44:14
疯狂英语·新读写(2023年4期)2023-05-10 10:44:22
青海草业(2022年2期)2022-07-23 09:34:58
今日农业(2020年19期)2020-12-14 14:16:52
河南畜牧兽医(2020年21期)2020-01-10 00:20:08
活力(2019年21期)2019-04-01 12:17:10
纺织科技进展(2015年1期)2015-11-28 05:56:28
中国科技信息(2015年2期)2015-11-16 08:18:32
植物营养与肥料学报(2014年1期)2014-03-11 20:24:38
