
丛枝菌根真菌(AMF)对蒙古沙冬青幼苗的促生特性及作用机制
2023-03-07 09:04:32王紫瑄解甜甜王雅茹杨杰艳杨秀清
干旱区研究 2023年1期
王紫瑄, 解甜甜, 王雅茹, 杨杰艳, 杨秀清
(山西农业大学林学院,山西 晋中 030801)
丛枝菌根真菌(Arbuscular Mycorrhizae Fungi,AMF)是分布最为普遍的菌根真菌类型,在绝大多数陆生植物根系皮层细胞间或细胞内增大形成丛枝状结构,与植物建立菌根共生体[1-3]。AMF作为植物磷素高效利用密切相关的微生物类群,因其共生特性,可在植物-AMF 互作(根际)、AMF-微生物互作(菌丝际)两个界面同时影响植物养分吸收利用[4],因此,菌根共生领域发展成为土壤学、生态学、植物学等学科国内外学者广泛关注和持续研究的重要研究领域之一[5]。2022 年New Phytologist 上报道了AMF 异形根孢囊霉中一个含有SPX 结构域的磷酸盐转运蛋白RiPT7 参与AMF 共生界面磷平衡与输出过程,调节菌根共生体发育和提高菌根植物对土壤磷的摄取[6]。同时,共生的真菌可通过增加根系表面积和体积扩展植物根系[7],扩大根系吸收面积,加强矿质养分的空间有效性,提高植物对土壤养分的吸收和利用[8]。此外,AMF 可通过与植物根系形成的菌根共生体诱导共生植物释放土壤酶等物质,活化土壤中被固定的矿质养分,提高矿质养分有效浓度[9],从而改善土壤质量[10]、促进根系养分吸收,在调节植物与土壤间以及植物体内营养平衡等方面均具有重要的生态功能[11]。有学者发现,菌根联合可以影响宿主植物根系分泌螯合物[12]、提高植株渗透调节能力[13]、促进光合作用[14]以及增强植物竞争力[15]等,从而影响植物形成及构建[16]。这些研究有助于深入认识AMF 对植物的促生长作用。目前在玉米[17]、甘草[18]、苜蓿[19-20]、黄花蒿[21]等的研究中已经表明,AMF对这些植物生长及生物量提高均有促进作用。
蒙古沙冬青(Ammopiptanthus mongolicus)隶属豆科蒙古沙冬青属,是荒漠地区特有的旱生阔叶观花灌木和孑遗物种[22],在防风固沙和荒漠植被恢复中发挥着重要生态作用[23],也是北方少有的重要常绿树种资源。但由于蒙古沙冬青属直根系,主根长、侧根不发达导致其苗木再生性差,移栽造林成活率低,资源利用非常困难[24]。蒙古沙冬青当前分布范围较小,主要在内蒙古乌海、贺兰山、鄂托克旗、阿拉善等地区呈斑块状分布,种质资源岌岌可危。AMF 与植物菌根联合作用对植物根系及植株生长的影响为解决蒙古沙冬青根系局限性导致苗木成活及生长受限的苗木培育瓶颈问题提供了借鉴。是否可通过AMF 侵染蒙古沙冬青幼苗根系形成菌根共生,改善其生理代谢过程及其根际土壤环境,从而有利于其壮苗培育和移栽成活率的提高。
鉴于此,本试验分析两种AMF不同接菌方式以及未接菌处理下蒙古沙冬青幼苗根系及植株生长和生物量的变化,根系及叶片生理特性、根际土壤酶活性变化及其与幼苗生长和生物量变化的关联性,旨在研究AMF对蒙古沙冬青幼苗的促生长作用及其影响机制。结果可为探索蒙古沙冬青菌根化种苗培育技术,提高蒙古沙冬青造林成活率,实现蒙古沙冬青资源高效利用提供实践依据和技术支撑。
1 材料与方法
1.1 供试材料
供试种子采自甘肃民勤当年成熟收集的蒙古沙冬青种子。供试菌种由北京市农林科学院丛枝菌根真菌种质资源库(BGC)提供,选取两种AMF:摩西斗管囊霉Funneliformis mosseae(XJ02),根内根孢囊霉Rhiaophagus intraradice(BJ09)。每克菌剂的孢子数约为40~200个。以白三叶草为寄主,灭菌后的黄沙为基质,在光照培养箱内(培养温度为25 ℃,光照强度为15000 lx,光照时间为15 h·d-1)进行AMF扩繁。3个月后,保留含有孢子、菌丝及侵染根段的黄沙,作为接种菌剂。
试验所用土壤取自山西农业大学植物园(0~10 cm)。去除大颗粒石子和根系,经自然风干后过筛混合,经高压蒸汽灭菌后作基质待用。
1.2 播种育苗试验
试验设置4 个接菌处理,每个处理10 次重复。分别为单一接菌(根内根孢囊霉,R.i;摩西斗管囊霉,F.m)、混合接菌(根内根孢囊霉+摩西斗管囊霉为1:1 的混合菌剂,R.i+F.m)和未接菌处理(未接菌,CK)。花盆(13 cm×12 cm)中放入灭菌土壤基质和接种菌剂(体积比为1:3)至盆高约4/5 处,将消毒后的20粒蒙古沙冬青种子均匀播入其中,其表面再用灭菌基质覆盖约2 cm,每盆质量控制为1.0 kg,置于培养箱中(温度25 ℃,光照强度2500 lx)培养。期间每3 d对各处理进行称重记录,补充蒸发的水分,保持水分含量在5%。幼苗培养30 d后进行各项指标测定。
1.3 指标测定
1.3.1 根系菌根侵染率及菌根依赖性测定 将接种
AMF 孢子30 d 后的蒙古沙冬青植株根部用去离子水洗净,剪刀剪至1 cm左右的长度,置于50 mL离心管中,加入30 mL 左右10%KOH(w/v)溶液,并置于90 ℃水浴锅中水浴30 min。蒸馏水冲洗三次,加入2%HCl酸化5 min,沥干后加入0.05%的台盼蓝染色液15 mL,混匀后置于90 ℃水浴锅中水浴加热15 min,沥干染色液,加入丙三醇40 mL,常温脱色2 d左右后制成封片于显微镜(型号:奥林巴斯CX31)下观测,检测幼苗根系内菌根侵染状况,参照Phillips等[25]采用网格交叉法统计形成菌根的根段数,计算根系的AMF侵染率。根据获得的地上部干重、根干重等计算菌根依赖性。菌根依赖性=(接种植株干质量-不接种植株干质量)/接种植株干质量×100%[26]。
1.3.2 幼苗生长及生理指标测定 对各处理下培养
30 d后的幼苗分别进行生长及生理指标测定。数显游标卡尺测量苗高、地径;将植株地上、地下部分离,万分之一电子天平(赛多利斯BS124S)测量地上与根部鲜重;根系扫描分析系统(WinRHIZO)分析总根投影面积、根表面积、根平均直径和总根体积。收集鲜样用考马斯亮蓝G-250染色法测定根、叶可溶性蛋白含量;乙醇浸提法测定叶绿素含量;根据Tisserant 等[27]人的方法测定根系磷酸酶;用离体法[28]测定根系硝酸还原酶;氯化三苯基四氮唑(TTC)还原法测定根系活力;使用Li-6400型便携式光合作用测定系统(Li-COR Inc,USA)进行光合特性测定。剩余样品植株按根、茎、叶分别装入牛皮纸袋置105 ℃烘箱杀青15 min 后75 ℃烘至恒重以测定各部分干重;烘干后的根、茎、叶干样研磨后,过100 目筛子,淋洗法测定根系阳离子交换量(CEC)。苗木质量指数=苗木总干重(g)/[苗高(cm)/地径(mm)+茎生物量(g)/根生物量(g)][29]。
1.3.3 根际土壤酶活性测定 采用靛酚蓝比色法和苯磷酸二钠比色法测定土壤脲酶和碱性磷酸酶活性以期反映植物对氮、磷的转化及利用;为体现土壤生物学活性,采用3,5-二硝基水杨酸比色法进行蔗糖酶活性测定[30]。
1.4 数据处理
数据分析采用SPSS 22.0 统计软件进行,采用单因素方差分析(One-Way ANOVA)各测定指标,Dun⁃can 法检验数据显著性,Pearson 法进行蒙古沙冬青幼苗根际土壤-植株生长指标间相关性分析。使用Origin 9.0软件作图。
2 结果与分析
2.1 蒙古沙冬青幼苗根系菌根侵染率及菌根依赖性
未接菌处理CK(图1a)根系皮层细胞间或细胞内未有丛枝或无隔菌丝等结构呈现,说明蒙古沙冬青根系中不含有内生AMF。接菌处理后蒙古沙冬青幼苗根系菌根侵染状况如图1a~图1d,供试AMF均可进入根内皮层细胞形成泡囊,不同程度地侵染植株根系(图1b~图1d)。分析比较不同AMF 接种方式下根系菌根侵染率(表1),R.i+F.m 混合接种后蒙古沙冬青植株根系菌根侵染率达61%,显著高于单一接菌。单一接种F.m较R.i植株根系菌根侵染率显著增加。菌根依赖性反映植物和菌根真菌之间的关系,表1可知,蒙古沙冬青幼苗对不同接菌处理后AMF的依赖性不同,R.i+F.m混合接种后植株菌根依赖性(39.8%)显著高于单一接种F.m(32.3%)和R.i(12.8%)。由此说明,AMF 可与蒙古沙冬青形成菌根共生联合体,蒙古沙冬青幼苗根系生长受混合接菌依赖性较单一接菌更强。

图1 丛枝菌根真菌侵染蒙古沙冬青幼苗根系情况Fig.1 Arbuscular Mycorrhizae Fungi infecting root system of Ammopiptanthus mongolicus seedlings

表1 不同AMF接种处理下蒙古沙冬青幼苗菌根侵染率及菌根依赖性Tab.1 Mycorrhizal infection rate and mycorrhizal dependent degree of Ammopiptanthus mongolicus seedlings inoculated with different AMF
2.2 接菌处理对蒙古沙冬青幼苗生长及生物量的影响
图2为接菌处理30 d后蒙古沙冬青幼苗的生长状况,可以看出,混合或单一接菌后幼苗地上部分苗高及地下部分根长较对照均存在差异。表2分析表明,接种AMF后,苗高和根长均显著大于对照,其中,R.i+F.m混合接菌处理的幼苗生长最快,较对照苗高显著增加了47.3%,根长显著增加了72.9%。混合接菌与两种单一接菌处理间也分别存在显著差异(P<0.05)。R.i+F.m混合和单一F.m接菌处理后幼苗地径生长表现一致,均显著大于单一接菌R.i及未接菌对照。总根投影面积、总根体积、地上部鲜重、根冠比、根和叶干重、植株总生物量在接菌后均显著大于对照,其中,R.i+F.m混合接菌的幼苗上述各指标较单一接菌R.i 或F.m 都高且存在显著差异(P<0.05)。幼苗根平均直径和根鲜重在3 种接菌处理间无显著差异,但经R.i+F.m 混合或单一F.m接菌处理的根平均直径和根鲜重显著大于对照。各处理间的根表面积和苗木质量指数存在显著差异,其中,R.i+F.m混合接种最有利于根表面积的扩大(较对照增加147.3%,P<0.05)和苗木质量指数的提高(较对照提高412.5%,P<0.05)。

图2 丛枝菌根真菌侵染蒙古沙冬青幼苗生长情况Fig.2 Growth of Ammopiptanthus mongolicus seedlings infected by AMF
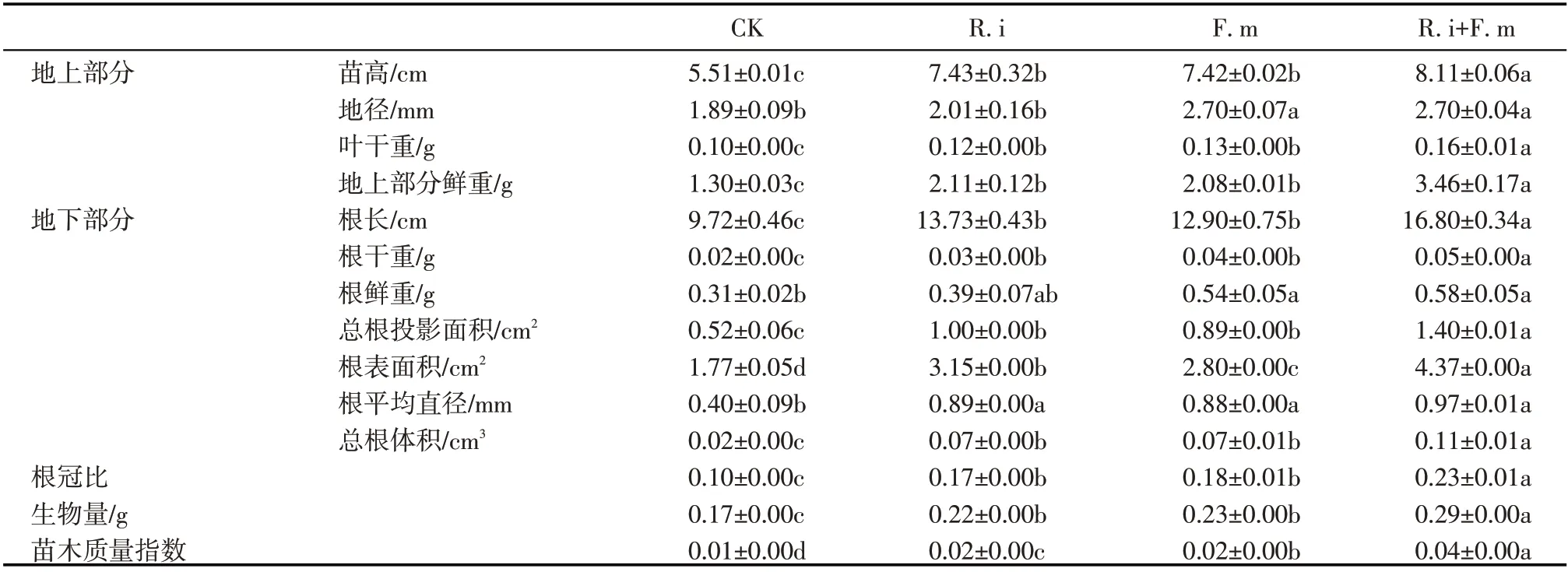
表2 不同丛枝菌根真菌接种处理对蒙古沙冬青幼苗生长指标和生物量的影响Tab.2 Effects of AMF inoculation on growth index and biomass of Ammopiptanthus mongolicus seedlings
2.3 接菌处理对蒙古沙冬青幼苗根系生理特性的影响
2.3.1 根系活力及阳离子交换量 根系活力和阳离子交换量(CEC 值)是衡量根系吸收能力的关键指标,并且间接反映植物生理代谢与生长状况的好坏。由图3可知,不同AMF接菌处理后蒙古沙冬青幼苗根系活力及阳离子交换量(CEC 值)较未接菌对照显著增加,各接菌处理间根系活力及CEC值也存在显著差异。R.i+F.m混合接种最有利于幼苗根系活力的提升(较对照提高了338.7%,P<0.05),单一接种F.m较R.i促进幼苗根系活力的提升效果更为显著。R.i+F.m混合接菌较单一接种R.i或F.m处理后幼苗根系CEC 值分别提高15.4%和25.0%,较对照显著提高177.2%(P<0.05)。单一接种R.i或F.m 的CEC 值较对照处理分别提高140.1%和121.7%(P<0.05)。由此说明接菌处理增强了蒙古沙冬青幼苗根系吸收能力,从而对幼苗地下及地上根、叶各器官生理代谢和生长状况产生间接影响。

图3 接种不同丛枝菌根真菌对蒙古沙冬青幼苗根系活力(a)和根系阳离子交换量(b)的影响Fig.3 Effect of AMF on root activity and cation exchange capacity of Ammopiptanthus mongolicus seedlings
2.3.2 根系可溶性蛋白含量、硝酸还原酶(NR)及碱性磷酸酶(ALP)活性 由图4a 可知,3 种AMF 接菌处理的蒙古沙冬青幼苗根系可溶性蛋白较CK处理含量均高,其中混合接菌处理最能促进根系可溶性蛋白的合成(较对照显著提高240.4%,P<0.05);单一接种F.m 或R.i 较对照分别显著提高161.5%和136.5%,两单一接菌处理间根系可溶性蛋白含量无显著差异。根系可溶性蛋白的变化是幼苗对氮、磷代谢以及氮、磷丰缺程度的响应,与氮、磷吸收密切相关。
进一步分析不同接菌处理后沙冬青幼苗根系氮、磷吸收关键酶活性可知(图4b~图4c),不同接菌处理下幼苗根系硝酸还原酶(NR)活性较对照均显著提高(P<0.05),其中混合接菌处理效果显著优于单一接种处理,而单一接种F.m 显著优于R.i 处理。接种AMF 的幼苗根系碱性磷酸酶(ALP)活性均显著高于对照,不同接菌处理以混合接菌处理效果最优,显著高于单一接种R.i或F.m处理(分别增加了42.1%和12.6%,P<0.05),单一接种R.i 和单一接种F.m 处理较对照分别增加了66.0%、109.4%(P<0.05)。AMF 接菌处理可通过诱导沙冬青幼苗根系硝酸还原酶(NR)及碱性磷酸酶(ALP)活性提升,以促进氮、磷吸收,进而增加根系营养物质(可溶性蛋白)的积累。
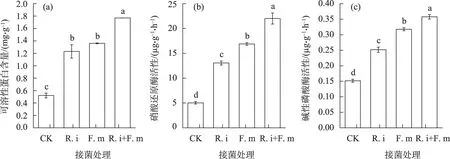
图4 接种不同丛枝菌根真菌对蒙古沙冬青幼苗根系可溶性蛋白、硝酸还原酶(NR)及碱性磷酸酶(ALP)活性的影响Fig.4 Effects of AMF inoculation on soluble protein,activities of NR and ALP in root system of Ammopiptanthus mongolicus seedlings
2.4 接菌处理对蒙古沙冬青幼苗叶片生理特性的影响
2.4.1 叶绿素含量及叶片净光合速率 对光合效能重要指标叶绿素含量(图5a)和叶片净光合速率(图5b)的分析表明,AMF接菌处理可显著提高蒙古沙冬青幼苗叶绿素总含量,单一接种R.i、单一接种F.m和混合接菌处理较对照分别提高47.0%、93.1%、101.1%(P<0.05),单一接种R.i 幼苗叶绿素总含量显著低于混合接种和单一F.m 接菌处理。接种R.i、F.m 及R.i+F.m 混合接菌幼苗净光合速率较未接菌对照分别显著提高198.0%、193.0%、237.5%(P<0.05)。接种AMF,特别是R.i+F.m混合接菌可显著增加蒙古沙冬青幼苗叶绿素合成及叶片光合作用能力,提高幼苗净生产量和有机物积累速率,促进幼苗生长。
图5 接种不同丛枝菌根真菌对蒙古沙冬青幼苗叶绿素总含量、净光合速率及叶片可溶性蛋白的影响Fig.5 Effect of AMF on chlorophyll content,net photosynthetic rate and soluble protein in leaves of Ammopiptanthus mongolicus seedlings
2.4.2 叶片可溶性蛋白含量 图5c 可知,接菌处理后幼苗叶可溶性蛋白含量显著高于未接菌处理。其中R.i+F.m混合接菌处理效果最为显著,较对照增加54.3%(P<0.05),R.i、F.m 单一接菌也显著增加了叶中可溶性蛋白的含量,但其相对增加量(较对照分别增加36.7%和39.5%)低于混合接菌处理。作为表征植物体总代谢的重要指标,沙冬青幼苗叶可溶性蛋白含量的显著增加,结合图4 中根可溶性蛋白的变化结果表明,通过AMF 接菌,促进沙冬青幼苗根系菌根化,不仅可增强沙冬青幼苗根系生理代谢,同时也促使幼苗叶代谢更加旺盛,接种AMF通过影响蒙古沙冬青幼苗生理特性而影响其生长及生物量的形成。
2.5 接菌处理对蒙古沙冬青根际土壤酶活性的影响
图6a~图6c 所示为接种AMF 后蒙古沙冬青幼苗根际土壤脲酶、碱性磷酸酶和蔗糖酶活性变化,单接种R.i或F.m及混合接种R.i+F.m均可显著提升3 种幼苗根际土壤酶活性,且各根际土壤酶活性均以R.i+F.m混合接菌较对照的提升速率最快。3种接菌方式中,R.i+F.m 混合接菌对土壤脲酶活性的激增作用(较对照提高564.7%)显著大于单接种R.i或F.m(较对照增加279.3%和255.0%)。对土壤碱性磷酸酶而言,混合接菌与单接种F.m对其活性的激增效果显著大于单接种R.i。单接种R.i、F.m或混合接种R.i+F.m 均可显著提高蔗糖酶活性达88.6%以上,3种接菌处理间无显著差异。这表明,接种AMF 可诱导宿主植物蒙古沙冬青有效释放土壤酶,从而有利于土壤矿质养分的活化及其有效浓度的提高,这可能成为根系菌根形成及影响幼苗根系生理代谢并促进根系及整个植株生长潜在的原因。

图6 接种不同丛枝菌根真菌对蒙古沙冬青根际土壤酶活性的影响Fig.6 Effect of AMF on enzyme activities in rhizosphere soil of Ammopiptanthus mongolicus
2.6 蒙古沙冬青幼苗根际土壤酶活性与植株生长及生理指标相关性分析
将蒙古沙冬青幼苗生长与根系及叶片生理特性、根际土壤酶活性进行相关分析表明(表3),幼苗总生物量与土壤脲酶和碱性磷酸酶活性呈极显著正相关(P<0.01),与蔗糖酶活性呈显著正相关(P<0.05)。幼苗生长根冠比与根际土壤3 种酶活性均呈极显著正相关。幼苗生长相关生理代谢指标根系活力、根系阳离子交换量、根系硝酸还原酶及碱性磷酸酶活性以及叶净光合速率、叶可溶性蛋白、与其根际土壤各酶活性也呈极显著正相关。对不同AMF 接菌处理蒙古沙冬青幼苗地上与地下部分相关性分析表明,幼苗叶净光合速率与根系活力呈显著正相关(P<0.05),与根系阳离子交换量呈极显著正相关(P<0.01),叶可溶性蛋白与根系活力和根系阳离子交换量均极显著相关。而幼苗生长的根冠比和总生物量与其根系活力及阳离子交换量、叶净光合速率及可溶性蛋白之间也表现为极显著正相关。这表明,幼苗根际土壤酶活性的增加可促进其根系阳离子交换及活力提升,有助于增强根系代谢过程中酶促作用及蛋白等营养物质的积累,同时影响和改善了幼苗地上部分代谢和营养吸收。

表3 蒙古沙冬青根际土壤酶活性与植株生长及生理指标的相关系数Tab.3 Correlation coefficients between root soil enzyme and plant growth of Ammopiptanthus mongolicus
3 讨论
3.1 接种丛枝菌根真菌对蒙古沙冬青幼苗生长的影响
AMF 通过侵染宿主植物并产生菌根效应以影响植物生长。菌根侵染率及菌根依赖性通常可作为衡量植物根系菌根化程度及植物生长对菌根依赖程度的重要指标[31]。AMF侵染率越高,植物根系菌根化程度越高;菌根依赖性越强,意味着菌根效应就越大,AMF对植物生长的影响也越大[32]。本研究中,两种AMF单一接种及混合接种均能够侵染蒙古沙冬青幼苗,其中R.i+F.m 混合接菌幼苗根系侵染率最高,说明混合接菌后沙冬青幼苗的菌根化程度最高。R.i+F.m 处理幼苗菌根依赖性比R.i 和F.m 单一处理分别提高了212.2%和23.3%,进一步表明混合接菌后幼苗生长受菌根化影响也最大。对接菌处理后沙冬青幼苗各生长指标的分析表明,混合接菌对幼苗苗高、地径、根长、根表面积、根总体积,以及根冠比和苗木质量指数的提升最为显著。这与邢红爽等对薰衣草[33]、刘雅辰对荆条幼苗[34]、刘兆娜等[35]对百合生长的研究结果相似。谷文超等[36]在木香试验中也得到了证实,混合接种AMF处理组的菌根效应最佳,木香生长达到最优。不同AMF同时接种相同植株时,被接种植株根系对与其结合的AMF 的选择性更大,而宿主植物对AMF 的选择性决定了菌根生长及其功能的发挥程度[37-38]。本试验两种AMF联合作用于沙冬青幼苗,菌根效应较单一接菌能更加发挥充分且产生互补,两种AMF生态位未重叠的部分为幼苗根系汲取更多养分创造了条件。因此,R.i+F.m 混合接菌能更显著地促进沙冬青幼苗根系生长,并为地上部分生长及植株总生物量积累打下基础。本研究中,与根系生长的比较结果一致,3种接菌处理以R.i+F.m混合接菌后沙冬青幼苗的植株总生物量最大,且显著高于R.i或F.m单一接菌。这与杨应等人[39]针对丛枝菌根促进喀斯特适生植物植株生物量积累的研究结果相似,也一定程度上印证了菌根的生态功能。接种AMF通过增大植物根系表面积和总体积而增加了土壤养分的空间有效性,提高了土壤对植物根系的养分供给。同时,菌根通过活化植物根部,加强了幼苗对营养物质的吸收代谢过程,可有效促进根部有机物积累并向地上部分输出,从而促进植物地上部分的生长和生物量的累积[40-42]。
3.2 接种丛枝菌根真菌对蒙古沙冬青幼苗生理代谢的影响
一般认为,土壤中接种AMF 后,植物根系菌根的形成改善了植株根际微生态环境。植物根系大量氨基酸等有机物质的积累离不开根酶的参与,AMF 通过诱导根际环境中与植物生长相关酶类的合成[43],激发植物根系吸收相关酶活性的提升[44],有助于增强植物根系活力,促进根系阳离子交换量增大。有研究表明,接种AMF 可使莴苣的LsaNHX基因表达上调及PIP2丰度增加,从而提高莴苣的耐盐性,使莴苣植株的生长得到改善[45]。Han 等[46]认为菌根诱导下青杨响应干旱胁迫的14-3-3 基因主要为Pc GRF10 和Pc GRF11,AMF 共生提高了干旱条件下植物体内的生理活性,菌根接种显著提高了青杨根系生物量,促进极度干旱下根系可溶性蛋白含量。本研究中,接种AMF,尤其是R.i+F.m 混合接菌可显著提升蒙古沙冬青幼苗根系活力、根系阳离子交换量、根系碱性磷酸酶及硝酸还原酶活性和可溶性蛋白含量。这表明,AMF接菌处理可诱导改变蒙古沙冬青的根系生理代谢过程,通过促进根系对土壤养分的吸收利用而起到对沙冬青的促生长作用。
有研究表明,AMF可通过改善作物光合参数提高植株叶片光合速率与光合作用能力,进而改善或促进植物生长[47]。说明AMF 对植物的促生长作用不仅仅体现在对根系生长及生理代谢的影响,还直接体现在对植株地上生理代谢过程的影响。本研究中,接种AMF显著提高了蒙古沙冬青幼苗净光合速率、叶绿素含量及叶片可溶性蛋白含量。王志刚等[48]研究表明,接种AMF对矿区植物的净光合速率及生物量均有显著提高。同时,君迁子幼苗植株接种AMF后一定程度上提高了净光合速率,改善其对光的适应性,对植株的生长有显著的促进作用[49]。这可能是由于植物接种AMF 后,菌根形成与AMF自身生长繁殖需要消耗宿主植物部分的光合产物,从而改变了植物碳水化合物的形成和输出,促进了植株净光合速率的提升[50],增加和改善了叶绿素的合成和叶片氮代谢过程,使更多的可溶性蛋白积累于叶片之中。
3.3 接种丛枝菌根真菌对蒙古沙冬青根际土壤酶活性的影响
土壤酶是土壤生化过程的动力,其活性大小可反映土壤肥力、质量、环境的演化情况[51-52]。有研究表明,AMF 可提高植株根际土壤酶活性,如罗方舟等[53]接种AMF 使旱稻根际土壤脲酶及蔗糖酶活性显著提高。任旭琴等[54]及贾红梅等[55]研究结果表明,接种AMF 显著提高淮安红椒连作土壤脲酶、蔗糖酶;丹参根际土壤磷酸酶、脲酶等活性也有所提高。本研究中,接种AMF可提高蒙古沙冬青根际土壤脲酶、碱性磷酸酶和蔗糖酶活性。由于AMF能够诱导植物根内分泌相关酶,并对根际土壤酶产生了刺激和分泌作用,因而有利于土壤酶的活化[56],增加根际土壤酶活性。AMF 通过影响蒙古沙冬青根际土壤酶活性可间接平衡土壤的营养元素水平,由于根际土壤酶活性的提升使得酶促作用加强,从而加速了土壤全氮、全磷向有机氮、有机磷的转化,增强了幼苗根系对土壤中氮、磷素的吸收能力[57]。
4 结论
丛枝菌根真菌(摩西斗管囊霉F.mosseae,根内根孢囊霉R.intraradice)可有效侵染蒙古沙冬青幼苗根系,与其构建互惠共生体。接种AMF对蒙古沙冬青幼苗的促生长作用首先表现为接菌可刺激幼苗根际土壤酶的产生和分泌,显著提高土壤脲酶、碱性磷酸酶和蔗糖酶活性,通过酶促作用促进土壤养分活化和有效浓度的提高,增加土壤肥力。同时,共生AMF 可增加根表面积、根总体积并诱导根系吸收相关酶(硝酸还原酶及碱性磷酸酶)活性的提升和促进根系阳离子交换和根活力增加,有助于增强根系生理代谢过程中营养物质(可溶性蛋白)积累和更大空间对土壤养分的吸收,同时影响和改善了幼苗地上光合代谢及营养存储,从而促进沙冬青根系及幼苗生长和生物量的累积。研究结果对于解决蒙古沙冬青幼苗根系不发达、再生性差、移栽造林成活率低的难题具有重要意义,并可为蒙古沙冬青菌根化造林技术研发提供理论依据。
猜你喜欢
中国土壤与肥料(2021年5期)2021-12-02 01:06:26
中华建设(2020年5期)2020-07-24 08:55:14
园林科技(2020年2期)2020-01-18 03:28:26
小学生必读(低年级版)(2018年6期)2018-09-08 07:44:36
天然产物研究与开发(2018年3期)2018-05-07 06:38:35
——致坚守奋斗的你
音乐天地(音乐创作版)(2017年1期)2017-02-12 18:58:38
中国蔬菜(2016年8期)2017-01-15 14:23:38
中央民族大学学报(自然科学版)(2015年3期)2015-06-11 02:13:46
环境与可持续发展(2013年6期)2013-03-11 16:21:51
植物营养与肥料学报(2011年6期)2011-10-24 05:53:04
