
国产柿属(Diospyros)的叶表皮形态研究及其分类学意义
2023-03-09 02:14邹璞龙开莲涂铁要李永泉李世晋
热带亚热带植物学报 2023年1期
邹璞, 龙开莲, 涂铁要, 李永泉, 李世晋**
国产柿属()的叶表皮形态研究及其分类学意义
邹璞1,2,3,4*, 龙开莲5*, 涂铁要1,2, 李永泉6, 李世晋1,2**
(1. 中国科学院华南植物园,中国科学院资源保护与可持续利用重点实验室,广州 510650; 2. 华南国家植物园,广州 510650; 3. 中国科学院核心植物园, 广州 510650; 4. 广东省应用植物学重点实验室,广州 510650; 5. 湖北省林科院石首杨树研究所,湖北 石首 434400; 6. 仲恺农业工程学院园艺园林学院,广州 510225)
为了解柿属()植物的叶表皮特征,在光学显微镜下观察了46种2变种植物的叶表皮微形态特征,并进行了主成分分析。结果表明,柿属植物的叶表皮微形态可分为2组:A组的表皮细胞为不规则形,垂周壁浅波状或深波状;B组的表皮细胞为多边形,垂周壁平直或弓形。气孔全部位于下表皮,以无规则型为主,但同时有辐射型、环列型和十字型气孔存在。大多数种具有表皮毛,表皮毛有非腺毛和头状腺毛的区别。柿属植物的叶表皮微形态特征类型多样,能够为部分类群的分类提供较好的形态学证据。
柿属;叶表皮形态;气孔;表皮毛;主成分分析
柿属()是柿树科(Ebenaceae)中最大的属,全世界约500种,广泛分布于热带地区,极少数物种分布于温带地区,其中亚洲约200种,南美洲100~130种,非洲大陆及马达加斯加各约100种[1–8]。我国约60种[9],其中特有种约44种,主要分布在我国东南、华南及西南地区。
自林奈于1753年建立柿属以来,不同的学者根据不同的性状提出了多个属下分类系统,然而这些分类系统大多是基于不同地区的柿属植物提出的,White[10–12]先后3次对非洲的柿属植物进行整理,将非洲的91种分为18组,Singh[13]记录了产于印度的66种,将其分为27组。这些分类系统在对中国产柿属植物的分类具有一定的局限性,加之国产柿属中特有种多达44种,占总种数的2/3, 至今仍然没有基于国产柿属的属下分类系统,仅有吴征镒等[14]整理的云南分布17种,其中包括11新种2新变种,属下的分类系统采用的是Hiern (1873)系统中的5组,分别为sect.、sect.、sect.、sect.和sect.,此外还有2种和,分别置于另外的2组,但是没有指出具体属于哪个组。分子系统学方面,Yonemori等[15]利用K和ITS两个片段探究14种柿(9泰国的种,4温带地区的种和1非洲的种)之间的关系,结果表明与柿()拥有共同的祖先,且油柿()与柿的亲缘关系较近。Duangiai等[1–2]的研究表明,(隶属于单属科Lissocarpaceae)应该包含在柿树科内,即形成了广义的柿树科,此外广义柿树科主要分为2个大支,分别对应2亚科Lissocar- poideae和Ebenoideae,而后Lissocarpoideae亚科又大致分为3支,对应3属(和)。虽然柿属的系统学位置已基本确定,但是其属内种间关系不清楚,分子系统的结果尚未得到形态学的很好支持。
对于国产柿属分类学方面的研究资料不多,最近Tang等[16]确定了我国现有65种柿属植物,其中45种为特有种,18种分布极为狭窄。最近童毅华等[17]发表了广东柿属一新组合,丹霞柿[(R. H. Miao & W. Q. Liu)Y. H. Tong & N. H. Xia]。袁录霞[18]基于K和L序列对大别山柿属植物DNA条形码及其适用性进行的研究,仅对部分种间关系进行了推断,认为乌柿()与岭南柿(),法国柿()与异色柿()可能同源。傅建敏[19]基于基因组浅层测序数据组装了柿、君迁子()、油柿、浙江柿()和金枣柿(‘Jinzaoshi’)等5种的叶绿体全基因组,系统发育分析表明柿与金枣柿、油柿和云南野毛柿()亲缘关系更近,而与浙江柿和君迁子亲缘关系较远。
迄今为止,尚未见对整个国产柿属植物进行系统的分类学研究,有许多具有潜在分类和系统学价值的性状值得挖掘。叶表皮的微形态解剖学研究是分类学和系统学的常用方法之一。Wilkinson[20]曾提出,在科及科以上水平,气孔器的结构与系统发育没有直接的相关性,但在某个限定的分类群中(如1科或1属),如果结合其他的演化性状,不同的气孔器类型间可能存在有系统学意义的联系。叶表皮的形态特征,如表皮细胞的式样、气孔的类型、保卫细胞的形状及角质层纹饰,通常在种间是恒定不变的,因此对于理解属间及属内的关系有着重要意义。20世纪中后期,Wilkinson[20]和Metcalfe等[21]曾报道柿树科植物叶表皮主要类型为无规则型和辐射型。近年来,Maiti等[22]报道了墨西哥2种柿属植物(和)叶表皮的气孔类型为无规则型,Ugbogu等[23]报道尼日尼亚30种柿属植物的叶表皮气孔类型有无规则型、多细胞型、环列型和十字型,表皮毛分为腺毛或非腺毛。Akinsulire等[24]研究了尼日尼亚西南部13种柿属植物的叶表皮形态和叶脉,认为叶表皮和脉络特征在柿属分类中很重要,并指出叶表皮气孔类型有无规则型、平列型、环列型和十字型。依泽等[25]对国产柿属5种1变种的叶片脉序、表皮形态、叶解剖结构、叶粉末等特征进行了比较观察,认为该属植物叶片的二级脉与三级脉具有鉴别意义; 发现该属植物叶表皮皆具有单细胞非腺毛,大多具有腺毛;个别种的粉末中可见石细胞,认为叶片特征可用于柿属药用植物营养期的鉴别。但目前对柿属植物叶表皮的研究还比较缺乏。本文详细研究了国产柿属44种和2亚种,以及2国外种的叶表皮气孔类型和表皮毛情况,以期为国产柿属植物的分类和系统学研究提供微形态学资料。
1 材料和方法
1.1 材料
本研究对46种2变种国产柿属()植物(含国外2种)进行了光镜观察,除少数材料为新鲜叶片外,其余均取自标本馆馆藏标本。本文分类单元参考, Vol. 15[9],凭证标本信息见表1。
1.2 方法
制片方法 采用过氧化氢-醋酸法[26]。经预实验后,将植物叶片近中脉两侧切成1 cm×1 cm的小块,浸泡在等量的30%过氧化氢-醋酸溶液中,在60 ℃烘箱内放置12~24 h (时间长短因叶质地不同而稍有不同,一般革质的叶片处理时间较长,必要时亦可延长处理时间,而纸质或膜质的叶面处理时间较短),待叶肉组织和表皮细胞分离后,将离析材料取出,用毛笔轻轻扫除叶肉细胞,漂洗干净,1%番红染色1~5 min,制作成临时装片,在光学显微镜(Olympus BX41)下观察和拍照。
气孔密度计算方法 在固定倍数下,用光学显微镜(Olympus BX41)对表皮的不同部位进行10次拍照,统计每张照片中气孔的数目,取平均值, 然后在相同的倍数下对标尺进行拍照,得出叶表皮的长(a)和宽(b),视野面积=×,最后用视野中气孔数除以即得气孔密度,以n/mm2表示。
本文气孔类型所用术语参考Dilcher[27]和Bara- nova[28]的描述,表皮毛参考Wilkinson[20]的描述。
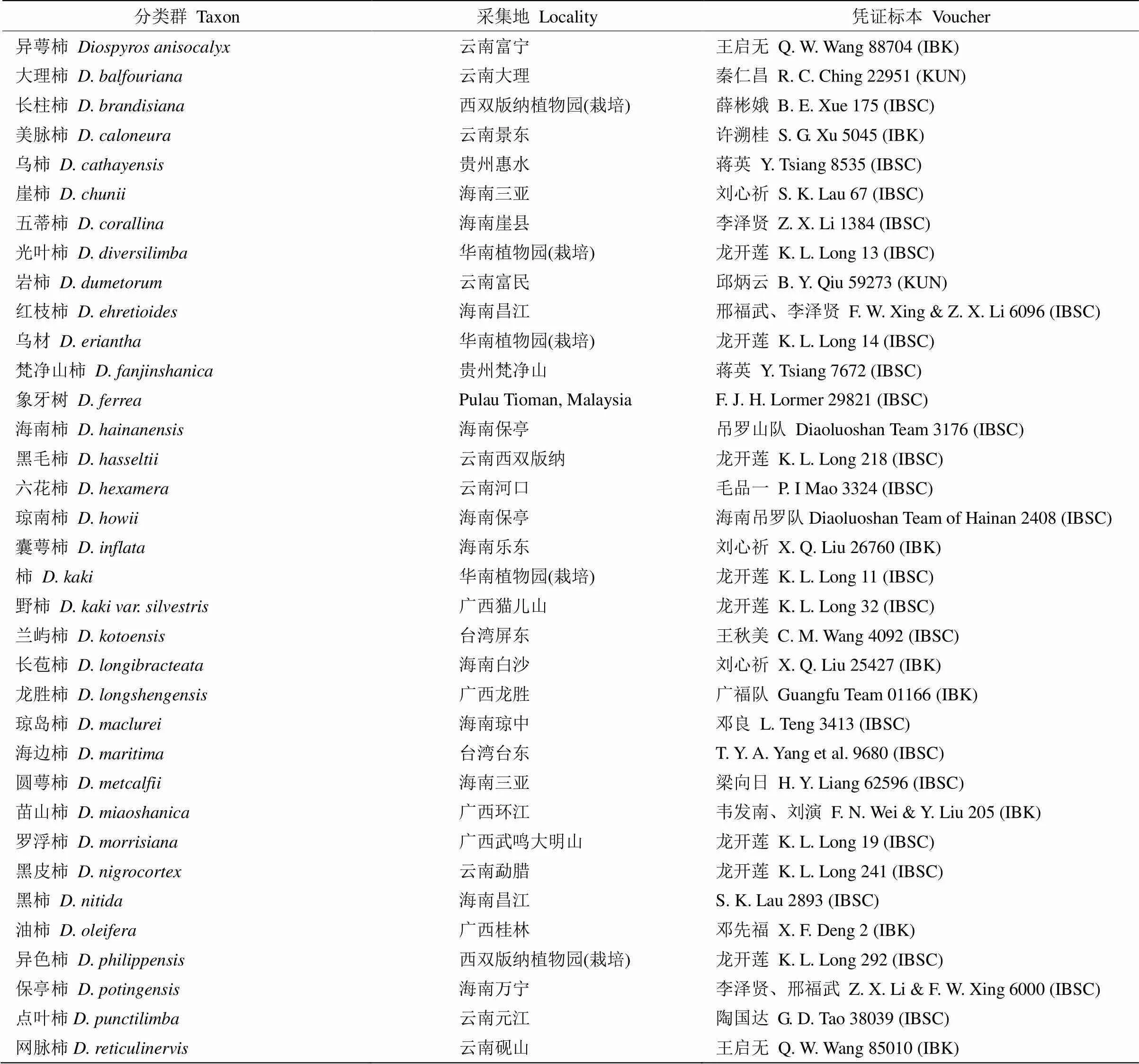
表1 国产柿属材料

续表(Continued)
1.3 数据分析
采用Rstudio软件分别对上、下表皮细胞的形状、垂周壁、有无加厚、有无表皮毛、表皮毛类型及其数目(有几种表皮毛),以及气孔器的类型、数目、密度、长、宽和长宽比等形态特征构成的矩阵进行主成分分析。
2 结果和分析
2.1 表皮细胞
根据观察,国产杮属植物的叶表皮细胞形状大致可以分为3种类型:(1) 上下表皮细胞均为多边形(常为5~6边形),大小不等,垂周壁平直或弓形,细胞壁又可分为2种亚型, 明显加厚[黑毛柿(),图2: A, B]和不加厚[海南柿(),图1: I, J)]。(2) 上下表皮细胞均为不规则形,垂周壁分为浅波形和波状,细胞壁亦可分为2种亚型, 加厚[五蒂柿(),图1: E, F)]与不加厚[长柱柿(), 图1: C, D)]。(3)上表皮细胞为多边形,垂周壁平直或弓形, 下表皮细胞则为不规则形,垂周壁浅波状,仅有大理柿(, 图1: A, B)、油柿(,图2: E, F)和保亭柿(, 图2: G, H)少数几种(表2)。
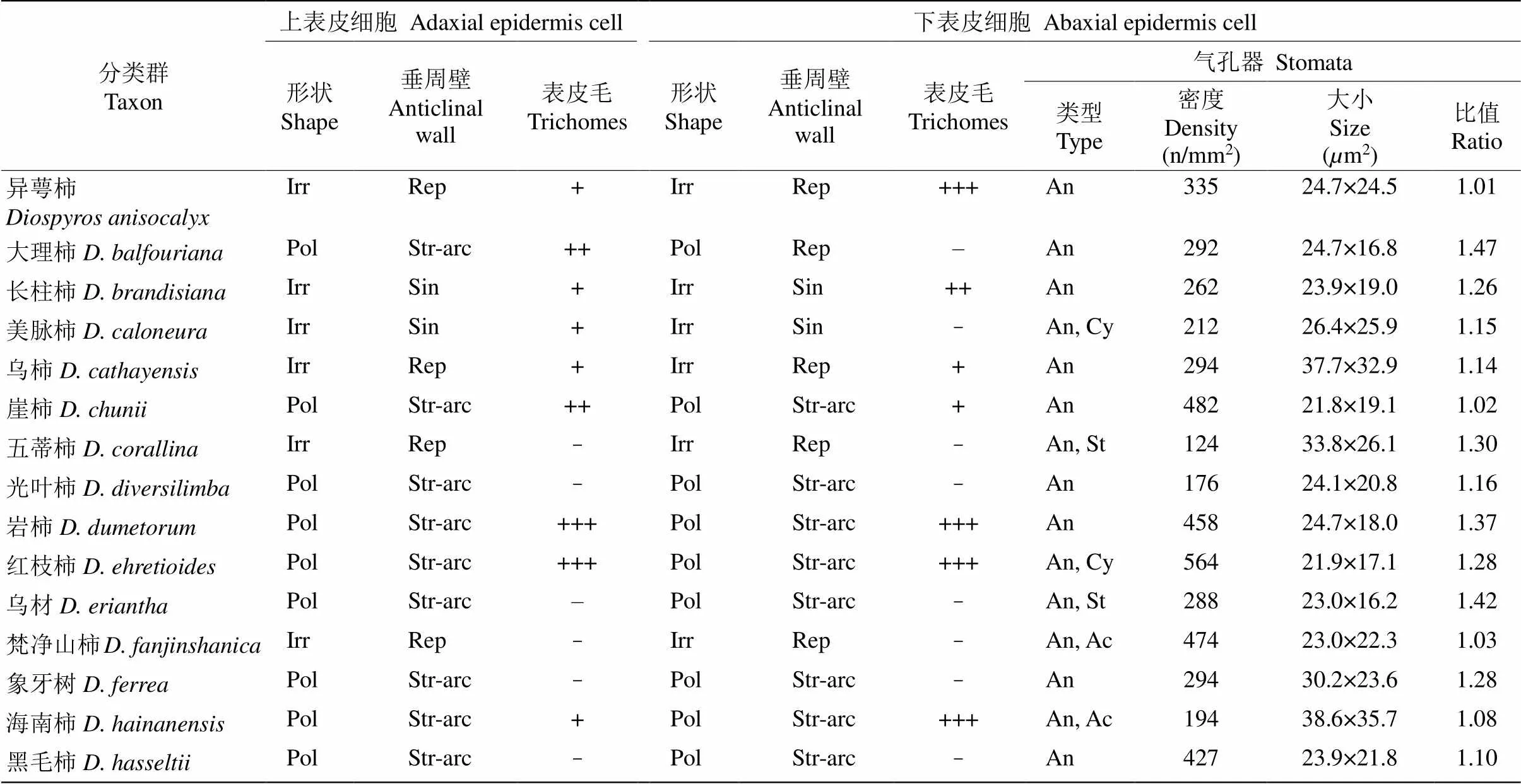
表2 光镜下国产柿属植物叶表皮细胞特征和气孔类型

续表(Continued)
Irr: 不规则型; Pol: 多边形; Str-arc: 平直-弓形; Rep: 浅波状; Sin: 深波状; An: 无规则型; Ac: 辐射型; Cy: 环列型; St: 十字型; –: 没有; +: 有; ++: 较多; +++: 极多。
Irr: Irregular; Pol: Polygonal; Str-arc: Straight to arched; Rep: Repand; Sin: Sinuous; An: Anomocytic; Ac: Actinocytic; Cy: Cyclocytic; St: Staurocytic; –: None; +: Have; ++: Many; +++: More.
2.2 气孔器
国产柿属植物叶片的气孔均分布在下表皮。气孔器主要为不规则型,同时也伴有其他类型, 如辐射型、环列型和十字型(表2)。不规则型的气孔周围常有4~6个细胞包围,其与正常细胞无异,如长柱柿(图1: D)。但是有些种气孔下陷,使得其气孔周围的轮廓不清晰,如光叶柿(, 图1: H)。辐射型的保卫细胞由呈辐射状延长的副卫细胞包围,如梵净山柿()、海南柿(,图1: J)和圆萼柿()。环列型的保卫细胞被1~2圈,每圈由4或4个以上的窄副卫细胞围绕,如美脉柿()、异色柿()、囊萼柿()和山榄叶柿(,图3: B)。十字型的保卫细胞被4个大小相似与之成直或斜十字形排列的副卫细胞所包围。如五蒂柿(图1: F)、乌材()、琼南柿()、过布柿()和岭南柿()。
国产柿属植物叶片的气孔形状大致分为扁圆形、近圆形或者近椭圆形,其气孔的长/宽比为0.77~1.71。气孔大小相差较大,最大的为海南柿(38.6m×35.7m)(图1: J),最小为黑皮柿(, 19.7m×17.1m)(图2: D)。平均气孔密度为319 n/mm2,最低的为五蒂柿(124 n/mm2, 图1: F),最高为毛柿(, 570 n/mm2, 图3: D)。

图1 柿属植物的叶表皮微形态。A, B: 大理柿; C, D: 长柱柿; E, F: 五蒂柿; G, H: 光叶柿; I, J: 海南柿。A, C, E, G, I: 上表皮; B, D, F, H, J: 下表皮。标尺=50 µm
2.3 表皮毛
46种2变种国产柿属叶片有无表皮毛的情况可分为4种(表2):(1) 上表皮和下表皮均无表皮毛(五蒂柿、光叶柿、乌材、梵净山柿、象牙树、黑毛柿、六花柿、长苞柿、琼岛柿、圆萼柿、黑皮柿、黑柿、网脉柿、过布柿和小果柿等15种);(2) 上表皮有表皮毛下表皮无(大理柿和美脉柿2种);(3) 下表皮有表皮毛而上表皮无(琼南柿、兰屿柿、异色柿、青茶柿和山榄叶柿等5种);(4) 上表皮和和下表皮均有表皮毛(异萼柿、长柱柿、乌柿、崖柿、岩柿、红枝柿、海南柿、囊萼柿、柿、龙胜柿、海边柿、苗山柿、罗浮柿、油柿、保亭柿、点叶柿、老鸦柿、石生柿、毛柿、信宜柿、岭南柿、单子柿、湘桂柿和云南柿等24种,野柿和无毛网脉柿2变种)。本研究没观察到有分枝的表皮毛,即表皮毛均为不分枝的单列毛,有的集中在叶脉处(如云南柿, 图3: G),有的分布于整个叶面[如异萼柿(,图3: I)]。其中12种和1变种[如异萼柿(图3:I)和野柿(var.)]有腺毛和非腺毛;另外12种和1变种仅有非腺毛[如云南柿(图3: G)和无毛网脉柿(s var.)]。非腺毛均为单细胞毛,细长,如云南柿(, 图3: G)。腺毛均为头状腺毛,其中异萼柿、红枝柿、柿、点叶柿()、乌柿()、油柿、信宜柿和岭南柿仅有1种头状腺毛(图3: G),但油柿为长柄腺毛,顶端为单个膨大细胞(图2: F)。老鸦柿()有2种头状腺毛,一种顶端为3~4个直径相同的单列细胞(图2: J),另一种顶端为2个较膨大的细胞(图3: J)。
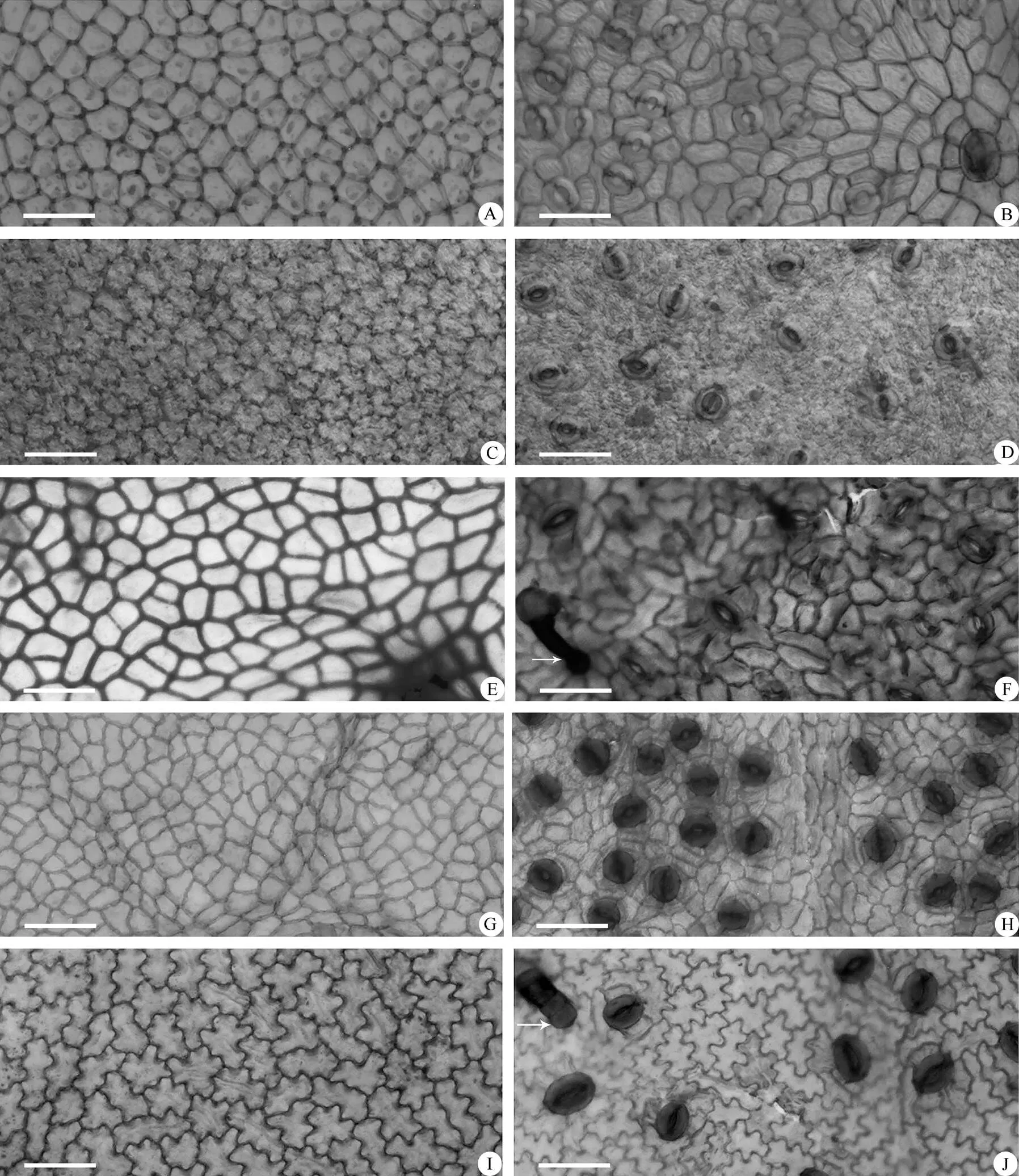
图2 柿属植物的叶表皮微形态。A, B: 黑毛柿; C, D: 黑皮柿; E, F: 油柿; G, H: 保亭柿; I, J: 老鸦柿。A, C, E, G, I: 上表皮; B, D, F, H, J: 下表皮。箭头示头状腺毛。标尺=50 µm
2.4 主成分分析
经主成分分析,前4个主成分的贡献率分别为31.3%、19.7%、13.2% 和8.8%,累计贡献率达73%,已经代表了原始性状的大部分信息,故选出前4个主成分为新选主成分,从原18个性状在新选4个主成分上的因子载荷量可以看出,上、下表皮细胞形状、上、下表皮细胞垂周壁、上、下表皮毛类型和数目、有无上表皮毛在前4个主成分上都有较高的载荷量。因此,根据前2个主成分做二维散点图,然而所有个体呈现交错分布,无法有效区分出柿属的不同种(图4),但是通过叶表皮细胞形态与垂周壁的式样,可以把48种柿属植物分为2大组:A组的叶表皮细胞无规则型,垂周壁为深波状或浅波状;B组的叶表皮细胞为多边形,垂周壁平直或弓形。单子柿(45)虽然位置略有偏差,但根据其叶表皮细胞为多边形,垂周壁平直或弓形应属于B组。
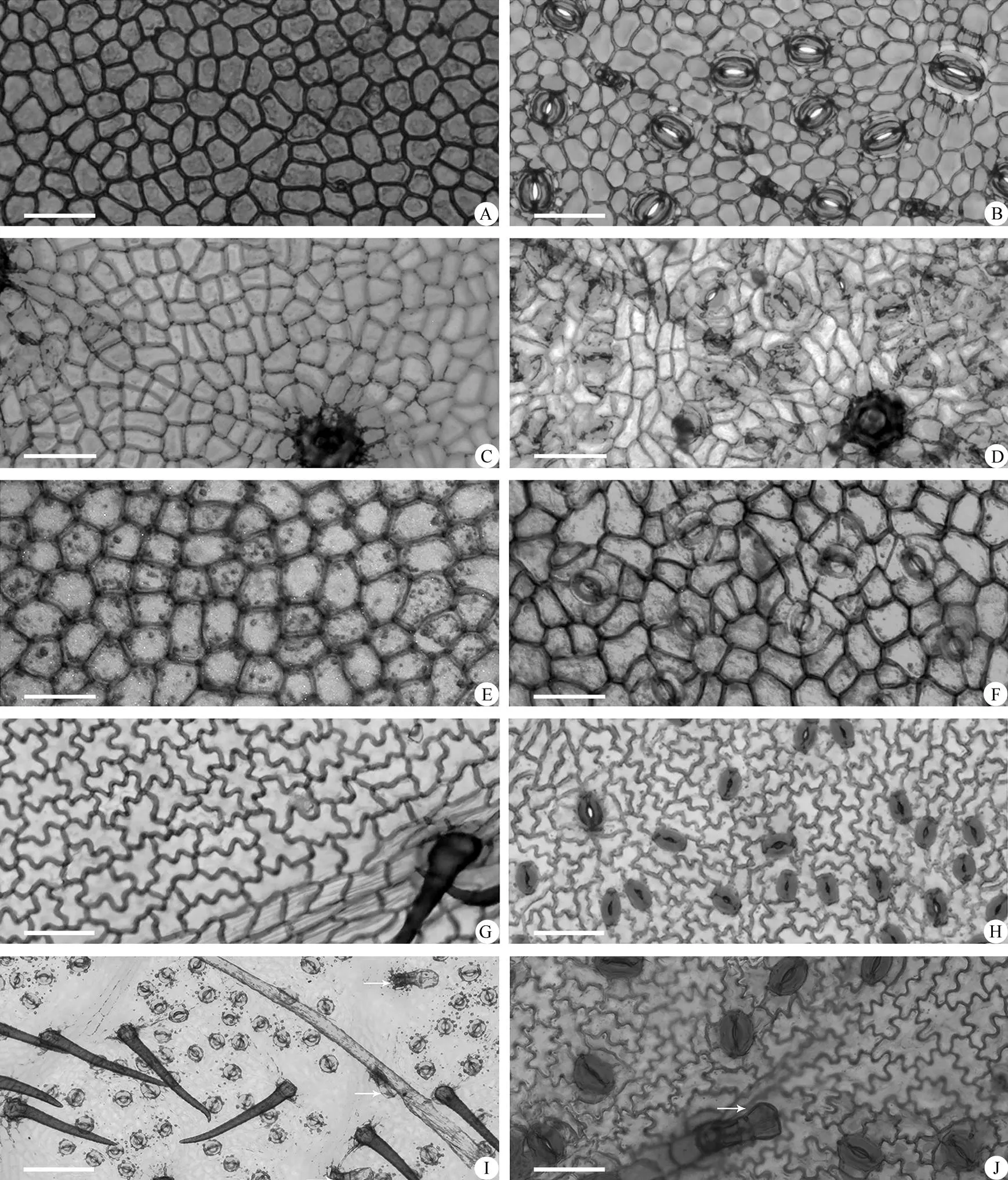
图3 柿属植物的叶表皮微形态。A, B: 山榄叶柿; C, D: 毛柿; E, F: 小果柿; G, H: 云南柿; I: 异萼柿; J: 老鸦柿。A, C, E, G: 上表皮; B, D, F, H, I, J: 下表皮。标尺=50 µm
3 结论和讨论
3.1 叶表皮特征及其系统学意义
本研究报道了国产柿属46种2变种的叶表皮微形态特征,首先是叶表皮细胞可以分为3种类型:上下表皮形状均为多边形,垂周壁平直或弓形;上下表皮细胞形状均为不规则形,根据垂周壁波状起伏的程度分为浅波形和波状;上表皮细胞为多边形,垂周壁平直或波状,下表皮细胞则为多边形或者不规则形,垂周壁垂直或波状。其次,仅在下表皮分布有气孔器,气孔器可分为4种类型:主要为无规则型,其余还有辐射型、环列型、十字型,通常与无规则型并存。再者,表皮毛的分布也可以分为3种情况,有的上表皮和下表皮均无毛;有的只有一面有毛,有的上下表皮都有毛。表皮毛又可以分为腺毛和非腺毛,腺毛又有单细胞和单列多细胞之分。
Duangjai等[2]利用6个叶绿体片段对99种广义柿树科(柿树属83种)植物进行了系统学研究,认为柿属有11个进化支(clade)。该研究仅包含了本研究中的7物种(长柱柿异色柿红枝柿象牙树柿海边柿和老鸦柿),7种植物分别位于4个进化支中,其中红枝柿和海边柿属关系近缘,聚于同进化支,并得到了叶表皮微形态特征的支持:如二者均属于B组具有多边形的叶表皮细胞,垂周壁平直或弓形,且细胞有加厚;均具有环列型和无规则型气孔器;上下表皮均被毛。虽然老鸦柿与红枝柿、海边柿同属于Clade Q,但它具无规则的叶表皮细胞,垂周壁深波状,仅有无规则型气孔器,上下表皮均被毛,属于B组叶表皮微形态与后2种(属于A组)有较大区别。长柱柿和异色柿同属于Clade K,但叶表皮微形态差异较大,前者属于A组具无规则的叶表皮细胞,垂周壁深波状,仅有无规则型气孔器,上下表皮均被毛;后者属于B组具有多边形的叶表皮细胞,垂周壁平直或弓形,具环列型和无规则型气孔器,仅下表皮被毛。柿和象牙树分别位于不同的进化支,没有其他种的叶表皮资料可以比较。
吴征镒等[14]曾经描述了17种2变种国产柿属植物,并分为7组。本研究的12种1变种植物包含在其中,但是叶表皮微形态特征不完全支持其划分。sect.中的乌柿、网脉柿和无毛网脉柿叶表皮微形态均属于A组,而琼南柿的叶表皮微形态属于B组;sect.中的单子柿和大理柿叶表皮微形态属于B组,而罗浮柿属于A组。
Wallnöfer[3]曾报道柿属植物和叶片上表皮有气孔器,但Ugbogu等[23]和Akinsulire等[24]在非洲尼日尼亚近50种柿属植物中没有观察到上表皮有气孔的现象,本研究中也没有观察到上表皮有气孔的情况,该性状还需要进一步的研究和确认。综合前人[23–24]的研究,目前认为全世界柿属植物的气孔器类型有无规则型、环列型、平列型、辐射型、多细胞型和十字型,但本研究中并没有观察到平列型和多细胞型气孔。Ugbogu等[23]报道尼日尼亚柿属植物叶片的表皮毛有3种类型:(1) 非腺毛,多细胞,无分枝;(2) 非腺毛,单细胞,无分枝;(3) 单列多细胞腺毛。本研究中的非腺毛与腺毛均为无分枝的单列毛,没有观察到多细胞的非腺毛,仅有单细胞的非腺毛,但观察到了多细胞的腺毛,均为头状腺毛,腺毛有4种类型: (1) 2个细胞组成,顶部膨大;(2) 2个细胞组成,具长柄, 顶端为单个膨大细胞;(3) 顶端为3~4个同样大小的细胞;(4) 顶端2个细胞。这几种腺毛在柿属中均为首次报道。
Ugbogu等[23]认为柿属叶表皮的气孔类型和每个区域的气孔数量在分类学上很重要,但是根据本研究主成分分析结果,我们认为国产柿属叶表皮最重要的分类性状应该是上表皮细胞形状、下表皮细胞形状、上表皮垂周壁式样、下表皮垂周壁式样、上表皮毛类型、上表皮毛类型的数目、下表皮毛类型、下表皮毛类型的数目和有无上表皮毛,而气孔器类型、气孔的密度与气孔长宽比值这些性状的可信度不高。通过主成分分析,本研究的48种柿属植物可分为2大类,A组中的岭南柿、乌柿、老鸦柿、异萼柿和美脉柿的叶表皮细胞为无规则型,垂周壁为深波状或浅波状,这些种的花冠常为坛状或者壶状,雌花单生,雄花常3朵成聚伞花序;而B组叶表皮细胞为多边形,垂周壁平直或弓形的琼南柿和小果柿的小枝均密被毛,雌雄花均单生,且雌雄花基本无差异。这些种的叶表皮微形态与其宏观形态一致。B组27种1变种柿属植物中至少有11种来自海南,叶表皮形态特征与产地的相关性值得进一步研究。
3.2 叶表皮特征可用于部分种的鉴定
在本研究中根据表皮毛特征可以鉴定油柿和老鸦柿:仅油柿的腺毛顶端膨大似气泡,而老鸦柿有2种不同的腺毛。
综上,目前国产柿属植物的叶表皮微形态特征类型多样,变化较明显,能够为部分类群提供较好的形态学证据。但是本研究明显的不足之处在于取样时仅仅选取了一份样品为代表,没有分不同居群采样,所以无法对叶表皮形态在不同居群是否具有稳定性进行评述,这一点将在今后的研究中改进。全世界的柿属植物有近500种,对其他种的叶表皮微形态的研究还非常有限,需要进一步深入的研究,以期获得更多有价值的形态学资料。
致谢 衷心感谢中国科学院华南植物园孔航辉副研究员和广西大学曾求标先生在论文修改中给予的帮助,同时感谢中国科学院华南植物园标本馆(IBSC)、中国科学院昆明植物所标本馆(KUN)、中国科学院广西植物研究所标本馆(IBK)和中国科学院西双版纳热带植物园标本馆(HITBC)提供标本。
[1] DUANGJAI S, SAMUEL R, MUNZINGER J, et al. A multi-locus plastid phylogenetic analysis of the pantropical genus(Ebenaceae), with an emphasis on the radiation and biogeographic origins of the New Caledonian endemic species [J]. Mol Phylogenet Evol, 2009, 52(3): 602–620. doi: 10.1016/j.ympev.2009.04.021
[2] DUANGJAI S, WALLNÖFER B, SAMUEL R, et al. Generic delimit- tation and relationships in Ebenaceae sensu lato: Evidence from six plastid DNA regions [J]. Am J Bot, 2006, 93(12): 1808–1827. doi: 10. 3732/ajb.93.12.1808
[3] WALLNÖFER B. The biology and systematics of Ebenaceae: A review [J]. Ann Naturhist Mus Wien, 2001 B, 103: 485–512.
[4] WALLNÖFER B. A revision of neotropical(Ebenaceae): Part 1 [J]. Ann Naturhist Mus Wien, 2007 B, 108: 207–247.
[5] WALLNÖFER B. A revision of neotropical(Ebenaceae): Part 2 [J]. Ann Naturhist Mus Wien, 2009 B, 110: 173–211.
[6] WALLNÖFER B. A revision of neotropical(Ebenaceae): Part 3 [J]. Ann Naturhist Mus Wien, 2010 B, 111: 101–133.
[7] WALLNÖFER B. A revision of neotropical(Ebenaceae): Part 4 [J]. Ann Naturhist Mus Wien, 2011 B, 112: 181–220.
[8] BAKHUIZEN VAN DEN BRINK, R. C. Revisio Enacearum Malayensium [J]. Bull Jard Bot Buitenzorg, serie 3, 1936–1955, 15(1–5): 1–515.
[9] LEE S G, GILBERT M G, WHITE F. Family Ebenaceae [M]// Wu Z Y, Raven P H. Flora of China, Vol. 15. Beijing: Science Press & St. Louis: Missouri Botanical Garden Press, 1996: 215–234.
[10] WHITE F. Notes on the Ebenaceae VIII: The African section of[J]. Bull Jard Bot Nat Belg, 1980, 50(3/4): 445–460.
[11] WHITE F. Ebenaceae [M]// Launert E. Flora Zambesiaca, Vol. 7(1). London: Flora Zambesiaca Managing Committee, 1983: 248–300.
[12] WHITE F. Twenty-two new and little-known species of(Ebenaceae) from New Caledonia with comments on section Maba [J]. Adansonia, 1992, 14(4): 179–222.
[13] SINGH V. Monograph on IndianL. (persimmon, ebony) Ebenaceae [M]. Kolkata: Botanical Survey of India, 2005: 1–323.
[14] WU Z Y, LI X W. Research Report on the Flora of Tropical and Subtropical Plants in Yunnan, Vol. 1 [M]. Beijing: Science Press, 1965: 9–23. [吴征镒,李锡文. 云南热带亚热带植物区系研究报告, 第1集[M]. 北京: 科学出版社,1965: 9–23.]
[15] YONEMORI K, HONSHO C, KANZAKI S, et al. Sequence analyses of the ITS regions and the matK gene for determining phylogenetic relationships of(persimmon) with other wild(Ebenaceae) species [J]. Tree Genet Genom, 2008, 4(2): 149–158. doi: 10.1016/j.scienta.2008.02.001
[16] TANG D L, ZHANG Q L, XU L Q, et al. Number of species and geographical distribution ofL. (Ebenaceae) in China [J]. Hort Plant J, 2019, 5 (2): 59–69. doi: 10.1016/j.hpj.2018.10.003.
[17] TONG Y H, XIA N H.(R. H. Miao & W. Q. Liu) Y. H. Tong & N. H. Xia (Ebenaceae), a new combination and a new synonym for a persimmon from Guangdong [J]. J Trop Subtrop Bot, 2019, 27(3): 346–348. [童毅华, 夏念和. 丹霞柿, 广东柿属(柿树科)一新组合及其一新异名[J]. 热带亚热带植物学报, 2019, 27(3): 346– 348. doi: 10.11926/jtsb.4053.]
[18] YUAN L X. Gentic diversity and phylogenetics of persimmon germ- plasm native to Dabieshan Region based on molecular marker and DNA barcoding [D]. Wuhan: Huazhong Agricultural University, 2012: 29– 85. [袁录霞. 基于分子标记和DNA条形码的大别山特产柿种质的遗传多样性和系统发育学[D]. 武汉: 华中农业大学, 2012: 29–85.]
[19] FU J M. Chloroplast genome sequences ofspp.: Phyloge- netic and genetic variation analyses [D]. Changsha: Central South University of Forestry and Technology, 2017: 11–65. [傅建敏. 柿属植物叶绿体测序及其系统发育和遗传变异研究[D]. 长沙: 中南林业科技大学, 2017: 11–65.]
[20] WILKINSON H P. The plant surface (mainly leaf) [M]// Metcalfe C R, Chalk L. Anatomy of the Dicotyledons, Vol. 1. 2nded. Oxford: Clarendon Press, 1979: 97–165.
[21] Metcalfe C R, Chalk L. Anatomy of the Dicotyledons, Vol. 2 [M]. Oxford: Clarendon Press, 1950: 880–886.
[22] MAITI R, RODRÍGUEZ H G, BALBOA P C R, et al. Leaf surface anatomy in some woody plants from northeastern Mexico [J]. Pak J Bot, 2016, 48(5): 1825–1831.
[23] UGBOGU O A, OLOWOKUDEJO J D, OGUNDIPE O T. Leaf epi- dermal morphology of(Ebenaceae) in Nigeria [J]. Phytol Balcan, 2016, 22(3): 377–385.
[24] AKINSULIRE O P, OLADIPO O T, ABDULRAHEEM O A, et al. Taxonomic significance of epidermal and venation characters in the genusL. (Ebenaceae) in Nigeria [J]. Braz J Biol Sci, 2018, 5(10): 499–514. doi:10.21472/bjbs.051026
[25] YI Z, QIAO J J, LU G Y, et al. Identification of six species of medicinalplants based on leaf macro-and micro-morphology [J]. China J Chin Mat Med, 2016, 41(21): 3942–3949. [依泽,乔娟娟,陆耕宇等. 柿属6 种药用植物营养期叶片辅助鉴别研究[J]. 中国中药杂志, 2016, 41(21): 3942–3949. doi: 10.4268/cjcmm20162110.]
[26] SUN T X, JIANG X S. H2O2-CH2COOH maceration: A simple and perfect method for leaf epidermis [J]. Guihaia, 2009, 29(1): 44–47. [孙同兴, 江幸山. 简便有效的叶表皮离析方法——过氧化氢-醋酸法 [J]. 广西植物, 2009, 29(1): 44–47.]
[27] DILCHER D L. Approaches to the identification of angiosperm leaf remains [J]. Bot Rev, 1974, 40(1): 1–157.
[28] BARANOVA M A. Historical development of the present classification of morphological types of stomates [J]. Bot Rev, 1987, 53(1): 62–63.
Leaf Epidermal Morphology offrom China and Its Taxonomic Significance
ZOU Pu1,2,3,4*, LONG Kailian5*, TU Tieyao1,2, LI Yongquan6, LI Shijin1,2**
(1. Key Laboratory of Plant Resources Conservation and Utilization, South China Botanical Garden, Chinese Academy of Sciences, Guangzhou 510650, China; 2. South China National Botanical Garden, Guangzhou 510650, China; 3. Core Botanical Gardens, Chinese Academy of Sciences,Guangzhou 510650,China; 4. Guangdong Provincial Key Laboratory of Applied Botany, Guangzhou 510650,China; 5. Shishou Research Institute of Poplar for Hubei Academy of Forestry,Shishou 434400, Hubei, China; 6. College of Horticulture and Landscape Architecture, Zhongkai University of Agriculture and Engineering, Guangzhou 510225, China)
In order to understand the leaf epidermal morphology of(Ebenaceae), the leaf epidermal characters of 46 species and 2 varieties were observed under light microscopes, and then the major characters were analyzed by principal component analysis. The results showed that the micro-morphology of the leaf incould be divided into two groups, Group A and Gruop B. The epidermal cells were irregular with repand or sinuous anticlinal walls in Group A, and which were polygonal with straight or arched anticlinal walls in Group B. The leaves of the genus were hypostomatic. The stomata were mainly anomocytic and sometimes actinocytic, cyclocytic or staurocytic. The trichomes were found in most species, which were either simple non-glandular or capitate glandular. Therefore, the epidermal characters would provide valuable morphological evidences for classification in some taxa of the genus.
; Leaf epidermis morphology; Stomata; Trichomes; Principal component analysis
10.11926/jtsb.4564
2021-11-12
2022-05-19
国家自然科学基金项目(31670193, 31070176)资助
This work was supported by the National Natural Science Foundation of China (Grant No. 31670193, 31070176).
邹璞,博士,研究方向为植物系统与进化。E-mail:zoupu@scbg.ac.cn
* 共同第一作者
E-mail:lisj@scbg.ac.cn
猜你喜欢
干旱地区农业研究(2022年1期)2022-01-28
建材发展导向(2021年14期)2021-08-23
装备制造技术(2019年12期)2019-12-25
祝您健康·文摘版(2019年4期)2019-06-11
科技创新与品牌(2019年12期)2019-02-06
中国烟草学报(2018年2期)2018-05-09
中国中西医结合皮肤性病学杂志(2016年4期)2016-07-18
中国房地产业(2016年9期)2016-03-01
中国铸造装备与技术(2015年5期)2015-12-10
湖南农业科学(2015年3期)2015-10-09
