
五台山区不同海拔臭冷杉叶、枝、根的C、N、P化学计量特征研究
2023-03-13 09:21郭卫红胡砚秋郑庆荣李眉红
中国野生植物资源 2023年2期
郭卫红,胡砚秋,郑庆荣*,李眉红
(1.忻州师范学院,山西 忻州 034000;2.山西臭冷杉自然保护区管理局,山西 忻州 034000)
生态化学计量学是研究多重化学元素(主要是C、N、P)平衡和生物系统能量平衡的科学[1-2],因此生态化学计量学的研究可以将生物学科不同层次(分子、细胞、个体、生物系统等)的理论结果有机统一起来[3]。植物作为生产者,是陆地生态系统的重要组成部分[4]。植物不同器官的物质结构和功能属性不同,同一植物不同器官C、N、P元素含量也不相同[5]。研究单一物种不同器官C、N、P含量及生态化学计量特征,对揭示物种的生态策略和环境适应性具有重要的生态学和植物生理学意义[6]。海拔作为重要的综合性地形因子,通过影响光照、水分、植被和土壤质地等改变植物的功能性状、养分吸收和利用[7-10],致使植物不同器官C、N、P化学计量特征产生明显的适应性调节[11]。
目前,针对海拔梯度上植物叶片化学计量的研究逐渐增多,但结果不尽相同。例如,Müller等通过对喜马拉雅山脉植物的研究发现,随着海拔的升高,植物叶片N、P含量及N/P逐渐下降[12]。而杨思琪等[13]对天山中段植物的研究表明,天山南北植物叶N含量与海拔间无显著的线性关系,随着海拔升高,叶P含量升高,N/P降低。现有研究结果加深了对海拔梯度上植物叶片化学计量特征的认识,但其变化规律及影响因素仍有不确定性,且仅将叶片作为研究对象探讨其沿海拔梯度的变化,容易忽略海拔梯度植物其他器官化学计量特征的变化趋势是否存在一致性。研究海拔梯度同一物种不同器官化学计量特征的变化规律,对探讨植物养分分配及对环境的适应能力具有重要的意义。
臭 冷 杉[Abies nephrolepis(Trautv.) Maxim.]为松科冷杉属植物,常绿针叶乔木,喜冷湿环境[14],是山西省重点保护植物。学者们对其物种多样性[15]、种群现状及保护措施[16-17]等方面进行了研究,但鲜见对臭冷杉不同海拔、不同器官化学计量特征的研究。保护区所在的山西五台山是华北地区最高山峰,海拔3 061 m,气候条件复杂,生态环境类型特殊,是研究植物不同器官化学计量特征沿海拔变化的理想区域。本文以五台山不同海拔臭冷杉为研究对象,通过分析不同海拔臭冷杉叶、枝、根的C、N、P含量及其化学计量特征的变化规律,阐明海拔梯度臭冷杉不同器官的养分变化特征,为保护区臭冷杉的科学经营和管理保护提供依据。
1 研究地区与研究方法
1.1 研究区概况
五台山臭冷杉森林群落位于山西臭冷杉省级自然保护区(39°02′04″~39°13′01″N,113°20′55″~113°37′22″E)内,地处山西省繁峙县境内五台山北坡。保护区属暖温带湿润半湿润气候,年平均气温4℃左右,极端最高气温38℃,极端最低气温-30℃,全年降水量700 mm。土壤类型主要为山地森林棕壤土、山地淋溶褐土以及山地褐土。山西臭冷杉省级自然保护区是臭冷杉分布在我国的最南界,是臭冷杉在山西省分布的唯一区域。臭冷杉为山西省特别稀有的珍贵树种。
1.2 样品采集
2021年8月,在核心区内分布臭冷杉的地段设置3个海拔梯度,即2 050 m,2 150 m,2 250 m。在每一个海拔梯度,选取臭冷杉长势良好、干扰较少的地段设置样地,建立3个20 m × 20 m样地,进行每木调查。在每个样地,选取3株生长良好、干型通直的平均木作为样树。用高枝剪采集样树冠层中央朝南部位枝条,采集适量健康成熟叶片及枝条,带回实验室。同时采集每株样树朝南部位的根系样品,带回实验室。
1.3 测定项目与方法
将采集的植物样品叶、枝和根系105℃杀青,70℃烘干至恒重,粉碎,过筛,然后密封贮存,以备分析。有机碳含量采用重铬酸钾外加热法测定,全氮含量采用凯氏定氮法测定,全磷含量采用NaOH碱溶-钼锑抗比色法测定。
1.4 数据处理
采用Excel 2010、SPSS 20.0软件对数据进行统计分析。采用单因素(one-way ANOVA)法进行方差分析(α=0.05);对C、N、P含量及其比值进行Pearson相关性分析。图表中数据为平均值±标准差。
2 结果与分析
2.1 不同海拔臭冷杉叶、枝、根C、N、P含量
从不同器官看,各海拔梯度叶、枝、根C含量均值分别为 532.51、505.53、521.04 g·kg-1,表现为叶>根>枝,但差异不显著。其中,叶C含量和枝C含量随海拔增加而增加,根C含量随海拔增加呈先增加后减少的趋势。各海拔梯度叶、枝、根N含量均值分别为 17.30、11.49、7.22 g·kg-1,表现为叶>枝>根,且叶N含量显著高于枝N和根N。其中,叶N含量和枝N含量随海拔增加呈先增加后减少的趋势,根N含量随海拔增加而增加。各海拔梯度叶、枝、根P含量均值分别为1.63、1.77、1.04 g·kg-1,表现为枝>叶>根,且枝P含量和叶P含量显著高于根P(表1)。

表1 不同海拔臭冷杉叶、枝、根的C、N、P含量Tab. 1 The concentrations of C, N and P in leaves, branches and roots at three different altitudes of A. nephrolepis(单位:g·kg-1)
2.2 不同海拔臭冷杉叶、枝、根C、N、P化学计量特征
从不同器官看,各海拔梯度叶、枝、根C/N均值分别为30.89、45.40、74.44,表现为根>枝>叶,且差异显著。各海拔梯度叶、枝、根C/P均值分别为329.22、295.45、580.93,表现为根>叶>枝,且根 C/P显著高于叶C/P和枝C/P。各海拔梯度叶、枝、根N/P均值分别为10.65、6.60、7.66,表现为叶>根>枝,且叶N/P显著高于根N/P和枝N/P(表2)。
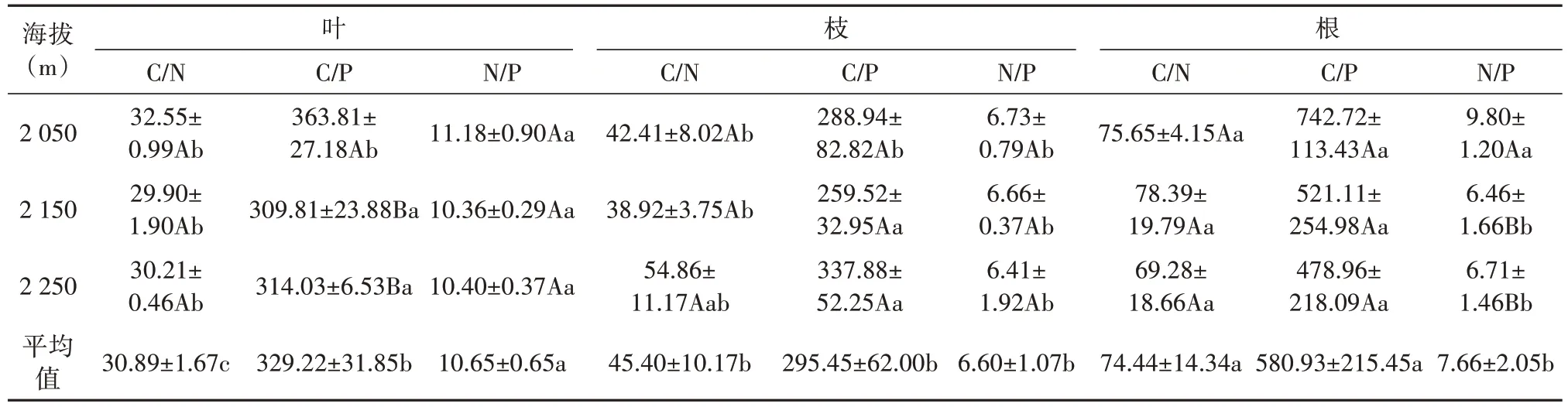
表2 不同海拔臭冷杉叶、枝、根的C/N、C/P、N/PTab. 2 The C/N, C/P and N/P in leaves, branches and roots at three different altitudes of A. nephrolepis
2.3 不同海拔臭冷杉叶、枝、根C、N、P含量及生态化学计量比的相关性分析
由表3可以看出,臭冷杉叶、枝、根C、N、P含量存在一定的相关性:臭冷杉叶N含量与叶C含量显著相关,叶P含量与叶C、N含量分别达到显著正相关和极显著正相关,枝C含量与叶C、N含量的相关性达到极显著水平,根C含量与枝N含量显著相关,根P含量与根N含量呈极显著正相关。臭冷杉各器官C/N、C/P与N/P之间关系也较为密切(表4):叶C/P与叶C/N、叶N/P呈极显著正相关,根C/P与叶C/P、根C/N存在显著正相关,根N/P与叶C/N、叶C/P、叶N/P以及根C/P表现出很强的相关性,并且与叶C/N、叶C/P、根C/P达到极显著正相关。
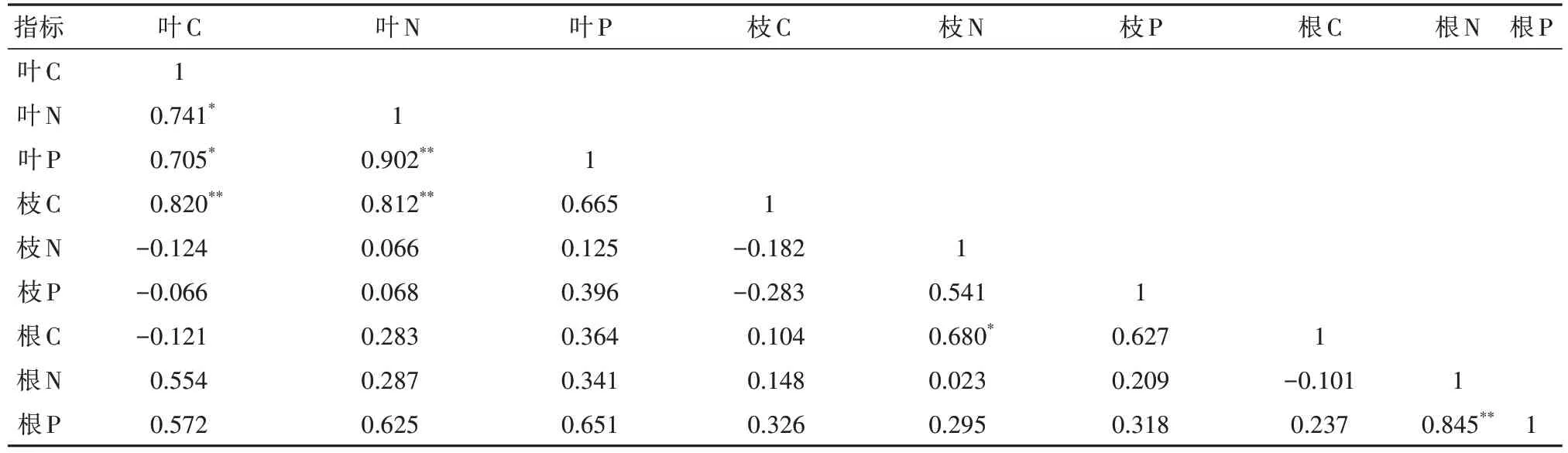
表3 臭冷杉叶、枝、根C、N、P含量的相关性Tab. 3 Correlation between C, N, P contents in leaves, branches, and roots of A. nephrolepis
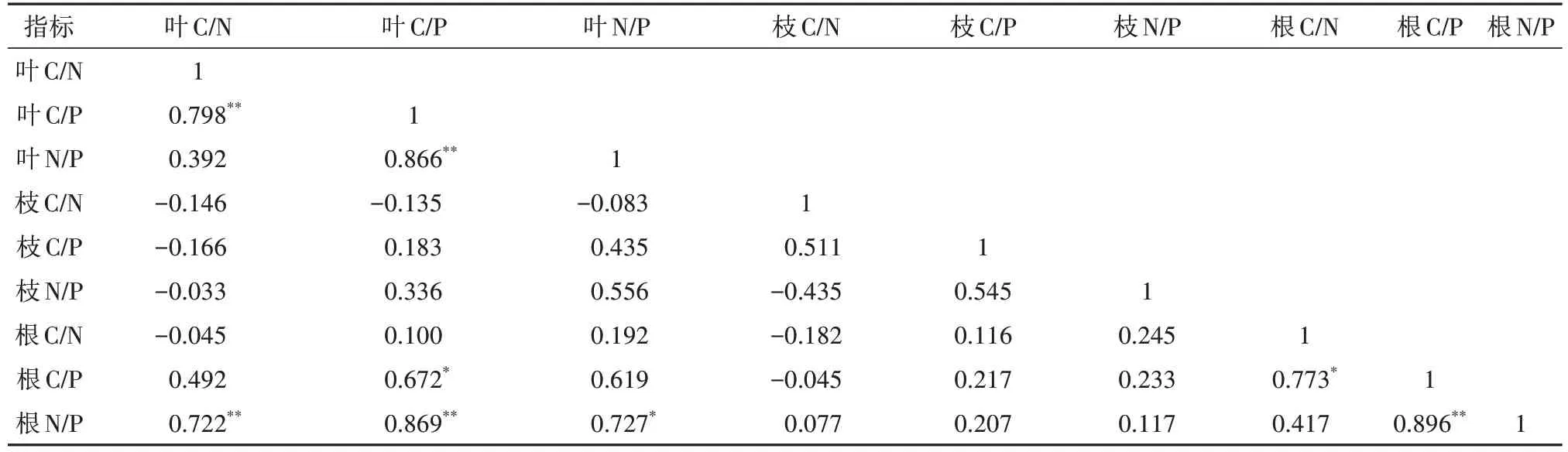
表4 臭冷杉叶、枝、根化学计量比的相关性Tab. 4 Correlation between stoichiometric ratios in leaves, branches, and roots of A. nephrolepis
3 讨论
3.1 臭冷杉叶片C、N、P含量及化学计量学总体特征
海拔的变化通常会导致光照、温度、降水等生态因子发生明显改变,形成特定生境,进而对植物叶片化学计量特征产生明显影响[18]。本研究中,臭冷杉叶片C含量平均值为532.51 g·kg-1,明显高于全球尺度的叶 C含量(461.60 g·kg-1)[3,19],表明臭冷杉叶片含有较多的有机物,固持碳的能力较强。这主要是由于五台山区太阳辐射强,昼夜温差较大,白天温度高,光合作用强,夜间温度低,抑制了呼吸速率,有利于有机质的积累。臭冷杉叶片N含量平均值为17.30 g·kg-1,高于我国针叶林叶N含量(13.13 g·kg-1)[20],但 低于 中 国 区 域 的 叶 N 含量(19.70 g·kg-1)[21-22]和 全 球 尺 度 的 叶 N 含 量(20.10 g·kg-1)[3,19]。臭冷杉叶片N含量较低,这可能与臭冷杉生长区域土壤环境中N素缺乏有关,表明臭冷杉生长具有较高的适应能力,以此应对N素长期供给不足,也说明N素是臭冷杉生态系统矿质营养的主要限制因素[23]。臭冷杉叶P含量平均值为1.63 g·kg-1,高于我国针叶林叶P含量(1.20 g·kg-1)[20],也 高 于 中 国 区 域(1.50 g·kg-1)[21-22]及 全 球 尺 度(1.40 g·kg-1)[3,19]的P含量。
植物叶片C/N、C/P能体现植物在吸收N、P时同化C的能力[24],故在一定程度上可以反映植物的生长速率以及植物的养分利用效率[25]。本研究中,不同海拔臭冷杉叶片C/N平均值为30.89,高于全球平均水平(22.50)[3],不同海拔臭冷杉叶片 C/P平均值为329.22,高于全球平均水平(300.90)[3]。许多研究将植物叶片N/P作为判断植物养分限制的指标[1],通常认为N/P>16表示植物生长受P限制,N/P<14则受N限制,N/P处于两者之间为植物生长受N和P共同限制[26]。本研究发现,不同海拔臭冷杉叶片N/P平均值为10.65,低于中国区域的叶N/P(18.00)和全球尺度的叶 N/P(13.80)[23],同时也低于植物生长受到土壤N元素限制的阈值(14),表明臭冷杉在不同海拔梯度均受到土壤N素的限制。因此,在保护区臭冷杉的抚育管理中,为了保证N素供应,需要合理施N肥,同时也可以在臭冷杉林中种植豆科固氮植物,以此来提高土壤的N素含量,改善土壤养分供给状况。
3.2 臭冷杉不同器官化学计量变化特征
分析植物不同器官的元素变化特征及分配策略有助于了解植物的生态适应机制[27]。本研究中,臭冷杉不同器官C、N、P含量变化基本一致,表现为叶>枝>根,这与叶作为植物获取光同化产物的器官,而枝和根作为养分吸收与传导的通道有关。本研究中,臭冷杉叶C含量高于枝C含量与根C含量,主要是因为叶作为主要的光合场所,在长期适应环境的过程中,臭冷杉将更多的C分配到叶中来获取更多的光能,增大光合产物的同化过程,抵抗外界的干扰,这与对不同林龄榧树(Torreya grandis)叶C含量高于枝C含量与根C含量的研究结果相似[28]。本研究表明,不同器官N含量表现为叶>枝>根,这与史军辉等[24]的研究结果一致。不同海拔臭冷杉枝P含量高于根P含量,与孙雪娇等[29]、陈美玲等[30]的研究结果一致。但对干旱荒漠区胡杨(Populus euphratica)枝、叶、根化学计量特征的研究表明,根P含量高于枝P含量[24]。这可能是因为臭冷杉侧根和须根发达,根系吸收营养,在供给自身的同时还需运输大量养分维持地上部分的生长[31-32]。
本研究中,臭冷杉不同器官的C/N平均值均高于中国东部南北样带优势种的(29.10)[33],同时也高于全球平均水平(22.50)[3],臭冷杉叶、根的C/P平均值均高于全球平均水平(300.90)[3]。不同器官C/N表现为根>枝>叶,C/P表现为根>叶>枝,根C/N显著高于枝C/N,同时枝C/N显著高于叶C/N,根C/P显著高于枝和叶C/P,整体上根的化学计量比大于枝和叶,说明相对于根,枝和叶的生长速率更快。这与马飞等[34]对9个种源中间锦鸡儿(Caragana liouana)的研究结果不同,这是因为中间锦鸡儿是抗寒、抗旱、耐瘠薄树种,主要生长在毛乌素沙地,根系具有较快的生长速率,有助于中间锦鸡儿从更深的土壤中吸收水分,以此应对旱季水分亏缺,而臭冷杉为耐荫树种,生长在冷湿环境中,因而具有不同的生长策略。臭冷杉不同器官N/P表现为叶>根>枝,叶N/P显著高于根和枝N/P,这可能与臭冷杉生长在冷湿环境,叶片P更容易被淋溶有关[35]。马飞等[34]的研究也发现类似结果。本研究结果对臭冷杉的经营管理具有科学的指导意义,为臭冷杉的养分吸收和利用提供科学依据。
猜你喜欢
山西林业科技(2022年4期)2022-03-12
散文诗(2021年22期)2022-01-12
数学物理学报(2021年6期)2021-12-21
应用数学(2020年2期)2020-06-24
环球时报(2019-05-23)2019-05-23
花火A(2019年1期)2019-03-22
数学年刊A辑(中文版)(2018年2期)2019-01-08
少儿美术(快乐历史地理)(2018年7期)2018-11-16
现代养生·上半月(2017年7期)2017-07-07
河北遥感(2014年4期)2014-07-10
