
Mitochondrial cytochrome oxidase 1 reveals genetic diversity of the African Snakehead fish Parachanna obscura, Gunther, 1861 from Nigeria’s freshwater environment
2023-03-20 07:39FridyElijhOshoBmideleOluwrotimiOmitoyinEmmnuelKolwoleAjniVitorAzuhAdedpoOlutolAdediji
Aquaculture and Fisheries 2023年1期
Fridy Elijh Osho, Bmidele Oluwrotimi Omitoyin, Emmnuel Kolwole Ajni,Vitor O.Azuh, Adedpo Olutol Adediji
aDepartment of Aquaculture and Fisheries Management, University of Ibadan, Ibadan, 200284, Nigeria
bVirology and Molecular Diagnostic Unit, International Institute of Tropical Agriculture (IITA), Ibadan, 200001, Nigeria
cDepartment of Crop Protection and Environmental Biology, University of Ibadan, Ibadan, 200284, Nigeria
Keywords:
Nucleotide composition
Haplotype diversity
Molecular variance
Phylogenetic analysis
Topology
A B S T R A C T
The study investigated the genetic variation of Parachanna obscura from five rivers (Anambra, Ibbi, Imo, Katsina-Ala and Ogun) in Nigeria using the mitochondrial cytochrome oxidase 1 gene.DNA was extracted from 19, 22,16, 18 and 21 fin clips per river population, respectively and subjected to polymerase chain reaction.A total of 96 sequences, each with 671 bp were obtained with 38 (5.6%) polymorphic, 27 (3.8%) parsimoniously informative and 659 (98.2%) conserved sites.Mean nucleotide composition was C =28.07%, T =29.43%, A =22.18%, G =20.32%.A total of 40 haplotypes with 38 unique sequences as well as 24 substitutions with 22 transversions and two transitions were obtained.Nucleotide diversity among populations ranged from 0.00184 to 0.00888 representing Ibbi and Imo, respectively while haplotype diversity ranged from 0.77056 to 0.95000 also, from Ibbi and Imo, respectively.Analyses of molecular variance showed that the intra-population variation accounted for 50.05%.Topology from phylogenetic analyses revealed that P.obscura from Imo River was distinctly different from the rest.
1.Introduction
Fish production from capture fisheries has been unstable and on the decline in recent years, resulting in increase in poverty and food insecurity (Ajani & Osho & Usman, 2019).Africa has natural endowment for fish production but produces only 2.7% of global aquaculture production while per capita fish consumption has been static and even decreasing (FAO, 2020).Consequently, sizeable share of fish consumed in developing countries consists of imports, and, this is expected to grow over the years owing to increasing demand (FAO, 2012).Increasing aquaculture output therefore, remains the best option to boost supply and bridge the gap between fish demand and supply.One major setback to aquaculture production in Africa is the limited number of species that have been successful cultured to commercial success (FAO, 2020).The diversification of aquaculture species and culture models demonstrates a clear trend towards achieving food security (Li, 2015).Thus, there is the need to investigate culturable species that exist within the continent’s territorial waters.
The assurance of a more resourceful and sustainable use of natural resources is crucial to safeguarding food and nutrition security.A better understanding of available resources and how they match current and future needs is crucial in confronting these challenges.A good knowledge of the status of resources and their inclinations as a consequence of their exploitation are important steps towards their sustainable management.Some declining species likeParachanna obscura,which reveal interesting aquaculture potentials, have been identified.It is a choice fish across many African countries.However, poor harvesting methods have affected its availability even in the wild and has become endangered (IUCN, 2017).Efforts to culture this fish in Nigeria has remained experimental with little success recorded due to poor understanding of the natural populations where the broodstocks and seeds are taken(Kpogue et al., 2013; Osho et al., 2020).
Researches into the domestication ofP.obscurainclude investigation into it is biology (Ama-Abasi & Affia, 2010), parasitic helminths infections (Osho, 2017), haematology (Adebayo et al., 2007), length weight relationship and fecundity (Osho & Usman, 2019).However,there is dearth of information on the genetic diversity of the natural populations in Nigeria that form the gene pool for its domestication.The careful selection of suitable natural stocks, based on genetic knowledge,can provide greater prospects for success in aquaculture production,species-recovery and sustainable management of natural populations(Quattro & Vrijenhoek, 1989).Therefore, this study sought to understand the genetic diversity ofP.obscurafrom Nigeria’s hydrologic areas using the mitochondria cytochrome oxidase 1 gene.
2.Materials and methods
2.1.Study sites and experimental design
All 12 River Basin Development Authorities (RBDAs) of Nigeria:Anambra-Imo, Lower Niger, Upper Niger, Benin-Owena, Chad, Cross river, Hadejia-Jama’are, Lower Benue, Niger Delta, Ogun-Osun, Upper Benue, and Sokoto-Rima were sampled for this study (Ezenweani,2017).Two stage sampling techniques (EPA, 2002, p.166) was adapted for the collection ofP.obscurasamples across the 12 RBDAs in Nigeria.All RBDAs were purposively selected while one river in each of the basins was randomly chosen as sampling site for collection of the fish samples from fishermen catch.These were rivers Anambra, Imo, Ibbi,Kaduna, Katsina Ala, Hadeija, Ogun, Sokoto, Great Kwa, Niger, Kaduna,Ovia, Lake Chad (representing Anambra-Imo, Niger Delta, Upper Benue,Lower Benue, Hadejia-Jama’are, Ogun-Osun, Sokoto-Rima, Cross river,Lower Niger, Upper Niger, Benin-Owena, Chad river basins as shown in Fig.1.

Fig.1.Map of Nigeria showing sampled rivers and collection points.
2.2.Sample collection
Live samples ofP.obscurawere collected from the fishermen catch,who used Malian traps bimonthly for 18 months.Samples were identified to species level using guide by Olaosebikan and Raji (2013) and Froese and Pauly (2018) and subsequently transported to the laboratory in oxygenated plastic aquaria.
2.3.Nucleic acids extraction and amplification of cytochrome oxidase 1 gene
Clips from the caudal fin of eachP.obscurawere taken and placed in microcentrifuge tubes.Nucleic acid extraction was carried out via the Wizard® SV Genomic DNA Purification System (Promega, Madison, WI,USA), according to manufacturers’ protocol by using approximately 25 mg of homogenized caudal fin clips (Ward et al., 2005).To investigate genetic diversity among the population, the primers FishF1 5′-TCAACCAACCACAAAGACATTGGCAC-3′and FishR1 5′-TAGACTTCT GGGTGGCCA AAGAATCA-3’ (Ward et al., 2005 was used to amplify the~700 bp COI gene using polymerase chain reaction (PCR).
2.4.Data analysis
The Chromas 2.4.4 (Technelysium) software was used to view the sequences and chromatograms produced.Sequences were end-trimmed via BioEdit v7.2.5 (Hall et al., 2013).The edited sequences were inspected using the Basic Local Alignment Search Tool implemented in National Centre for Biotechnology Information (NCBI).All sequence readings were checked for possible contaminations and species identities by comparing their applicable similarities with the previous sequences of same species available in the system.Subsequently, multiple alignments of the sequences were conducted through ClustalX software version 2.0.10 (Larkin et al., 2007).Haplotype generation was done by exporting aligned sequences to an online fasta sequence toolbox(Villesen, 2007).The pairwise genetic distance between haplotypes and phylogenetic analysis were done using the neighbor joining method within MEGA v4.0 (Tamura et al., 2007).Pairwise identities were performed using SDT v1.2 (Muhire et al., 2014) with pairwise gap deletions to confirm haplotype frequencies.The Fixation Statistic (Fst), showing the level of population subdivision (Hudson et al.,1992) between populations together with the Chi-square probability test for population differentiation with 1000 permutations of the datasets were also estimated using DnaSP 5.1.Analysis of molecular variation was calculated on Arlequin 3.5.
3.Results
3.1.Fish availability
Significant amount ofP.obscurawere only encountered in Rivers Anambra, Ibbi, Imo, Katsina-Ala and Ogun.The fish was not encountered in Rivers Kaduna and Hadejia while the catch was scanty in Rivers Niger, Great Kwa, Ovia and Sokoto.Lake Chad could not be assessed due to theBoko Haraminsurgency in that region of Nigeria.The partial COI sequences were deposited at the NCBI data bank with accession numbers MT857805 to MT857900.
3.2.Mitochondrial DNA sequence variations
A total of 96P.obscuraCOI individual sequence data were obtained,Rivers Anambra, Ibbi, Imo, Katsina-Ala and Ogun with 19, 22, 16, 18 and 21 sequences, respectively.After alignment and trimming, the final sequence length was 671 bp of which 38 (5.6%) polymorphic sites, 27(3.8%) parsimoniously informative sites and 659 (98.2%) conserved sites were identified.The mean total nucleotide composition was C =28.07%, T =29.43%, A =22.18%, G =20.32% (Table 1).The nucleotide distribution percentages did not differ significantly (p>0.05)among the populations ofP.obscura.

Table 1Percentage MtDNA sequence nucleotide composition of Parachanna obscura from Nigeria freshwater environment.
From the 96 sequences analysed, 40 haplotypes were distinguished(Table 2) with 38 unique and two shared sequences among the populations.Overall, 24 substitutions were found among the haplotypes,including 22 transversions and two transitions.P.obscurahaplotypes 1,3 and 4 were the most common and found in four out of five populations sampled, being present in all but River Ogun population.Haplotype with the highest number of individuals wasP.obscura1 (n =13), representing four populations (Anambra, Ibbi, Imo and Katsina-Ala).Anambra (13) and Ibbi (6) populations had the highest and lowest numbers of haplotypes, respectively.The nucleotide diversity among populations ranged from 0.00184 (Ibbi) to 0.00888 (Imo).On the other hand, the haplotype diversity ranged from 0.77056 to 0.94737 in all populations,with the least and highest values found in the Ibbi and Imo populations,respectively.Some haplotypes were shared among different populations were some were unique.To further consolidate the data on haplotype frequencies and rule out the presence of cryptic species, pairwise analyses was performed using SDT and revealed high similarity between theP.obscurapopulations from River Imo and other populations in the study (Supplementary Fig.1).The populations from River Imo had at least 97.16% nucleotide similarity with others within Nigeria while populations from River Ibbi were totally homogenous at 100% (Table 3).

Table 2Distribution of 40 observed haplotypes, nucleotide diversity, number of haplotypes, haplotype diversity and number of polymorphic sites among populations of Parachanna obscura from Nigeria freshwater environment.

Table 3Percentage pairwise similarities among the mtCOI sequences of Parachanna obscura populations from Nigeria freshwater environment.
Mismatch distribution of the expected and observed pairwise genetic diversity of each studied population and the combined 40P.obscurahaplotypes are presented in Fig.2.The graphs showed marked differences between the expected and observed pairwise genetic relationship for all the populations and the combined.A multimodal mismatch distribution was observed for the Anambra Ogun, Katsina-Ala and the combined population while unimodal mismatch distribution was observed for the Ibbi river population.A bimodal graph was observed for the mismatched distribution of the Imo river population (Fig.2).TheFstindex of Anambra is 0.10918, 0.07859, 0.00015 and 0.52015 related to populations from Ibbi, Imo, Katsina-Ala and Ogun river populations,respectively (Table 4).TheP.obscurapopulation from Ibbi was also separate from those of Imo, Katsina-Ala and Ogun populations withindices of 0.77084, 0.17208 and 0.00667 respectively.The analyses of molecular variance (AMOVA), as shown in Table 5, revealed that the within population variation (50.05377%) was slightly higher than the among populations sources of variation (49.94623%).

Fig.2.Mismatch distribution of P.obscura from Nigeria Freshwater Environment.P.obscura populations from Anambra (A), Ibbi (B), Imo (C), Katsina-Ala (D), Ogun(E) and the whole population combined (F).
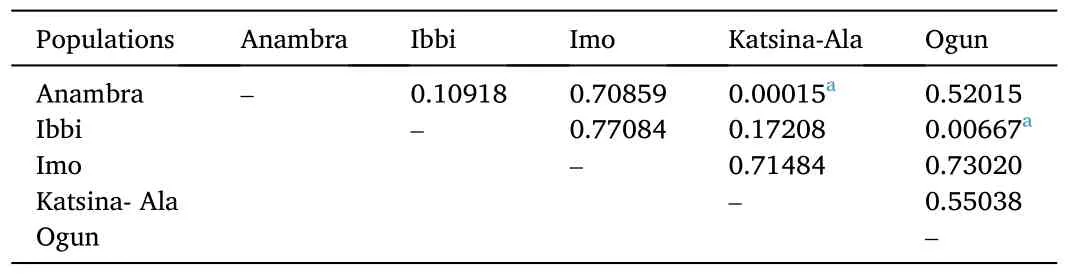
Table 4Fixation statistics for population differentiation based on 1000 permutations of the sequence dataset of Parachanna obscura from Nigeria freshwater environment.
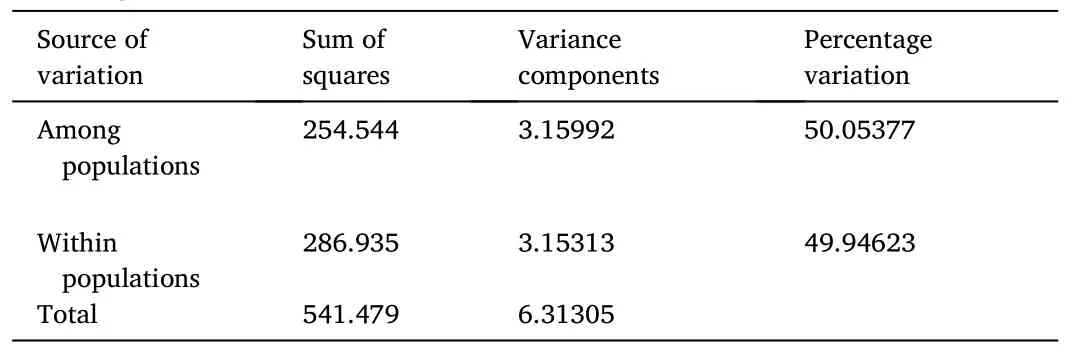
Table 5Analysis of Molecular Variance (AMOVA) for haplotypes of Parachanna obscura from Nigeria freshwater environment.
3.3.Phylogenetic relationships among haplotypes of P.obscura from the freshwater environment of Nigeria
The phylogenetic molecular analyses of the combined haplotypes ofParachanna obscurafrom the freshwater environment of Nigeria using the neighbor joining algorithm is depicted in Fig.3.HaplotypesP.obscura21, 24, 27, 20, 22,25, 26, 23, 28, which comprised individuals from the Imo River population, formed a distinct cluster from the rest.The others from the remaining four populations showed a markedly different cluster from this group.

Fig.3.Neighbor joining phylogenetic tree showing relationships among the haplotypes of P.obscura populations in Niegrian freshwater using the mitochrome cytochrome oxidase I gene.
This later group formed two major clusters which, as a minor cluster,comprised haplotypesP.obscura38, 36, 33, 30, 37, 35, 32, 31, 29, 34 and was different from the remaining.These haplotypes containing individuals from the Ogun River populations exhibits various levels of minor groupings within them.The remaining populations comprising Anambra, Ibbi and Katsina-Ala showed more levels of similarities by forming a separate cluster.HaplotypesP.obscura8,1,13,11, 5, and 6 which comprised individuals from Anambra, Ibbi and Katsina-Ala formed a cluster, the small in-between haplotypes wereP.obscura4,19, 39 and 10 which comprised the three populations and one Ogun individual while the remaining haplotypes formed an independent cluster.
3.Discussion
The mean total nucleotide composition in descending order of was T>C>A>G.This general pattern of distribution was observed in all the populations.This shows that theP.obscurain all the studied populations have strand bias for T.Chen et al.(2016) documented relative nucleotide frequencies in tandem with one obtained in the present study when they recorded base compositions of similar patterns (T>C>A>G) forBrama japonica.A related trend of nucleotide composition was also recorded by Lakra et al.(2011), who noted overall relative nucleotide composition of T>C>A>G in six genera of Scombrids and seven species of Groupers.The current study, however, contrasts to the percentage base composition of A>G>T>C forChanna striatafrom Thailand (Boonkusol & Tongbai, 2016).Likewise, Persis et al., 2009,revealed nucleotide frequencies T>A>C>G in the mtDNA CO1 of Carangid fishes of India.All of these showed variations in the nucleotide compositions of the mitochondrial genomes within different fish species.Observed differences could be as a result of the fact that the COI locus undergoes a high mutational rate even for mtDNA (Saccone et al., 1999).
Half of the haplotypes found in the Ibbi Populations were also found in the Anambra and Katsina-Ala populations.In addition, the Anambra River population had many unique haplotypes which it did not share with the other two populations.This reveals that there are probably current gene flows from the Ibbi and Katsina-Ala to the River Anambra population.The fact that up to half of Ibbi river population haplotypes were also found in the Anambra populations while the Anambra populations still retained a lot of their own unique haplotypes indicated that the gene flow maybe from the Ibbi populations to Anambra and not necessarilyvice versa.The existence of shared haplotypes at more than one locality denotes that genotype arising at one locality spread rapidly to other localities (Lankford et al., 1999).Kornfield and Bagdanowicz(1987) also deduced trends of gene flow from spreads of unique Mitochondrial DNA haplotypes and their supposed predecessors and thus predicted that under restricted gene flow, same unique haplotypes should occur in the populations as their precursors.The presence of unique haplotypes to the Imo and Ogun populations suggest absence of regular or recent gene flow between each of these populations and other populations.The observed patterns of mtDNA haplotype variations in all the populations shows that they do not comprise a single breeding stock(Gold & Richardson, 1998).This is reflected in the high number of haplotypes (40) obtained from the study.These observed patterns of variation may also reflect historical rather than present day gene flow.O’Brien et al.(1987), suggested that low haploype diversity in groupers showed isolated populations of small effective size recently diverged from a population that possessed low levels of genetic variations.
Nucleotide diversity values revealed that the various populations demonstrated different degrees of genetic variability, with the Rivers Imo and Ibbi populations recording highest and least genetic variability,respectively.Meanwhile, low genetic diversities are likely outcomes of inbreeding and a limited number of founder stocks in a population (Wu and Yang, 2012).Minimizing the mating of closely related individuals has been suggested to be very helpful for maintaining the genetic diversity of a population (Martins et al., 2003).Breeding methods inevitably influence the genetic variability of populations.Similar results have also been observed in the kin relationships of parents’ match experience ofOreochromis esculentus(Agnese et al., 1999).Uncontrolled and continued extraction of specimens from nature could dramatically reduce effective population size, increasing the risk of losing genetic diversity by genetic drift (Garcia-Cisneros, 2016).
Generally, the haplotype and nucleotide diversities of theP.obscurafrom all the sampled populations were high and this showed that they have not experienced severe bottleneck recently.Sato et al.(2004)similarly recorded variated haplotype and nucleotide diversities from the mitochondrial DNA sequences of populations of chum salmon from the Pacific Rim.Boonkusol and Tongbai (2016) also documented high haplotype and nucleotide diversities for the striped Snakehead Fish,Channa striatain River Basin of Central Thailand.
Demographic changes occur in effective population size over time with signatures shown as distribution of allelic frequerncies.The mismatch distribution is a frequency graph of pair wise differences between alleles.A multimodal mismatch distribution was observed for the Anambra, Ogun, Katsina-Ala and the general/combined population while unimodal mismatch distribution was observed for the was observed for the Ibbi river population.A bimodal graph was observed for the mismatched distribution of the Imo river population.It is usually multimodal in samples drawn from populations at demographic equilibrium while it is unimodal in samples drawn from populations that have passed through populations that has gone through a recent expansion (Rogers & Harpending, 1992, Wang et al., 2008) or through a range of expansions with high levels of migration between neighbouring demes (Excoffier, 2004).
Analysis of molecular variation revealed that sources of variation for the mitochondrial DNA were the components of variations within each populations ofP.obscuraand the variations due to the interactions among the populations.Though each of the two sources of variations contributed closely to the observed variations (50.05377% and 49.94623% respectively), the within population contributed more,suggesting a panmictic populations (Grünwald & Hoheisel, 2006).Intra-species population genetic diversity has been conventionally evaluated using departures of allele frequencies from panmictic expectations (Ramo et al., 2016).When a species exhibits subdivision, there will be expected increase haplotype diversity and higher number of segregating sites for genomes sampled from different demes (Slatkin,1987).Excoffier et al.(1992) also suggested that application of AMOVA to human mtDNA haplotype data shows that population subdivisions are better resolved when some measure of molecular differences among haplotypes is introduced into the analysis.These gene flow indices give direct comparison between populations to know the level of genetic interactions between them.
Decline in heterozygosity caused by subdivision within a population are usually quantified using the fixation index.This fixation index ranges from 0 (showing no differentiation between the overall population and its subpopulations) to a hypothetical maximum of 1.In this study,P.obscurapopulations from Rivers Katsina-Ala and Anambra were the most closely related population.They had the lowestFstvalues of 0.00015, depicting that these populations have recent and/or current breeding history.This is in tandem with the shared haplotpyes by these populations.Populations from Rivers Ibbi and Anambra also showed evidence of similarity akin to the pattern of the Katsina-Ala and Anambra populations.These two phenomena could be as a result of flow of water shed from Upper and Lower Benue hydrologic areas to the Anambra hydrologic area.The Katsina-Ala and Ibbi populations pairings exhibited the fourth closest gene flow.These two rivers are in regions that are closely located (Lower and Upper Benue respectively) and there are incidences of annual floods in these areas that may allow the two populations to mix and therefore encourage gene flow.Boonkusol and Tongbai (2016) also found high genetic variability and high gene flow between regional populations ofChanna striatafrom river basin of central Thailand due to fish dispersal by flood and aquaculture.
However, Anambra and Imo which were geographically close showed very low level of mixing and gene flow while Imo and Ibbi had the highest value ofFst, indicating the least gene flow between those populations.The topology from the phylogenetic analysis revealed that the Imo river population was markedly and distinctly different from theP.obscurafrom the other populations.This is evident in their formation of a completely different cluster from the rest of the populations.This could infer that this population has been isolated for long and there have been little or no gene flow with the other populations.The other haplotypes, which consisted of the remaining four populations showed a markedly different cluster from this group.Among these populations,the Ogun river population also form a subcluster cluster.However, the remaining populations comprising Anambra, Ibbi and Katsina-Ala extractions showed increased levels of similarities, indicating a more recent gene flow among them.Congeneric species always clustered together and in most cases so did the co-familial species (Lakra et al.,2011).This phylogenetic results is also in tandem with that of the fixation statistics earlier discussed.Phenotypic analysis of the populations revealed well conserved characteristics (data not shown) and confirmed the out come of the molecular analysis.
4.Conclusion
The present study revealed thatP.obscurafrom Nigeria’s freshwater environment have bias for the T nucleotide.The study also revealedP.obscurapopulations from Rivers Imo and Ibbi populations have the highest and least genetic variability, respectively.Phylogenetic analysis revealed that the River Imo population was markedly and distinctly different from theP.obscurafrom the other populations.The findings in the current study provides useful guide to resource managers in their conservation and breeding efforts in order to achieve optimum utility on a sustainable basis.
CRediT authorship contribution statement
Friday Elijah Osho: Conceptualization, Data curation, Formal analysis, Funding acquisition, Investigation, Methodology, Project administration, Resources, Software, Supervision, Validation, Visualization, Roles/Writing –; original draft, Writing – review & editing.Bamidele Oluwarotimi Omitoyin: Funding acquisition, Supervision,Validation.Emmanuel Kolawole Ajani: Conceptualization, Funding acquisition, Supervision, Validation, Methodology, Project administration, Resources, Visualization.Victor O.Azuh: Software, Visualization.Adedapo Olutola Adediji: Software, Visualization, Writing – review &editing.
Declaration of competing interest
All authors have no conflict of interest.
Acknowledgement
The authors appreciate the International Foundation for Science(IFS), Stockholm, Sweden for funding this research through their Grant Number I-2-A-6090-1 provided to Friday Elijah Osho to study the phenotypic and genetic characterization ofParachanna obscurafrom Nigeria’s freshwater environments.
Appendix A.Supplementary data
Supplementary data to this article can be found online at https://doi.org/10.1016/j.aaf.2021.08.007.
 Aquaculture and Fisheries2023年1期
Aquaculture and Fisheries2023年1期
- Aquaculture and Fisheries的其它文章
- Conservation aquaculture of Ompok bimaculatus (Butter catfish), a near threatened catfish in India
- Cloning, prokaryotic expression, purification, and functional verification of the insulin gene in black carp (Mylopharyngodon piceus)
- Effective CRISPR/Cas9-based genome editing in large yellow croaker(Larimichthys crocea)
- Effects of BDE-209 exposure on growth performance, intestinal digestive enzymes, and intestinal microbiome in common carp (Cyprinus carpio L.)
- Expression of mapk1 and egr1 genes in Onchidium reevesii under tidal stimulation
- Assessment of bactericidal role of epidermal mucus of Heteropneustes fossilis and Clarias batrachus (Asian catfishes) against pathogenic microbial strains