
甘蓝型冬油菜气孔特性与抗寒性的关系
2023-03-09 08:40:18范军强武军艳刘丽君马骊杨刚蒲媛媛李学才孙万仓
中国农业科学 2023年4期
范军强,武军艳,刘丽君,马骊,杨刚,蒲媛媛,李学才,孙万仓✉
1甘肃农业大学农学院,兰州 730070;2省部共建干旱生境作物学国家重点实验室,兰州 730070
0 引言
【研究意义】气孔是植物适应逆境的重要叶片结构。气孔开闭直接影响植物体内部的CO2及水分含量,进而影响光合作用及蒸腾作用的速率,逆境下的气孔关闭导致植物光合作用下降,同化产物供应减少,因此,气孔调节在碳同化和有关水分的生理活动中具有重要作用,在植物用水效率及碳和水循环方面具有重要意义[1],同时,在植物应对逆境胁迫中也发挥重要作用[2]。了解气孔的响应机制与植物抗寒性的变化规律,对解析植物在低温逆境下的适应性有重要的意义。【前人研究进展】气孔随着环境温度的变化而发生运动[3]。低温环境胁迫使气孔趋于关闭,直接引起气孔导度的变化[4]。植物通过其体内大量的生理代谢抵抗非生物胁迫引起的内环境动态变化。低温不但会引起气孔的异常开闭[5],同时还可以提高ABA的含量[6],通过增加气孔中二氧化碳的浓度,间接降低气孔开度,影响光合作用[7]。夜间低温条件下,番茄叶片气孔导度和气孔面积显著降低[8],对开花期水稻孕穗进行低温处理,低温胁迫越强,气孔导度的值就越小[9]。目前为止,发现各种生物和非生物胁迫因素可能会迅速改变气孔开度,而对这些刺激的反应取决于保卫细胞内复杂的信号传输网络[10]。近年来,人们发现并鉴定了气孔调控网络的关键信号节点[11]。这些分子在特定的生境中,在亚细胞定位和迁移、蛋白质翻译后修饰、细胞内氧化还原稳态变化、离子通道开闭和基因转录调控等环境因子的持续波动下,对气孔进行精确调控[12]。据报道,冬性油菜品种的净同化率和 CO2交换率高于春性品种[13],这是气孔对低温的反应而非适应[14],表明气孔开度的调节对抗寒性有重要作用。有学者补充烟草叶片在保卫细胞增加外源钙吸收基础上表现出冷诱导气孔关闭,此反应不会发生在冷敏感的品种中[7]。【本研究切入点】北方地区甘蓝型冬油菜越冬前地上部与地下部会逐渐停止生长[15],叶片逐渐干枯死亡[16]。此时,叶片中发生复杂的代谢活动,从而适应冬季低温环境。气孔作为光合作用、呼吸作用、蒸腾作用的门户直接影响越冬前的生物量积累[17],间接影响冬油菜对越冬时环境的适应。2000年起,甘肃农业大学等对北方甘蓝型冬油进行了抗寒性品种改良研究,但是,改良品种在低温胁迫后的气孔运动是否发生变化,尚不明确。【拟解决的关键问题】本研究以抗寒性不同的 14个甘蓝型冬油菜品种为材料,对低温条件下不同抗寒性材料气孔运动特性与抗寒性进行相关性分析,探究甘蓝型冬油菜抗寒性与气孔运动特征存在的关系,为解析强冬性甘蓝型冬油菜抗寒机理提供新依据。
1 材料与方法
1.1 试验材料
选用 14个抗寒性不同的甘蓝型冬油菜品种为研究材料,其中10个来源于甘肃农业大学,4个来自天水市农业科学研究所。各品种基本信息见电子附表1。
1.2 试验设计
2021年8月21日将供试材料种子置于培养皿中萌发,于2021年8月25日统一移栽至直径为25 cm、高度为12 cm的树脂花盆中。每盆装入4 kg已加水搅拌均匀的育苗基质,每盆种3穴,每穴1苗,放置于甘肃农业大学百草园生长。六叶期取样处理及观测。
处理:试验共设3个处理,分别为24℃处理12 h、0℃处理 1 h和 0℃处理 12 h(全部盆栽均需先置于24℃培养12 h),处理过程中光照及水分条件一致。每个处理设置3个生物学重复。
采样:处理结束后快速采样。第2片叶用于气孔观察。每株展开的第3片叶用于生理指标的测定。第4片叶用于叶片相对含水量的测定。
1.3 测定项目及方法
1.3.1 半致死温度(LT50)测定 试验材料于 24℃培养12 h后,用打孔器将展开的第3片叶上打3个直径为5 mm的叶圆片(注意避开叶脉),装入10 mL离心管中,并加入0.2 mL去离子水,对照只加入去离子水,分别置于0℃、-4℃、-8℃、-12℃和-16℃条件下处理30 min,每个温度设置3个重复。取出后再加入10 mL去离子水,室温震荡1 h,用电导仪测处理样品的电导率C1和对照的电导率C0。置于120℃烘箱20 min,冷却至室温后,测电导率C2,相对电导率(relative electric conductivity,REC)=(C1-C0)/(C2-C0)×100%。
1.3.2 生理指标测定 使用烘干法测定叶片相对含水量;氮蓝四唑法(NBT)测定超氧化物歧化酶(superoxide dismutase,SOD);使用愈创木酚法测定过氧化物酶(peroxidase,POD);采用紫外吸收法测定过氧化氢酶(catalase,CAT);采用硫代巴比妥酸法(TBA)测定丙二醛(malondialdehyde,MDA)的含量;采用磺基水杨酸浸提-酸性茚三酮显色法测量游离脯氨酸(proline,Pro)含量[18]。生理指标的测定设置3个生物学重复,不设置技术重复。
1.3.3 气孔制片及观察测量 气孔制片:采用改良后的指甲油涂抹撕取法[19]。选用不含色素的透明指甲油,在植株展开的第2片叶片下表皮叶脉左侧上下居中的固定位置均匀涂抹一个薄层,大小约为1 cm×1 cm的矩形。晾干10 min左右(时间长短因环境条件不同),涂抹痕迹边缘会出现离口。剪下长宽适宜的透明胶带,胶面平贴于涂抹部位,轻轻撕下后贴于载玻片上,制片完成。
制片观察:使用正倒置一体显微镜(型号为Echo Revolve FL)拍照观察,放大倍数为 40×10。使用ImageJ软件测量,测量指标包括气孔孔隙长(stomatal pore length)、气孔孔隙宽(stomatal pore width)、气孔孔隙面积(area of stomatal pore)、气孔孔隙周长(circumference of stomatal pore)、气孔器长(stomatal apparatus length)、气孔器宽(stomatal apparatus width)、气孔器面积(area of stomatal apparatus)、气孔器周长(circumference of stomatal apparatus)、保卫细胞宽(guard cell width)、视野内气孔数(number of pores in visual field)、气孔开度(stomatal aperture)、气孔关闭率(stomatal closure rate),具体意义见图1。其中,气孔开度=气孔孔隙长×气孔孔隙宽,视野内气孔数指在400倍放大下圆形视野内的气孔个数(圆形视野面积:0.2409 mm2),气孔关闭率=视野中关闭气孔的数量/视野中全部气孔数。每张制片测定3个视野,3个视野内的全部气孔均需测量全部12个指标。
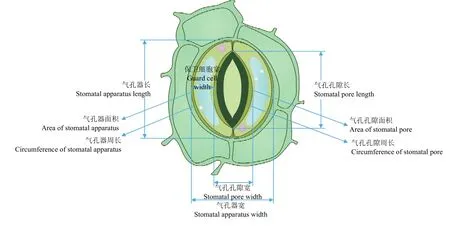
图1 气孔指标Fig.1 Stomatal index
1.4 数据处理及分析方法
1.4.1 Logistic回归方程分析 对测定的相对电导率进行Logistic回归分析,回归模型为:
其中,y为相对电导率,t代表处理温度,k为细胞伤害率的饱和容量,a、b为方程参数。因本试验中细胞伤害率消除了干扰,故k值为100。为确定a、b值,将Logistic方程转化为:
则方程转化为y′与t之间的线性方程,即:y′=ln(a)-by
1.4.2 综合评价数据分析 参照蒲媛媛等[20]方法计算单项指标耐寒系数,耐寒系数(K)=(低温测定值/常温测定值)。多重比较采用Tukey b的方法进行相同处理温度下品种间对比。
1.4.2.1 数据标准化 运用极差法对各指标耐寒系数进行标准化处理:
式中,xij表示i品种第j个指标的K值;xmin表示第j个指标K值的最小值;xmax表示第j个指标K值的最大值。指标与抗寒性成正相关用标准化公式(1)计算,指标与抗寒性呈负相关用公式(2)计算。
1.4.2.2 隶属函数综合评价 采用标准差系数法计算权重。标准差系数计算公式为(3),权重系数计算公式为(4)。
式中,n表示i个品种,m表示SOD、POD、CAT、MDA、Pro、叶片相对含水量j个指标。
综合评价值公式如下:
1.4.2.3 主成分分析综合评价 建立变量之间的相关系数矩阵:相关系数矩阵R=(rij)m×m
式中,rii=1,rij=rji,rij是第i个指标与第j个指标的相关系数。
计算相关系数矩阵R的特征值和特征向量:计算相关系数矩阵R的特征值λ1≥λ2≥…≥λm≥0,及对应特征向量u1,u2,……,um,其中uj=(u1j,u2j,…,unj)T,由特征向量m个新的指标变量。
式中,y1是第一主成分,y2是第二主成分,……,ym是第m主成分。
写出主成分并计算综合得分:计算特征值λj(j=1,2,…,m)的信息贡献率。
贡献率,当αp接近于 1(αp=0.85,0.90,0.95)时,则选择前p个指标变量y1,y2,……,yp作为p个主成分,代替原来m个指标变量,从而可以对p个主成分进行综合分析。接着计算综合得分,其中,bj为第j个主成分的信息贡献率。
2 结果
2.1 不同抗寒性甘蓝型冬油菜半致死温度(LT50)
相对电导率可表示细胞膜透性,离体叶片在一定温度范围内的渗透情况具有反映细胞膜受伤程度的作用。当处理低温低于半致死温度时,植物将受到不可逆的伤害,相对电导率快速升高。低温半致死温度高低是反映植物抗寒性强弱的重要指标。强抗寒性品种相对电导率升高速率较弱抗寒性品种慢,其中,gau系列相对电导率在-12℃达到较高水平,抗寒性相对较弱的天油系列在-8℃达到较高水平。根据LT50鉴定14个品种的抗寒性为gau-1(-8.06)>gau-24(-7.83)>gau-30(-7.58)>gau-39(-7.44)>ts309(-7.28)>ts312(-7.08)>nts158(-6.81)>npz269(-6.62)>天油 14 号(-5.98)>16-2444(-5.4)>17-2251(-5.13)>天油2266(-4.8)>天油2238(-4.6)>天油2288(-4.38)。回归分析后的拟合度处于 0.88—0.97(表1),可用于进一步分析。
2.2 不同抗寒性甘蓝型冬油菜低温处理下的生理响应特征
2.2.1 叶片中抗氧化酶活性变化 在低温处理后的叶片中,3种抗氧化酶(SOD、POD和CAT)的活性均呈上升趋势,但增幅不同。常温下,3种酶的活性处于最高值的品种分别是天油 2238(SOD酶活性为440.86 U·g-1FW)、npz269(POD 酶活性为 40.11 U·g-1·min-1)和天油 2238(CAT 酶活性为 32.60 U·g-1·min-1),均是半致死温度较高的材料,低温处理前后,品种间增幅差异显著(图2-A—C),且规律明显。增幅较大的品种有 gau1(SOD 40.26%、POD 165.61%、CAT 66.54%)和gau24(SOD 30.95%、POD 146.11%、CAT 60.83%)。常温条件下,天油 2238的3种酶活性均表现出较高水平,低温后增幅较小。


图2 甘蓝型冬油菜低温下的生理响应Fig.2 Physiological Responses of winter Brassica napus under low temperature
2.2.2 叶片渗透调节物质含量的变化 渗透调节物质的积累也是植物调节抗逆性的方式。试验结果(图2-D—E)显示,低温下丙二醛和游离脯氨酸含量均有所增加,但增幅变化趋势明显相反。常温下游离脯氨酸含量最高的品种为天油2238(369.65 mg·g-1),最低为 gau-24(141.12 mg·g-1),低温处理后增幅最为明显的是gau-1(149.66%)。常温条件下,天油2238、天油 2266、天油 2288的丙二醛含量(2.10、1.69和1.24 μmol·g-1)较低,低温处理后的变化量较为突出,较对照分别增加160.76%、145.64%和194.15%。常温条件下,gau-24、gau-39的丙二醛含量(3.59和3.07 μmol·g-1)较高,低温处理后增幅较小,分别为62.27%和78.72%。
2.2.3 叶片相对含水量的变化 叶片相对含水量在低温处理后明显降低,且反应灵敏。常温下叶片相对含水量品种间差异不显著,处于 82.52%—93.59%。低温处理后变化明显,天油2238、天油2266和天油2288下降最为明显,降至61.41%、66.25%和61.47%。
2.3 参试材料抗寒性评价
2.3.1 隶属函数法综合评价 将依据耐寒系数公式计算得到的数据列于表2,除LRWC以外,其余指标耐寒系数均大于 1。14个品种各指标耐寒系数均值POD及MDA较大,表明2个指标相对于对照增幅更大。运用极差法对各指标耐寒系数进行标准化,得到的无量纲数据列于表3,最大值1多分布于gau-1,最小值0全部分布于天油2288。对每个指标赋予利用标准差系数法算得的权重,求得各品种的综合评价值(D),该数据越大表明抗寒性越强。D值最大的品种为gau-1(0.990),最小的为天油2288(0.000)。据此获得的抗寒性为gau-1(0.990)>gau-24(0.876)>gau-30(0.693)>gau-39(0.644)>ts309(0.534)>ts312(0.463)>nts158(0.439)>npz269(0.388)>天油 14号(0.352)>16-2444(0.307)>17-2251(0.282)>天油2266(0.236)>天油2238(0.126)>天油2288(0.000)。
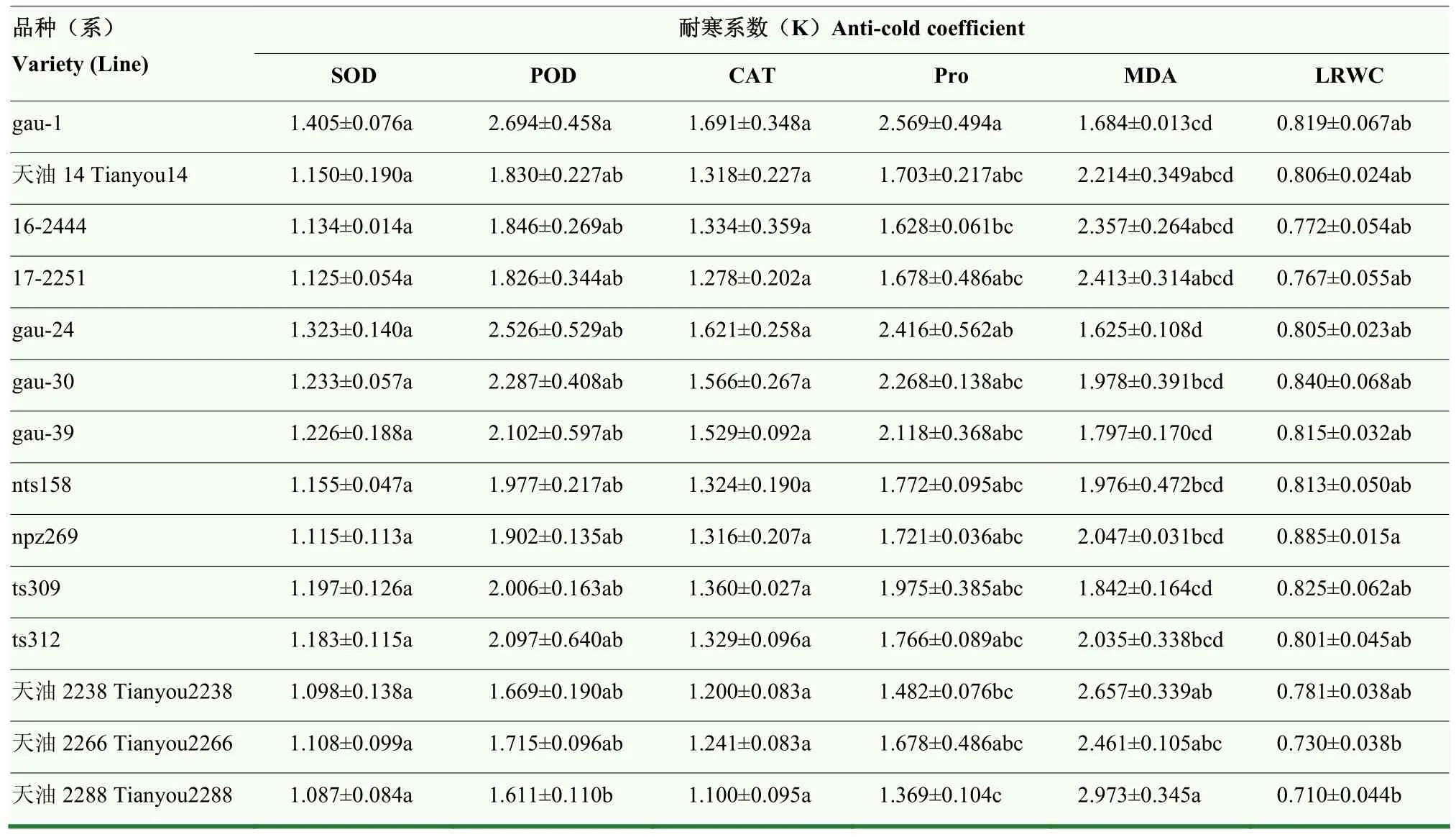
表2 主要生理指标的耐寒系数评价Table 2 Coefficients of cold tolerance evaluated of main physiological traits

表3 隶属函数值和综合评价值结果Table 3 Subordinator functional components and the integrated evaluation index for winter Brassica napus
2.3.2 半致死温度与相关指标间相关性分析 对半致死温度与综合评价值(D)、SOD、MDA、POD、CAT、Pro、LRWC分别做相关性分析(表4)。结果显示,半致死温度与综合评价值(D)、SOD、POD、CAT、Pro之间呈现极显著负相关性(P<0.01),相关系数处于-0.959—-0.831。与MDA间呈极显著正相关性,相关系数为0.959。

表4 半致死温度与生理指标的相关性Table 4 Correlation analysis of semi-lethal temperature and physiologic index
2.4 不同抗寒性甘蓝型冬油菜气孔运动特性
各指标的分析样本数量列于表 5。各处理品种间气孔孔隙长差异显著(图 3-A),连续的低温处理,使大部分品种气孔孔隙长在1 h时出现下降,处理12 h后,gau-1、gau-24、gau-30、gau-39与对照持平,天油2238、天油2266天油2288较对照明显下降。气孔孔隙宽低温处理后下降幅度大,天油2266和天油2288下降明显,天油2266 12 h处理后自7.34 μm降至5.84 μm,天油2288由9.14 μm降至6.68 μm(图3-B)。处理12 h后气孔孔隙面积天油2266自144.35 μm2降至 107.56 μm2,天油2288自 211.17 μm2降至142.37 μm2(图3-C),2个品种低温处理后降低明显。气孔孔隙周长(图 3-D)与气孔器长(图 3-E)各品种 3个处理间差异不显著。天油2288气孔器宽度(图3-F)低温处理12 h后减小5.85 μm,降幅较大。天油2288气孔器面积在低温处理后与对照相比下降了 28.53%(图 3-G)。气孔器周长(图 3-H)变化不明显。保卫细胞宽度的变化(图3-I),gau-1、天油14、ts312品种低温处理与对照持平,12 h处理后变化范围为0.01—0.05 μm。视野内气孔数除16-2444和npz269以外,其余品种低温处理后均呈现增加趋势(图3-J)。
气孔开度是综合气孔孔隙长和气孔孔隙宽2个指标的综合指标,变化幅度明显。14个品种总体变化范围为251.15—101.29 μm2。14个品种的平均气孔关闭率(图3-L)为 31.01%(24℃)、40.07%(0℃ 1 h)和 35.99%(0℃ 12 h),0℃低温处理后有明显上升趋势。
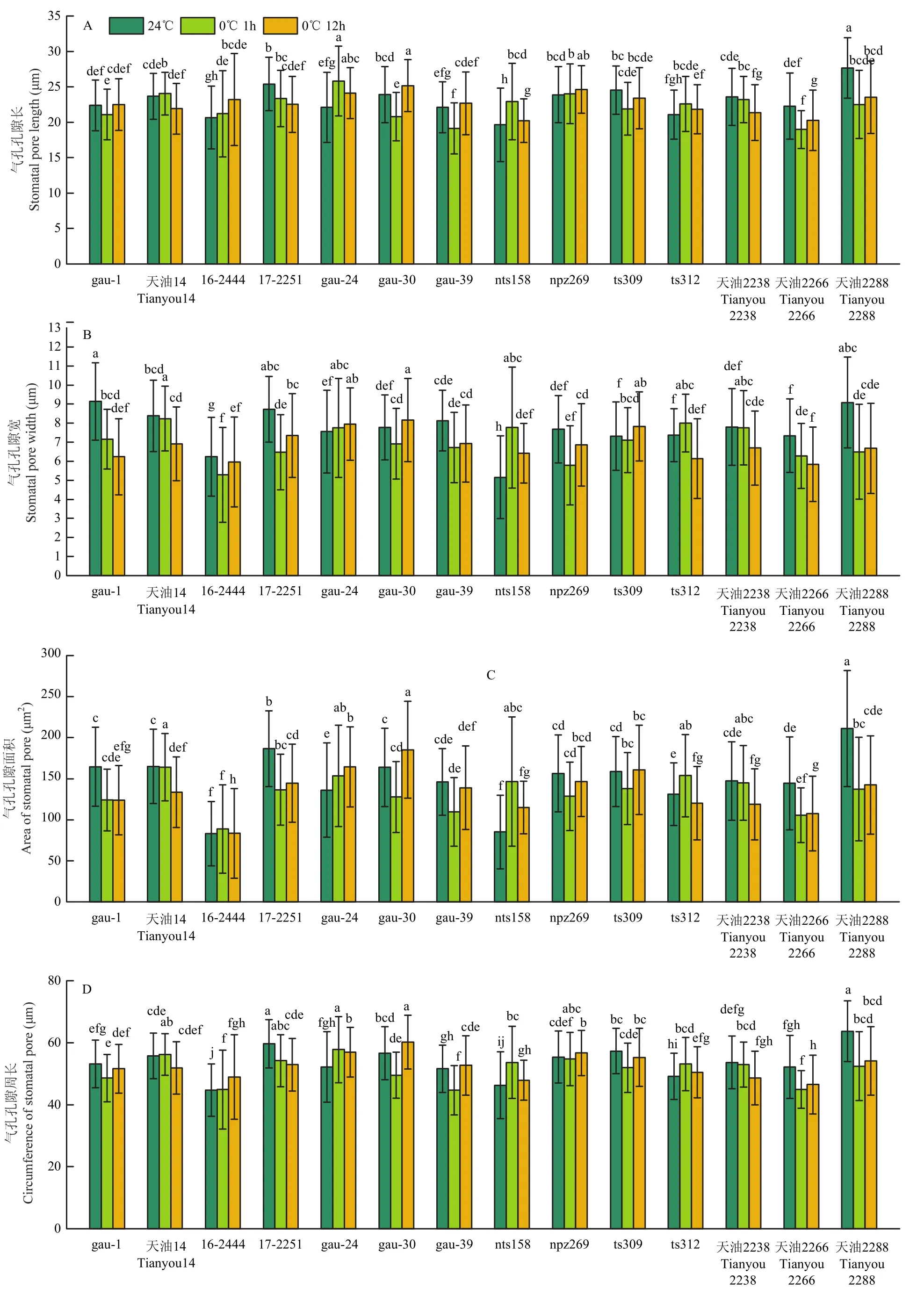
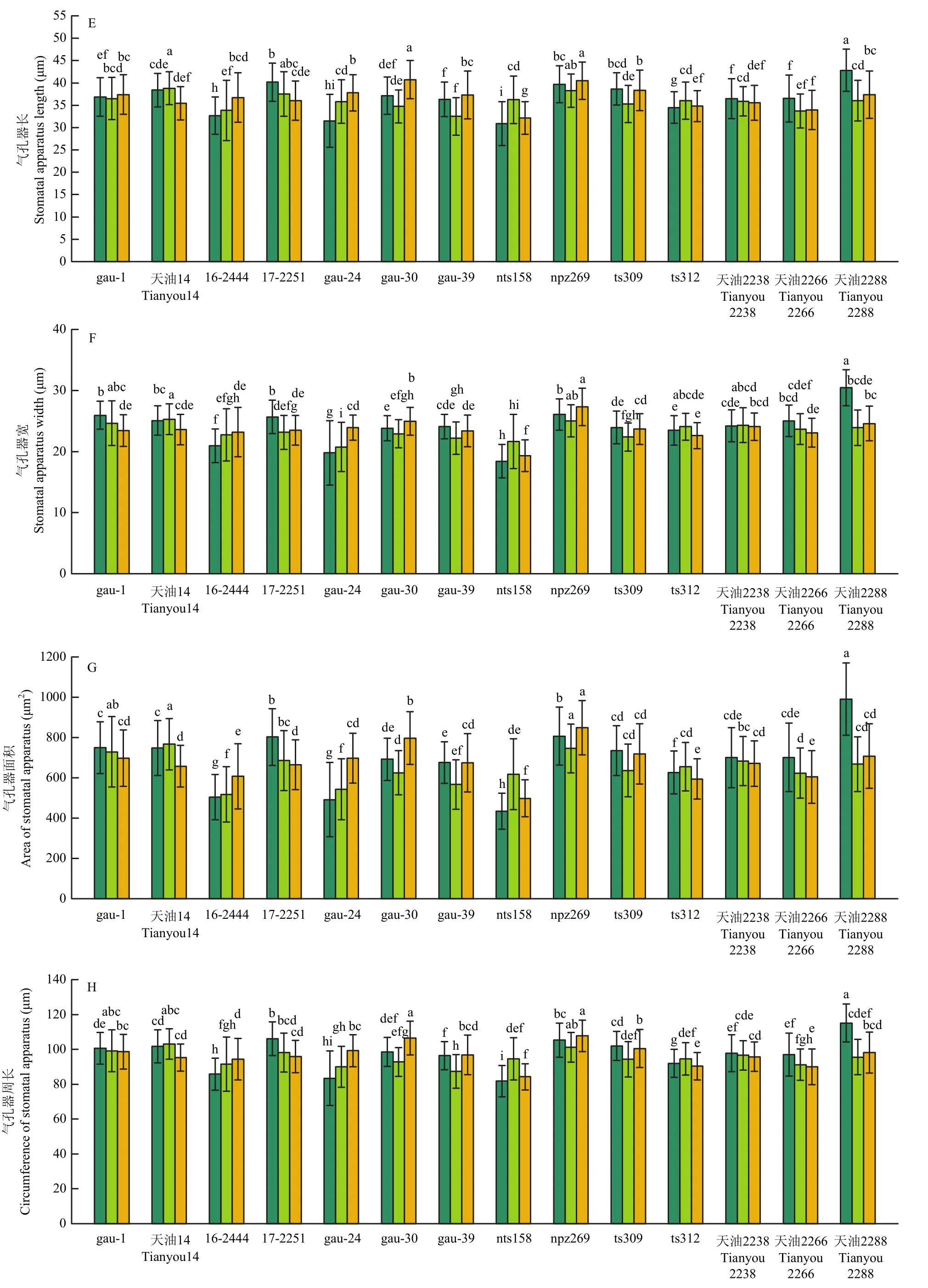
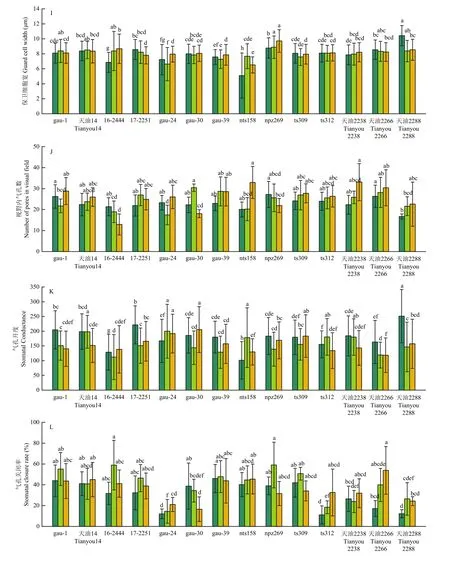
图3 甘蓝型冬油菜低温下的气孔运动Fig.3 Stomatal movement of winter Brassica napus at low temperature

表5 各指标测量的样本数量Table 5 The number of samples measured by each index
将强冬性品种gau-24、冬性品种天油14及弱冬性品种天油2288的气孔制片按不同处理排列于图4。24℃条件下全部品种视野内大部分(平均为68.99%)表现为张开,保卫细胞能快速对 0℃低温做出反应,在对照的基础上表现出部分气孔的关闭,视野中未关闭的气孔也有关闭趋势(气孔孔隙面积等指标较对照减小)。不同抗寒性品种在 0℃处理后气孔形态保持能力不同,强冬性品种gau-24在0℃处理12 h后1个气孔关闭;弱冬性品种天油2288在1 h处理后全部气孔的气孔孔隙均有明显的变小,12 h处理后关闭数量为4个;冬性品种天油14的气孔对低温的耐受性居中。
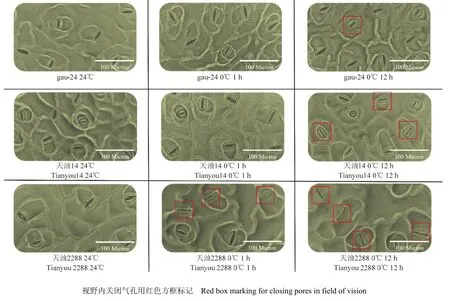
图4 甘蓝型冬油菜低温下的气孔图Fig.4 Stomatal of winter Brassica napus at low temperature
2.5 不同抗寒性甘蓝型冬油菜气孔特性综合评价
计算0℃处理1 h的12个气孔评价指标的耐寒系数(表6)。参试材料间耐寒系数差异显著(P<0.05),14个品种相同指标K值变化无明显规律。分析12个气孔评价指标的耐寒系数与品种抗寒性间的关系,进行相关性分析(表 7),除气孔关闭率与半致死温度间有P<0.05水平上的正相关外,其余指标间并无显著相关,不继续分析。

表6 0℃ 1 h耐寒系数评价Table 6 Coefficients of cold tolerance evaluated of treatment at 0℃ for 1 h
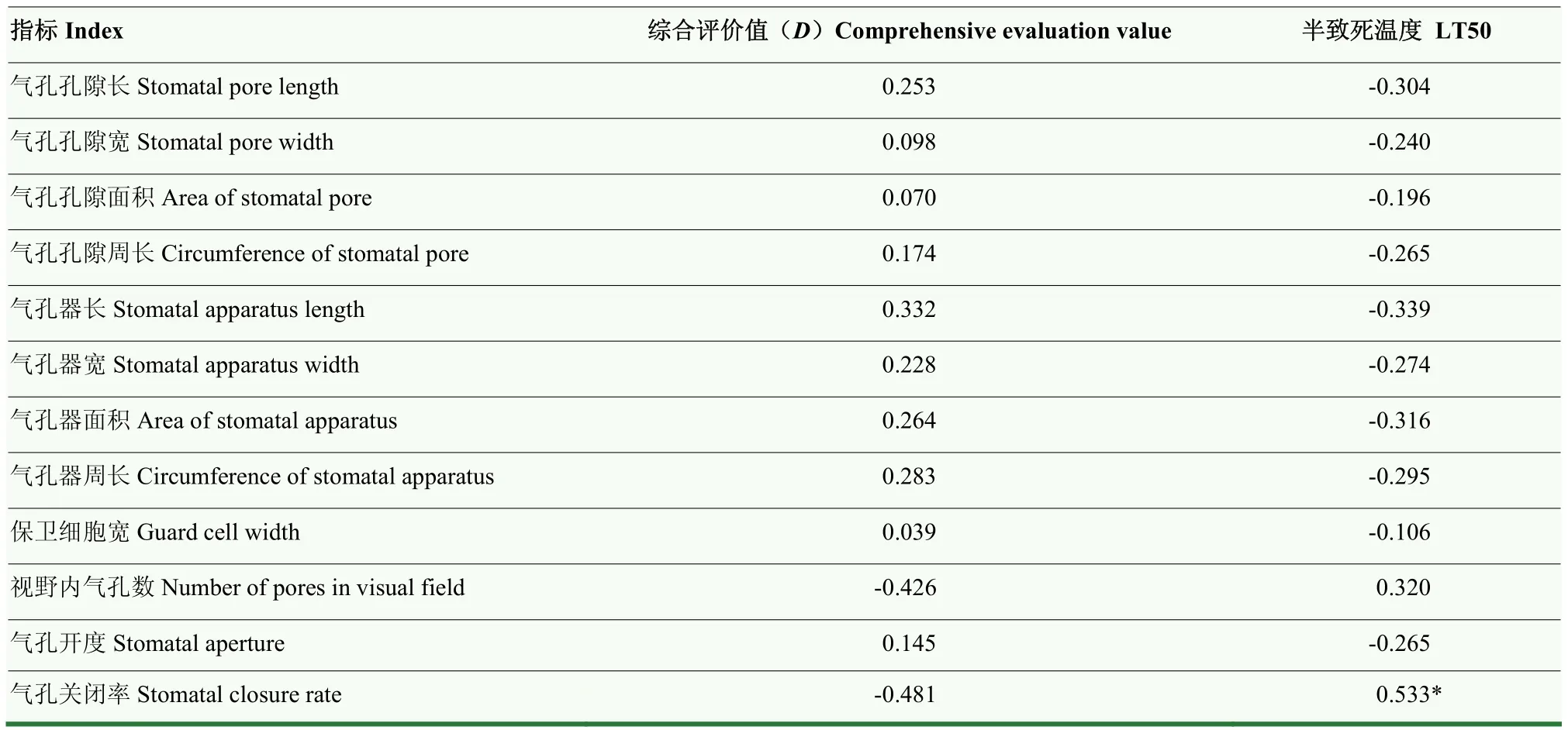
表7 0℃ 1 h抗寒性指标与气孔评价指标间的相关性Table 7 Correlation between cold resistance index and stomatal evaluation index
0℃处理12 h后,参试材料间的耐寒系数差异显著(P<0.05)。提取各指标K值较大的4个品种,高频率出现的品种有gau-24、gau-30、16-2444和nts158。求各品种 12个指标平均 K值,较大的 4个品种为nts158、gau-24、16-2444和gau-30,前后2种结果相同,但并不与抗寒性排序相近,所以需要进行主成分分析来综合评价低温下的气孔运动状态及分析各个指标对于综合评价值Z的影响(表8)。
根据耐寒系数列出各指标间的相关系数矩阵并计算相关系数矩阵R的特征值及特征向量,再使用提取成分的特征值和贡献率联系各成分下各指标中的因子载荷算得权重,特征值、因子载荷、贡献率,算得的权重列于表9。

表9 0℃ 12 h主成分分析相关指标Table 9 Indicators related to principal component analysis for 12 h at 0℃
12 h处理下的数据共提取出3个成分,3个成分的累积贡献率达92.690%,可以充分表述该项目。主要影响成分1的10个指标按因子载荷由大到小排
序为气孔器面积、气孔孔隙周长、气孔器周长、气孔器长、气孔器宽、气孔开度、气孔孔隙长、气孔孔隙面积、保卫细胞宽、气孔孔隙宽,其中气孔器面积、气孔孔隙周长、气孔器周长的值更接近于1,对成分1的影响更大。主要影响成分2的指标有视野内气孔数,影响成分3的主要指标为气孔关闭率。成分1的贡献率达到71.080%,故成分1中的主要影响指标就可以说明气孔的运动。贡献率为13.128%和8.482%的成分2和成分3对气孔运动的评价不能忽略。
对标准化后的耐寒系数赋予权重得到综合评价值Z(表10)。弱冬性品种天油2288的12个指标标准化的数据中9个为最小值0,综合评价值也为最小。对求得的综合评价值Z从大到小排序按从大到小排序为gau-24、nts158、16-2444、gau-30、npz269、ts309、gau-39、ts312、天油2238、gau-1、天油14、天油2266、17-2251、天油2288。其排序与抗寒性评价指标排序大致相同。
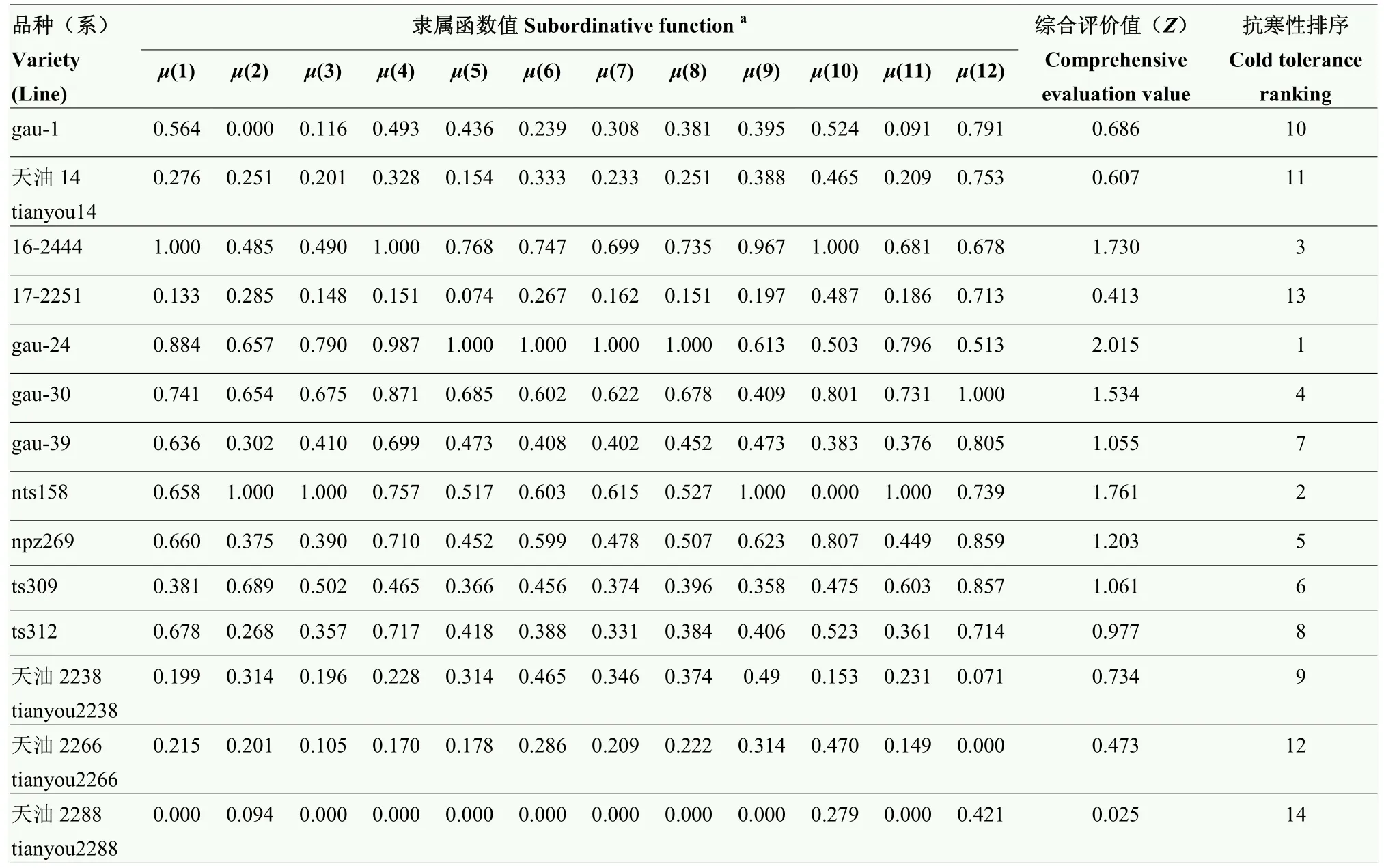
表10 0℃ 12 h标准化的数据和综合评价Table 10 0℃ for 12 h standardized data and comprehensive evaluation
分析抗寒性与低温下气孔运动的相关性是利用综合评价值D、半致死温度与12个指标及综合评价值Z之间的相关性(表11)。0℃ 12 h条件下,与综合评价值D相关性显著的指标有气孔孔隙长、气孔孔隙周长、气孔器长、气孔器面积、气孔器周长。与D值显著相关的指标均与半致死温度显著相关,气孔孔隙面积及综合评价值Z也与半致死温度显著相关,气孔孔隙周长及气孔关闭率与半致死温度极显著相关。

表11 0℃ 12 h抗寒性指标与气孔评价指标间的相关性Table 11 Correlation between cold resistance index and stomatal evaluation index at 0℃ for 12 h
3 讨论
3.1 逆境下的气孔运动评价
多个指标的气孔特性综合评价更能全面地解释和阐述气孔运动过程。何荆洲等[21]对 25个蝴蝶兰品种分类时综合了 13个气孔相关指标进行主成分分析后准确分类为大花型(花长>8 cm)、小花型(花长<4.5 cm)和花瓣无线纹或线纹非紫色三类。王孟珂等[22]在评估文冠果种质资源的抗旱性时采用了主成分分析加隶属函数的方法,从来自干旱和半干旱地区的 108份文冠果种质资源中选出了31份强抗旱种质资源。本研究采用主成分分析提取出3个成分,分析每个指标对不同成分的影响,发现其均对于气孔运动评价都有贡献,综合这些指标并赋予合适的权重能获得科学且适宜的评价。相关性分析结果,12个气孔相关指标的综合评价值与抗寒性间显著相关。在叶片生长过程中,保卫细胞的发育调控机制尚不完全清楚[23],但在此过程中外界环境因素及品种特性等均会改变下表皮细胞的发育方向[24]。低温处理会使气孔向不同方向或相同方向发生不同程度的变化[25]。因此,相较于只利用气孔性状的单个指标评价低温下气孔运动特性及与抗寒性的关系,综合评价更加全面客观。
3.2 利用耐寒系数评价气孔运动与抗逆性的关系
多数学者在利用气孔指标评价抗逆性时只考量静态气孔结构,有时会出现抗逆性评价结果相反的情况。如钟克友等[26]在研究水稻耐冷性时认为气孔较大的品种耐冷性较弱,赵跃龙等[27]在水稻上也得出了相同的结论。而李映晖等[28]对3个菠萝品种的抗寒性研究发现气孔大的品种抗寒性强。吴林等[29]认为气孔大小等单一气孔性状与越橘抗寒性无明显的关系。崔国文等[30]和王晖等[31]也认为气孔大小这一指标与植物抗寒性无显著相关。再如气孔密度,宋绪忠[32]在茶树上的试验结果认为气孔密度与抗寒性呈正相关。但李国华等[33]在澳洲坚果上的研究表明气孔密度小的抗寒性强。这些存在一定相互矛盾的结论从侧面印证了静态气孔结构评价抗寒性具有一定的片面性。本研究通过对比气孔静态结构评价指标在低温处理前后的相对变化,分析了不同抗寒性甘蓝型冬油菜气孔特性存在的差异。常温下及0℃处理1 h后12个气孔指标与半致死温度均无显著相关,0℃处理12 h后做相关分析发现气孔孔隙长、气孔孔隙宽、气孔孔隙面积、气孔孔隙周长、气孔开度5个指标与半致死温度存在显著相关。引入对照组使用耐寒系数后7个指标与抗寒性显著相关。
3.3 不同抗寒性品种间气孔特性差异
保卫细胞细胞壁不均一性[34]与微丝骨架[35]共同作用调节气孔运动过程中气孔形状的改变,根据气孔制片的观察及12个指标的统计分析结果,甘蓝型冬油菜气孔遭受低温胁迫后整体气孔(包括气孔孔隙与气孔器)变小。气孔动态变化及静态结构的差异构成不同抗寒性甘蓝型冬油菜品种的气孔特性。不同抗逆性的植物在应对逆境的调节过程中气孔特性不同[36],本试验统计的气孔关闭率表明强抗寒性品种低温处理后保持张开的比率高,强冬性甘蓝型冬油菜品种 0℃处理12 h后气孔孔隙长、气孔孔隙面积、气孔孔隙周长、气孔器长、气孔器面积、气孔器周长的耐寒系数及综合评价气孔特性的Z值明显大于冬性、半冬性及弱冬性甘蓝型冬油菜,表明强冬性品种气孔在低温处理后变化小,趋于保持张开。
3.4 甘蓝型冬油菜气孔运动与抗寒性的关系
植物对低温损伤的响应是通过改变相关组织或细胞结构实现的[36]。其中,气孔的开张是一个重要变化[5,37]。普遍的理论认为气孔运动与抗寒性间存在密切关系[38]。JURCZYK 等[14]研究发现耐寒性较差的大麦植株气孔导度会降低,相反抗寒性强的品种低温处理后气孔导度变化不大。同样,WILSON[39]研究发现抗寒性强的物种在低温下通常表现为气孔开放。本研究共观测14个品种低温下的12个气孔指标,发现甘蓝型冬油菜气孔运动确与抗寒性相关。研究显示12个单一鉴定指标中有7个与抗寒性指标显著相关,分别为气孔孔隙长、气孔孔隙面积、气孔孔隙周长、气孔器长、气孔器面积、气孔器周长、气孔关闭率,7个指标均与抗寒性正相关。同时,本试验在对0℃处理1和12 h材料的观测发现,1 h气孔运动并不与抗寒性相关,到12 h时相关性才显著。
气孔的运动与生理代谢有较强的关联作用,研究中常以生理指标为抗寒性评价的重要依据。有人认为植物体内大量的生理代谢引起细胞内环境稳态的变化,继而导致气孔的异常开闭[4]。影响气孔运动最直接的因素是保卫细胞水分含量的变化,DAVIES等[40]认为低温切断了根系供水导致了气孔关闭。试验对抗氧化酶、渗透调节物质及叶片相对含水量的测定结果显示低温处理后相对含水量高且能保持内环境稳定的品种气孔保持张开状态的能力更强。
4 结论
低温处理可显著影响甘蓝型冬油菜的气孔运动,且随低温处理时间的延长气孔趋向关闭的特征更为明显。同时,不同感温性品种其气孔的相对变化差异大,抗寒性较强的品种在低温处理后气孔保持张开的能力更强。
猜你喜欢
今日农业(2022年1期)2022-11-16 21:20:05
电脑迷(2022年10期)2022-11-08 02:41:30
西藏农业科技(2019年3期)2019-11-04 00:35:14
西藏农业科技(2019年1期)2019-07-25 00:37:02
福建林业科技(2019年1期)2019-04-10 03:15:44
西藏农业科技(2018年4期)2018-04-25 06:39:28
西藏农业科技(2018年4期)2018-04-25 06:39:26
江苏农业科学(2017年10期)2017-07-21 13:45:32
西南农业学报(2016年6期)2016-04-16 05:12:46
江苏农业科学(2015年7期)2015-08-20 11:59:23
