大肠杆菌型奶牛乳房炎对产奶性状相关基因表达的影响
2023-03-07 12:56:30李彦霞王晋鹏冯芬包斌武董益闻王兴平罗仍卓么
生物技术通报 2023年2期
李彦霞 王晋鹏 冯芬 包斌武 董益闻 王兴平 罗仍卓么
(1. 宁夏大学农学院,银川 750021;2. 宁夏回族自治区反刍动物分子细胞育种重点实验室,银川 750021)
大肠杆菌(Escherichia coli)是引起奶牛乳房炎的主要病原菌之一,感染奶牛乳房时会引起急性临床乳房炎,造成奶牛乳腺上皮细胞(bovine mammary epithelial cells,bMECs)的乳合成能力和分泌功能障碍,导致牛奶品质显著下降[1-3]。研究表明,即使患病奶牛的临床症状和致病菌消失,其乳腺也不会迅速恢复到原状[4],这将导致产奶量持续受到影响。除了上述影响,乳房炎的致病菌还会诱导炎症因子表达,产生炎症反应。脂多糖(lipopolysaccharide,LPS)作为E.coli表面的活性成分,是E.coli型乳房炎的主要致病因子[5],它可以引起肿瘤坏死因子-α(tumor necrosis factor-α,TNF-α)、白细胞介素-6(interleukin-6,IL-6)和白细胞介素-1β(interleukin 1β,IL-1β)等基因在奶牛乳腺组织和bMECs 炎症过程中表达量升高[6]。此外,乳房炎也可引起外周血液[7]和乳腺组织[8]的基因表达量发生变化,进而严重影响了奶牛的产奶量和乳品质。因此,从分子和细胞水平上研究抑制产奶量减少和乳品质下降的原因,是当下学者们研究的热点领域,为高产优质奶牛的分子育种奠定基础。
在奶牛产奶性状方面,学者们挖掘了许多与奶牛产奶量、乳蛋白率和乳脂率相关的基因,如脂肪酸结合蛋白3(fatty acid binding protein 3,FABP3)、脂蛋白脂酶(lipoprotein lipase,LPL)、α-酪蛋白家族(casein alpha-S1,CSN1S1)、β-酪蛋白(casein beta,CSN2)、α-乳白蛋白(lactalbumin alpha,LALBA)、乳脂球表皮生长因子8(milk fat globule-EGF factor 8 protein,MFGE8) 基因。FABP3和LPL主要与乳脂相关,其中FABP3调控乳脂主要合成通道,LPL参与乳脂合成过程和代谢过程[9-10];牛奶蛋白主要由4 种酪蛋白和乳清蛋白组成,CSN1S1和CSN2是乳蛋白组成成分中的两种酪蛋白,而LALBA是乳蛋白组成成分中的乳清蛋白[11]。MFGE8主要与炎症以及产奶量相关[12-13]。虽然已经发现上述基因在奶牛乳合成中发挥着重要作用,但是对于它们在E.coli型奶牛乳房炎和bMECs 炎症反应中的表达模式目前还尚不清楚,有待进一步研究。
因此,本研究采用qPCR 技术检测了CSN1S1、CSN2、LALBA、FABP3、LPL和MFGE8基因在E.coli型乳房炎奶牛乳腺组织和bMECs 炎症反应中的表达模式,以期为解析E.coli型乳房炎奶牛的产奶量、乳蛋白率及乳脂率下降的原因,以及为高产优质奶牛的分子育种提供理论依据。
1 材料与方法
1.1 材料
本实验所用的乳腺组织为2-3 岁健康奶牛和E.coli诱导乳房炎的荷斯坦奶牛的乳腺组织,为实验室前期采集、鉴定和冻存[8]。其中E.coli诱导乳房炎的荷斯坦奶牛的乳腺组织是采用无菌手术法活体采集奶牛的乳腺组织,主要步骤:将E.coli冻干粉溶解,扩大培养,用PBS 稀释,用通乳针将E.coli悬浮液经乳导管一次性注入到试验组奶牛的右后乳房中,在接种后的第7 天,采集其乳腺组织[14];bMECs 为课题组前期鉴定并保存于液氮罐中的细胞[15]。
实验所用的主要仪器:CO2培养箱(Thermo,美国)、CFX96 实时荧光定量PCR 仪(Bio-Rad,美国)、高压灭菌锅(普和希,日本)、电热恒温水浴锅(一恒,上海)、普通PCR 仪(Bio-Rad,美国)。
实验所用的主要试剂:DMEM/F12 细胞培养基(Hyclone,美国)、0.25%的胰酶消化液(Hyclone,美国)、PBS 缓冲液(Hyclone,美国)、胎牛血清(BI,以色列)、脂多糖(Sigma,美国)、Real time-荧光定量PCR 试剂盒(聚合美,北京)、逆转录试剂盒(TaKaRa,北京)、TriZol 试剂(TaKaRa,北京)、RNase-free H2O(天根,北京)。
1.2 方法
1.2.1 bMECs 的复苏与传代培养 从液氮中取出bMECs,37℃水浴锅解冻;接种在含有10%胎牛血清的DMEM/F12 培养液中,在37℃、5% CO2和100%湿度的培养箱中培养;待细胞融合度为90%时传代,即用PBS 漂洗细胞1-2 遍,再用胰酶消化液吹打消化2-3 min,加入2 mL 培养基终止消化;转移至15 mL 离心管中,1 000 r/min 离心5 min;弃上清,加入1 mL 培养基吹打混匀后,接种到新培养基中培养。
1.2.2 bMECs 炎症模型的建立 将bMECs 接种于6孔细胞培养板中培养24 h,待细胞融合度达60%-70%时,向每孔细胞中滴加LPS 溶液(终浓度为50 ng/μL),诱导细胞产生炎症反应[16]。在LPS 诱导的0、3、6、12 和24 h,分别收集细胞,用于RNA 提取。通过检测细胞炎症因子mRNA 的表达量,判断炎性细胞模型是否构建成功。
1.2.3 RNA 提 取 与cDNA 合 成 采 用TriZol 法 分别提取约1 cm3的健康奶牛右后乳区的乳腺组织和E.coli诱导乳房炎奶牛右后乳区的乳腺组织,以及LPS 诱导不同时间(0、3、6、12 和24 h)bMECs的RNA。电泳检测RNA 的完整性,全波长酶标仪检测RNA 的浓度与纯度(1.8 < OD260/280< 2.0)。
采用PrimeScriptTMRT Reagent Kit with gDNA Eraser 逆转录试剂盒,按照说明书,通过两步反应(去除基因组DNA 反应和反转录)合成cDNA,以此cDNA 为模板,进行下一步实验。多功能酶标仪检测cDNA 的浓度,稀释至100 ng/μL,-20℃冻存。
1.2.4 qPCR 反应 参照GenBank 数据库中各mRNA的序列信息,利用Primer premier 5.0 软件设计qPCR引物(表1),委托通用生物(安徽)有限公司合成,RNase free H2O 溶解。
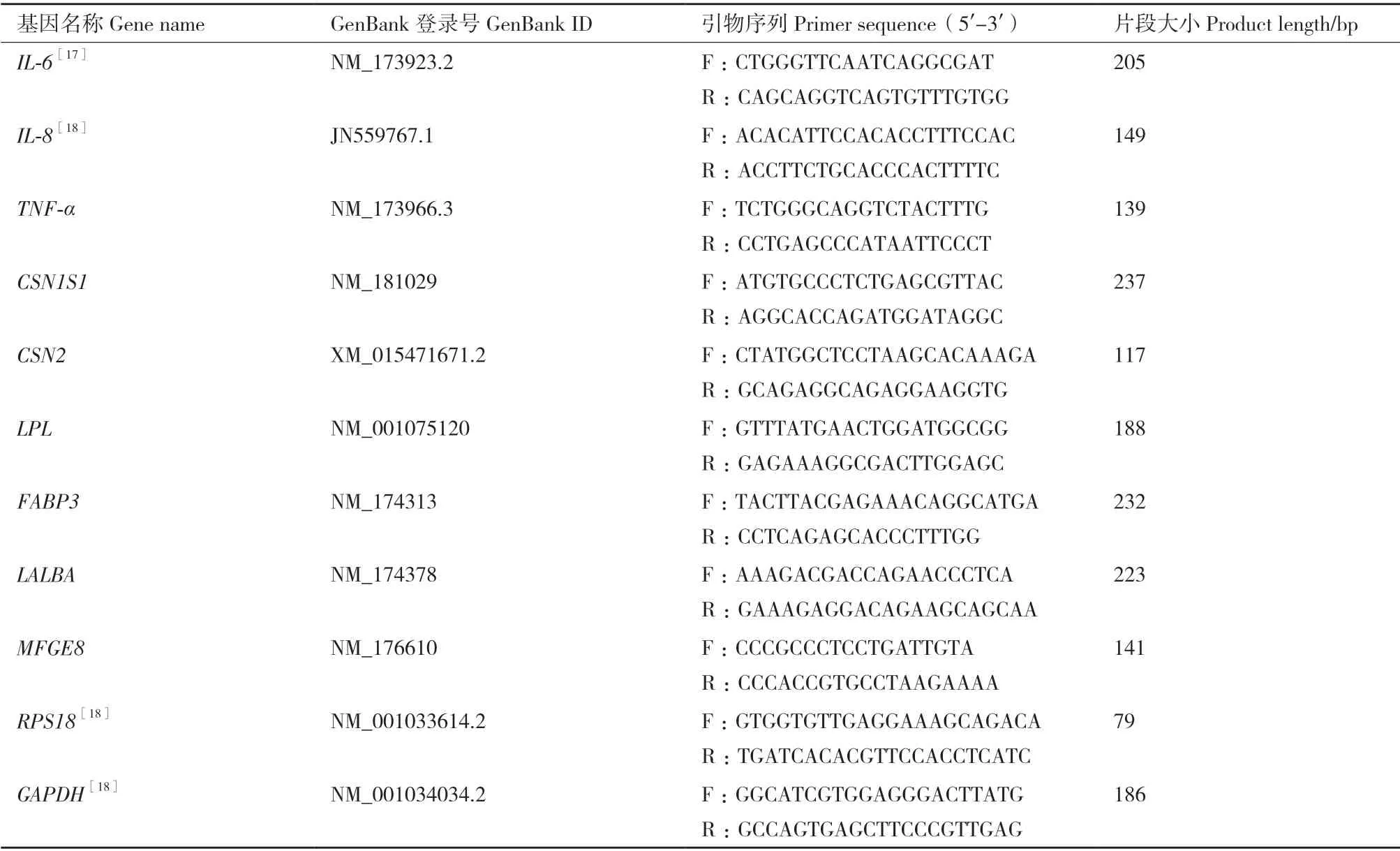
表1 qPCR 引物信息Table1 qPCR primer information
qPCR 的反应体系:2×M5 Hiper SYBR Premix Es Taq (with Tli RNaseH) 10.0 μL,100 μmol/L 的 上下游引物各0.8 μL,DNA 模板1.5 μL,加灭菌去离子水至20.0 μL。qPCR 的反应程序:(预变性95℃30 s,95℃ 5 s,60℃ 30 s)40 个 循 环;95℃ 10 s,65℃ 5 s。
1.2.5 数据分析 采用2-△△ct方法计算基因的相对表达量(n=3),所有数据均以“平均值±标准差”表示。利用GraphPad 8.3 软件的单因素方差(one-way AVONA)进行组间基因表达水平的差异显著性分析,P< 0.05 表示差异显著,有统计学意义。
2 结果
2.1 候选基因在E.coli型乳房炎奶牛的乳腺组织中的表达分析
2.1.1 乳蛋白和泌乳量相关候选基因在奶牛乳腺组织中的表达分析 利用qPCR 技术检测了乳蛋白和泌乳量相关的CSN1S1、CSN2和LALBA基因在健康奶牛和E.coli型乳房炎奶牛的乳腺组织中的表达量。结果表明,与健康奶牛相比,乳蛋白相关的CSN1S1、CSN2和LALBA基因在E.coli型乳房炎奶牛的乳腺组织中的mRNA 表达量均极显著下调(P<0.000 1)(图1)。该结果提示,在奶牛E.coli型乳房炎发生后,上述3 个基因表达量下降可能导致牛奶的乳蛋白含量及泌乳量减少。
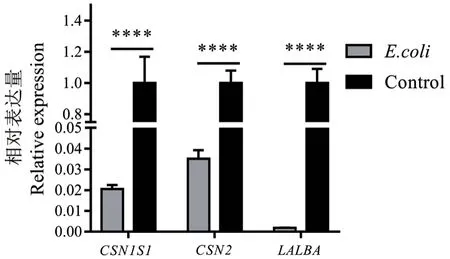
图1 CSN1S1、CSN2 和LALBA 基因在E.coli 型乳房炎奶牛的乳腺组织中的表达量Fig. 1 Expressions of CSN1S1, CSN2 and LALBA gene in the cow mammary tissues of dairy cows with E.coli type mastitis
2.1.2 乳脂调控相关候选基因在奶牛乳腺组织中的表达分析 利用qPCR 技术检测了乳脂调控相关的FABP3和LPL基因在健康奶牛和E.coli型乳房炎奶牛的乳腺组织中的表达量。结果表明,与健康奶牛相比,上述2 个基因在E.coli型乳房炎的乳腺组织中的表达量都极显著下调(P< 0.000 1)(图2)。上述结果提示,FABP3和LPL基因表达量下调可能导致了在E.coli型乳房炎奶牛分泌的牛奶乳脂减少和乳脂率降低。

图2 FABP3 和LPL 基因在E.coli 型乳房炎奶牛的乳腺组织中的表达量Fig. 2 Expressions of FABP3 and LPL gene in the mammary tissues of dairy cows with E.coli mastitis
2.1.3 炎症和泌乳量相关MFGE8基因在奶牛乳腺组织中的表达分析 利用qPCR 技术检测了MFGE8基因在健康奶牛和E.coli型乳房炎奶牛的乳腺组织中的表达量。结果表明,与健康奶牛相比,MFGE8基因在E.coli型乳房炎的乳腺组织中的表达量都极显著下调(P< 0.000 1)(图3)。结果提示,MFGE8基因的表达量降低导致抗炎反应减弱,炎症反应加剧,最终乳腺组织的功能减弱,导致产奶量减少。
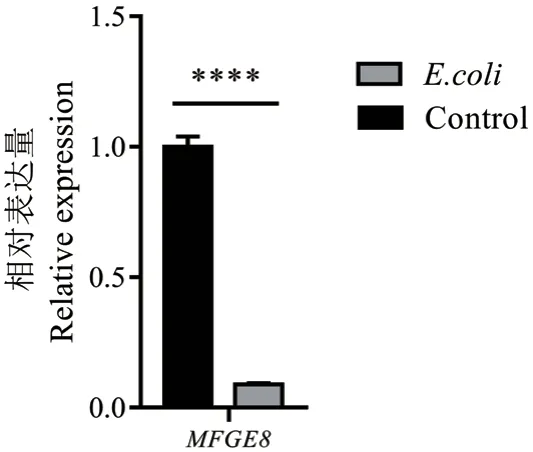
图3 MFGE8 基因在E.coli 型乳房炎奶牛的乳腺组织中的表达量Fig. 3 Expression of MFGE8 gene in mammary tissue of dairy cows with E.coli mastitis
2.2 bMECs炎症模型构建的验证结果
本研究通过体外培养bMECs(图4-A),利用LPS 诱导bMECs 构建细胞炎症模型。为验证炎症细胞模型的可靠性,利用qPCR 技术检测了LPS 诱导不同时间细胞内主要炎症因子基因的表达情况。结果表明,与0 h(对照组)相比,炎症因子TNF-α在6 h 的表达量极显著上调(P< 0.000 1)(图4-B);炎症因子IL-6在3、6 和12 h 时的表达量极显著上调(P< 0.01)(图4-C);炎症因子IL-8在6 h、12 h 和24 h 的表达量极显著上调(P< 0.01)(图4-D),说明本实验成功构建了bMECs 炎症模型。
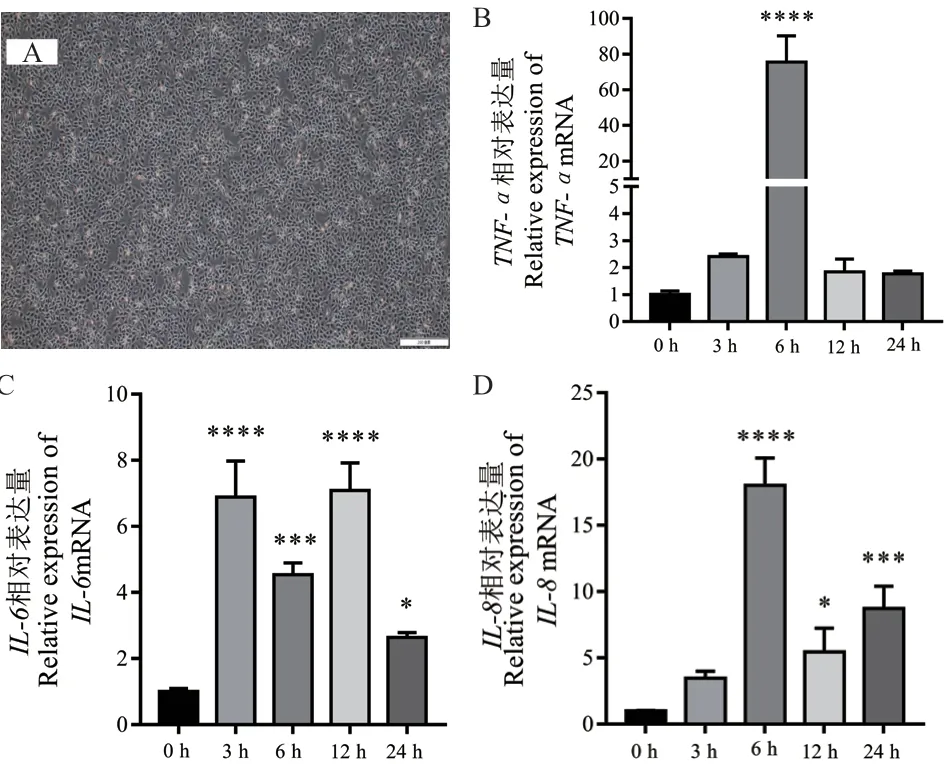
图4 主要炎症因子基因在LPS 诱导bMECs 不同时间的表达量Fig. 4 Expression of main inflammatory factor genes in LPS-induced bMECs at different times
2.3 候选基因在LPS诱导bMECs炎症反应中的表达分析
2.3.1 乳蛋白和泌乳量相关候选基因在bMECs 炎症反应中的表达量 为揭示乳蛋白和泌乳量相关的CSN1S1、CSN2及LALBA基因在bMECs 炎症反应中的动态表达情况,采用qPCR 技术检测了它们在LPS诱导bMECs 产生炎症反应不同时间的表达量。结果表明,与对照组(0 h)相比,CSN1S1、CSN2及LALBA基因的表达量都显著降低,其中CSN1S1在LPS 诱导3、6、12、24 h 后的表达量都极显著下调(P< 0.000 1)(图5-A);CSN2在LPS 诱 导3 h 后表达量降低,但未达到显著效果,当LPS 诱导6、12、24 h 时,CSN2的表达量极显著下调(P<0.000 1)(图5-B);LALBA在LPS 诱 导3、6、12 h表达量显著下调(P< 0.05),而24 h 时表达量降低,但差异不显著(P> 0.05)(图5-C)。结果提示,在LPS 诱导细胞产生炎症后,下调了上述基因的表达,可能降低了bMECs 中乳蛋白的合成与乳汁分泌量。
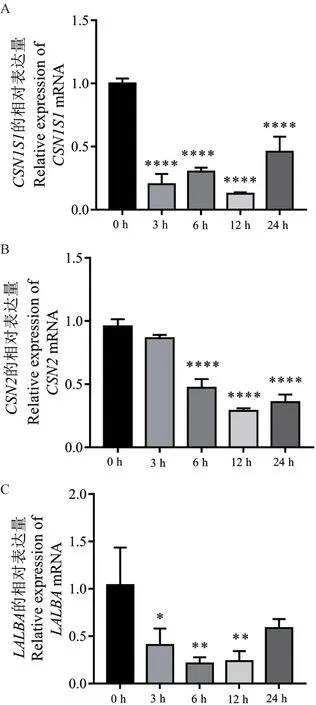
图5 乳蛋白相关基因在LPS 诱导bMECs 不同时间的表达量Fig. 5 Expressions of milk protein-related genes in LPSinduced bMECs at different times
2.3.2 乳脂调控相关候选基因在bMECs 炎症反应中的表达量 为探讨乳脂相关基因FABP3和LPL在bMECs 炎症反应中的动态表达情况,采用qPCR 技术检测了它们在LPS 诱导bMECs 不同时间的表达量,结果表明,与对照组(0 h)相比,FABP3和LPL在LPS 诱导bMECs 产生炎症反应的3、6、12和24 h 的表达量均显著下调,其中FABP3在3、6、12 和24 h 的表达量极显著下调(P< 0.000 1)(图6-A);LPL在3 h 的表达量显著下调(P< 0.05),在6、12 和24 h 的表达量极显著下调(P< 0.000 1)(图6-B)。结果提示,在LPS 诱导bMECs 产生炎症过程中上述基因表达量下调,可能导致了乳中的乳脂率降低。
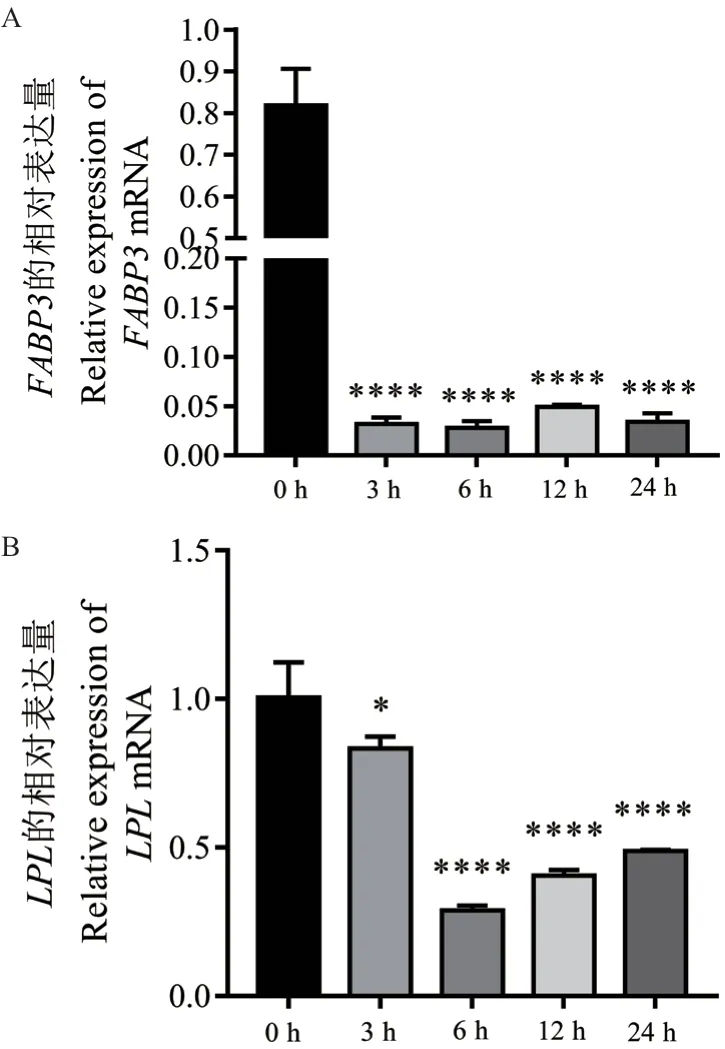
图6 乳脂调控相关基因在LPS 诱导bMECs 不同时间的表达量Fig. 6 Expressions of milk fat regulation related genes in LPS-induced bMECs at different times
2.3.3MFGE8基因在bMECs 炎症反应中的表达量 为探讨基因MFGE8在bMECs 炎症反应中的动态表达情况,采用qPCR 技术检测了其在LPS 诱导bMECs 不同时间的表达量,结果表明,与对照组(0 h)相比,MFGE8在3、12 和24 h 的表达量极显著下调(P< 0.001),在6 h 的表达量也极显著下调(P< 0.000 1)(图7)。结果提示,MFGE8表达量降低,吞噬凋亡细胞的能力下降,抗炎反应减弱,导致bMECs 炎症反应的抵抗力下降,乳合成能力和泌乳量减少。
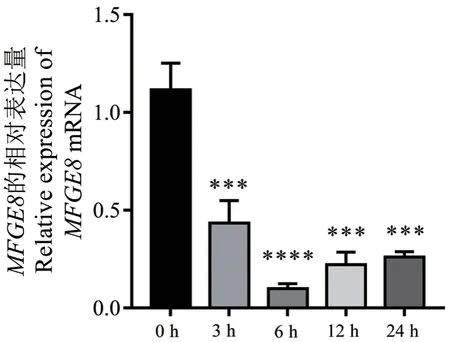
图7 MFGE8 基因在LPS 诱导bMECs 不同时间的表达量Fig. 7 Expressions of MFGE8 gene in LPS-induced bMECs at different times
3 讨论
乳房炎作为奶牛最常见的疾病之一,患病后奶牛健康状况急剧下降,泌乳量减少,但是其分子层面的原因尚不清楚。乳腺组织是奶牛泌乳的主要功能器官,而bMECs 是泌乳的主要功能细胞,也是乳腺防御病原菌入侵的第一道防线。LPS 作为E.coli的主要毒力因子,可诱导bMECs 产生固有免疫应答和炎症反应,并且可导致细胞损伤和凋亡[19]。当奶牛乳腺组织受到E.coli刺激时,其生理状态发生改变,表现出乳房炎临床症状,产奶量明显降低,牛奶体细胞数上升[20]。值得一提的是,乳腺组织发生炎症反应时,除了bMECs 之外,其他免疫细胞也参与炎症反应[14]。此外,有研究发现,LPS 诱导的乳房炎症反应可影响乳蛋白和乳脂的合成[21-22]。因此,为全面探讨E.coli型乳房炎中泌乳量、乳蛋白和乳脂率下降的原因,本研究从组织水平(E.coli型乳房炎奶牛和健康奶牛的乳腺组织)和细胞水平(LPS诱导的bMECs)2 个层面研究了6 个泌乳性状相关候选基因(CSN1S1、CSN2、LALBA、FABP3、LPL和MFGE8)的表达情况,结果发现上述6 个候选基因在炎症前后的组织水平和细胞水平的表达量变化是完全一致的,说明这6 个基因参与了乳腺组织和bMECs 的泌乳或炎症反应过程。
据报道,牛奶中酪蛋白编码的基因CSN1S1、CSN2和LALBA的表达量与产奶量显著相关[23]。CSN1S1和CSN2是奶牛泌乳性状的关键基因[24],两者相互作用、相互影响。Wu 等[25]研究发现,在肽聚糖(peptidoglycan, PGN)和脂磷壁酸(lipoteichoic acid, LTA)引起的奶牛乳房炎中,CSN1S1和CSN2基因的mRNA 表达量降低,并通过降低组蛋白H3乙酰化,减少了泌乳量。LALBA是泌乳期的bMECs产生的一种对牛奶和乳腺具有特异性的牛奶蛋白[26],与溶菌酶具有高度同源性[27],可以通过催化乳糖合成进而调控乳产量[28]。在小鼠乳腺中注射LPS 24 h 内,LALBA的表达量显著降低,其表达量降低可能是通过LPS 诱导核因子-κB(NF-κB)和细胞因子的产生来介导[29]。本研究中也发现CSN1S1、CSN2和LALBA基因在E.coli型乳房炎奶牛乳腺组织和炎性细胞模型中的表达量显著降低。结合上述前人的研究结果,推测CSN1S1、CSN2和LALBA基因的表达量降低,可能是引起E.coli型乳房炎产奶量和乳蛋白率降低的原因。
乳脂是牛奶的主要成分,也是乳品质的主要检测指标之一。在乳脂代谢调节的研究中,学者们发现LPL和FABP3是参与乳脂合成与分解代谢过程中的关键蛋白[30-31]。FABP3作为结合脂肪酸的FABP家族成员之一,可协同其他基因形成一个调控奶牛乳腺脂肪酸向乳脂合成的通道[9]。已有研究表明,FABP3与乳脂合成信号通路中的相关转录因子成正相关关系[32]。本研究发现,FABP3在E.coli型乳房炎奶牛的乳腺组织和炎症细胞模型中的表达量显著下调。结合前人的研究结果,推测在E.coli型奶牛乳房炎中,FABP3基因表达下调,抑制了乳脂合成,降低了牛奶的乳脂率。
LPL 是脂肪代谢的关键酶,催化产生甘油三酯,并将脂蛋白水解为脂肪酸和单甘油三酯,参与了脂肪酸合成和组织吸收利用的整个过程[10,33-34]。乳腺上皮细胞可以生成LPL,哺乳期LPL的酶活性较高,增加了乳腺中脂肪酸的释放[31]。与健康奶牛的乳汁相比,乳房炎奶牛的乳汁中游离脂肪酸含量较高,但LPL活性较低[35]。与上述研究结果一致,本研究中LPL基因在E.coli型乳房炎奶牛的乳腺组织和炎性细胞模型中的表达量也显著降低,这可能导致E.coli型乳房炎奶牛的乳腺中LPL的活性降低,抑制了脂肪酸的释放,使得牛奶的乳脂率降低。
MFGE8基因在乳腺组织、乳腺上皮细胞及巨噬细胞中均存在[36],但在乳腺上皮细胞中以乳糖蛋白的形式大量存在,且与泌乳量密切相关。此外,它还具有抗菌和抗病毒作用,在乳腺退化期间可清除凋亡的乳腺上皮细胞[12-13]。当缺乏MFGE8时,导致bMECs 凋亡和MFGs 的清除功能延迟,以及乳腺开始退化和产生炎症反应[37]。以上研究表明,MFGE8在乳腺组织中发挥着重要作用。该结果与本实验结果相似,即在E.coli型乳房炎发生或在bMECs 产生炎症后,乳腺组织和bMECs 中MFGE8的表达量显著降低。结合前人结果,推测MFGE8的表达量降低可能使得吞噬细胞凋亡能力降低,抗炎反应减弱,炎症反应加剧,最终乳腺组织的功能减弱,导致乳脂合成和产奶量减少。
4 结论
E.coli型奶牛乳房炎发生和LPS 诱导bMECs 产生炎症后,产奶相关基因CSN1S1、CSN2、LALBA、FABP3、LPL和MFGE8的表达量均显著下降,可能导致乳蛋白合成减少、脂肪酸释放降低和产奶量下降,解释了E.coli型乳房炎发生过程中产奶量减少和乳脂率降低的原因,研究结果可为高产优质的奶牛分子育种提供参考。
猜你喜欢
现代畜牧科技(2021年11期)2021-12-21 06:11:16
现代畜牧科技(2021年2期)2021-03-19 07:48:22
中国生殖健康(2020年5期)2021-01-18 02:59:38
中国生殖健康(2020年2期)2021-01-18 02:51:38
中国生殖健康(2020年2期)2021-01-18 02:51:16
今日农业(2020年24期)2020-12-15 16:16:00
中国乳业(2019年12期)2020-01-17 08:47:40
中国生殖健康(2018年2期)2018-11-06 07:10:38
分析化学(2018年4期)2018-11-02 10:16:04
农民致富之友(2018年18期)2018-07-14 05:54:18