
马仑亚高山草甸土壤可培养微生物数量特征
2023-03-02 03:17:40张宝贵刘晓娇徐满厚常丽娟吴佳灵冯煜清赵慧茹
太原师范学院学报(自然科学版) 2023年4期
张宝贵,刘晓娇,朱 茜,徐满厚,常丽娟,吴佳灵,冯煜清,赵慧茹
(太原师范学院 地理科学学院,山西 晋中 030619)
0 引言
微生物存在于各类极端环境中,如沙漠[1]、冰川[2]、冻土[3]、海底热液[4]等,并在以上各类环境中扮演着极其重要的角色.作为土壤生物的重要组成部分,土壤中微生物数量也十分庞大[5].虽然在土壤有机质中土壤微生物生物量所占比例很小,但土壤微生物却是土壤中最活跃的组成部分,它们在养分有效性和维持土壤质量方面发挥着至关重要的作用[6].研究土壤微生物及其影响因素对于预测生态系统对环境变化的响应以及改善各种生态系统的变化至关重要[7].因此,有必要研究各类生境土壤微生物数量及群落如何受环境变化或干扰的影响.
气候变暖已经成为不争的事实,自1850年以来,全球地表平均温度已上升约1 ℃,且在未来几十年里,所有地区的气候变化都将继续加剧[8].高山草甸是一种特定的气候产物,对气候变化极其敏感,该生态系统中土壤微生物数量引起了众多学者的广泛关注,如相关研究发现高寒草甸土壤微生物数量对增温的响应随着海拔的升高而变化,且与土壤温度显著正相关[9-10];也有研究表明青藏高原东端高寒草甸退化过程中土壤三大类微生物数量及微生物总数均呈现先增加后减少的趋势[11];类似地,笔者对青藏高原东北边缘过渡带研究发现翻耕补播措施并没有恢复该区域高寒草甸土壤微生物数量[12];也有研究表明海拔梯度通过影响土壤温度、植被类型、物种数和土壤养分等环境因子来改变高山草甸土壤微生物数量[13].
与高山草甸一样,亚高山草甸也是一种特定的气候产物,不同研究表明不同区域亚高山草甸土壤微生物数量或多样性具有不同特点.如滇西北地区植物种群扩散使该区域亚高山草甸土壤细菌、放线菌、真菌数量均显著增加[14];而人类干扰条件下该地区亚高山草甸只有土壤真菌数量显著增加,其他类型微生物数量均显著减少[15].类似地,草地利用强度的增加使得祁连山亚高山草甸土壤微生物总DNA量呈减少趋势[16].在青藏高原东部弃耕地恢复演替过程中亚高山草甸土壤微生物多样性则得到了较好的恢复[17].相关分析表明,川西北亚高山草甸土壤真菌多样性受雪被厚度的显著影响[18-19],而土壤深度则是若尔盖亚高山草甸土壤微生物数量变化的主要影响因子[20].
马仑亚高山草甸地处山西省吕梁山脉北部地区,该地区亚高山草甸分布于海拔2500 m以上,是华北地区最典型的亚高山草甸之一,属于全球气候变化的敏感区[21].已有学者对该地区植物进行了研究[22-23],而关于该地区土壤微生物的研究还未见报道.基于此,本研究以该地区不同海拔、不同深度亚高山草甸土壤为研究对象,借助可培养方法系统研究该区域土壤三大类微生物数量特征,并结合环境因子分析影响该地区微生物数量的关键因子.研究结果不仅可以填补该区域微生物研究的空白,同时可为区域亚高山草甸土壤微生物响应气候变暖提供理论依据与数据支撑.
1 研究区概况与研究方法
1.1 研究区概况
马仑亚高山草甸地处晋西北黄土高原,位于山西省吕梁山北段宁武县境内的管涔山顶(112°36′~112°37′ E、38°57′~39°03′ N),最高海拔2721.7 m,面积大约230 hm2.该区年平均气温6.2 ℃,最热月为7月,平均气温20.1 ℃.年均降水量475 mm,属于温带大陆性季风气候;土壤类型主要为亚高山草甸土;主要植被类型为典型的亚高山草甸[23].
1.2 样地设计与样品采集
前期对该区域进行实地考察后,如表1所示,于2019年9月在该区域亚高山草甸的分布范围内设置采样点,海拔分别为2600 m(MLL)、2650 m(MLM)和2700 m(MLT),每个采样点均在地势平坦处设置三个平行样地,每个样地均采集0~10 cm和20~30 cm两个不同深度的土壤,包括微生物样品和土壤理化性质样品,具体采集方法见参考文献[5].

表1 采样点信息表
1.3 三大类土壤微生物的分离培养
本研究采用稀释涂布平板法对三大类土壤微生物进行分离培养,其中采用富营养培养基牛肉膏蛋白胨对细菌进行分离培养,采用改良高氏一号培养基对放线菌进行分离培养,真菌的分离培养则采用马丁氏孟加拉红培养基.在正式涂布之前,对每类微生物的最适合涂布浓度进行预实验以确保计数的科学性与准确性.最终确定10-6为细菌生长的最佳浓度,10-5为放线菌生长的最佳浓度,10-2为真菌生长的最佳浓度.之后对各类微生物进行涂布,具体操作步骤见参考文献[5].涂布后于25 ℃恒温培养箱对细菌进行培养并在培养7 d后计数,放线菌置于28 ℃恒温培养箱培养8 d后计数,真菌置于28 ℃恒温培养箱培养10 d后计数.
1.4 土壤理化性质的测定
采用雷磁电导率仪 FE38-Standard测定土壤总溶解性固体(TDS)、电导率(EC)与盐度(SS);采用METTLER TOLEDO FE28-Standard酸度计测定土壤pH值;采用烘干法(105 ℃,24 h)测定土壤含水量(SWC);采用德国耶拿C/N 分析仪2100测定土壤全碳(STC);采用土壤温度传感器(S-TMB-M006)测定土壤温度(ST).
1.5 数据处理与分析
所有数据均采用Excel 2016进行处理并利用SPSS 19.0统计分析软件进行单因素方差分析和相关性分析;用Origin 9.0进行绘图.
2 结果与分析
2.1 不同海拔、不同深度土壤理化性质变化特征
表2为马仑亚高山草甸不同海拔、不同深度土壤理化性质特征.从表中可知,EC介于50.89~152.88 μS·cm-1之间,STC含量介于26.9~57.63 g·kg-1之间,且同一海拔20~30 cm STC含量显著低于0~10 cm(P<0.05);而随着海拔高度的增加同一深度STC含量则呈显著上升趋势(P<0.05).ST介于6.45~10.82 ℃之间,SWC 介于40.58~54.74 %之间,且变化趋势与STC一致.TDS含量介于25.44~49.91 mg·L-1之间,其中海拔2700 m处20~30 cm TDS含量高于0~10 cm,其余两个海拔则表现为0~10 cm TDS含量高于20~30 cm;此外,随着海拔高度增加,同一深度TDS含量呈上升趋势,但没有达到显著水平.土壤pH值总体呈弱碱性,变化范围介于7.48~8.55之间;SS变化范围介于0.01~0.06 psu之间.两者与STC的变化趋势相同,但均未达到显著水平.

表2 不同海拔梯度亚高山草甸土壤理化性质
2.2 不同海拔、不同深度土壤可培养细菌数量变化特征
如图1所示,研究区域可培养细菌数量介于2.13×109~16.53×109CFU·g-1之间,其中MLM1样点20~30 cm土壤细菌数量最少,为2.13×109CFU·g-1,而MLL1样点 0~10 cm土壤可培养细菌数量最多,为16.53×109CFU·g-1,高一个数量级.此外,由图1可知,每个海拔下的三个样地均表现为0~10 cm土壤细菌数量显著高于20~30 cm土壤细菌数量(P<0.05),表明可培养细菌数量受土壤深度的显著影响.
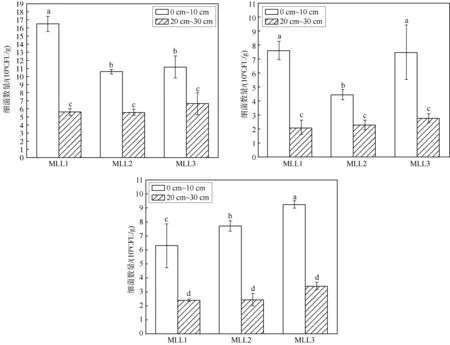
图1 马仑亚高山草甸不同海拔、不同深度土壤细菌数量特征
此外,图2表明可培养细菌数量受海拔影响显著.具体表现为不同海拔、不同深度土壤细菌数量均随海拔高度的升高而显著下降(P<0.05),MLM与MLT样点土壤细菌数量显著低于MLL样点(P<0.05)、但MLM与MLT样点之间土壤细菌数量未达到显著性差异.

图2 马仑亚高山草甸不同海拔、同一深度土壤细菌数量变化特征
2.3 不同海拔、不同深度土壤可培养放线菌数量变化特征
如图3所示,研究区域可培养放线菌数量介于5.6×107~1.56×109CFU·g-1之间,其中MLM3样点0~10 cm土壤放线菌数量最少,为5.6×107CFU·g-1,而MLL3样点0~10 cm土壤可培养细菌数量最多,高达1.56×109CFU·g-1,高两个数量级.此外,由图3可知,MLL样点的三个样地均表现为0~10 cm土壤放线菌数量显著高于20~30 cm土壤放线菌数量(P<0.05);而MLT样点的三个样地变化趋势恰恰相反,均表现为0~10 cm土壤放线菌数量显著低于20~30 cm土壤放线菌数量(P<0.05).MLM样点的MLM1样地0~10 cm土壤放线菌数量显著高于20~30 cm土壤放线菌数量(P<0.05);MLM3样地则恰恰相反,0~10 cm土壤放线菌数量显著低于20~30 cm土壤放线菌数量(P<0.05);MLM2样地不同深度土壤放线菌数量之间不存在显著性差异.与细菌一样,以上结果表明可培养放线菌数量同样受土壤深度的显著影响.

图3 马仑亚高山草甸不同海拔、不同深度土壤放线菌数量特征
此外,图4表明可培养放线菌数量受海拔影响显著.不同海拔、不同深度土壤放线菌数量均随海拔高度升高呈先下降后上升的显著趋势(P<0.05).具体表现为0~10 cm MLL样点土壤放线菌数量显著高于MLT样点(P<0.05),而20~30 cm MLL样点土壤放线菌数量则显著低于MLT样点(P<0.05).

图4 马仑亚高山草甸不同海拔、同一深度土壤放线菌数量变化特征
2.4 不同海拔、不同深度土壤可培养真菌数量变化特征
如图5所示,研究区域可培养真菌数量介于6×103~8.42×105CFU·g-1之间,其中MLL1样点20~30 cm土壤真菌数量最少,仅为6×103CFU·g-1,而MLT1样点 0~10 cm土壤真菌数量最高,高达8.42×105CFU·g-1,高两个数量级.此外,由图5可知,除MLL1、MLL3两个样地外,其余样地均表现为0~10 cm土壤真菌数量显著高于20~30 cm土壤真菌数量(P<0.05),表明可培养真菌数量同样受土壤深度的显著影响.

图5 马仑亚高山草甸不同海拔、不同深度土壤真菌数量特征
此外,图6表明可培养真菌数量受海拔影响显著.不同海拔、不同深度土壤真菌数量均随海拔高度的升高而显著上升(P<0.05),具体表现为MLT样点土壤真菌数量显著高于MLM与MLL样点(P<0.05).
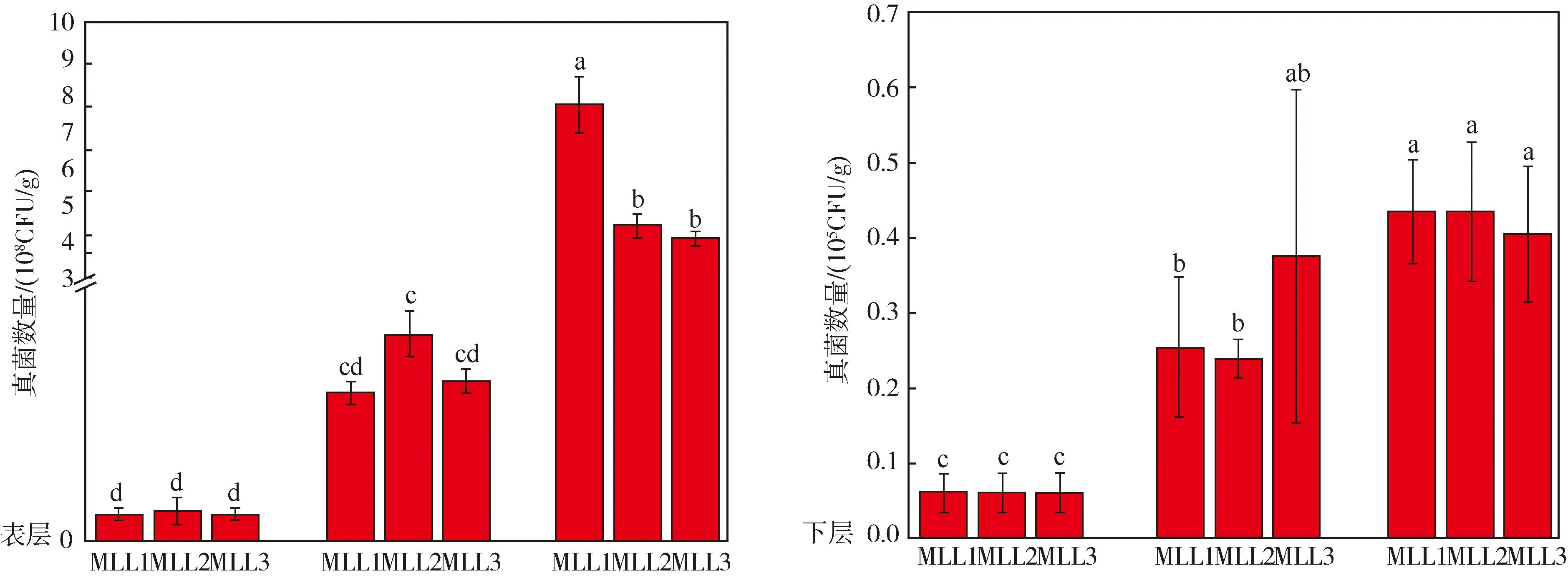
图6 马仑亚高山草甸不同海拔、同一深度土壤真菌数量变化特征
2.5 环境因子与土壤微生物数量的相关分析
由表3可知,可培养细菌数量与ST显著正相关(P<0.05);真菌数量与土壤pH值极显著正相关(P<0.01);放线菌数量与TDS含量极显著正相关(P<0.01),与ST显著正相关(P<0.05).

表3 环境因子与可培养微生物数量相关性分析
3 讨论
3.1 可培养微生物数量变化特征
马仑亚高山草甸不同海拔、不同深度土壤可培养细菌数量最多,介于2.13×109~1.65×1010CFU·g-1之间,且明显高于其他相似高寒草甸生态系统.如甘南高寒草甸不同海拔、不同根系细菌数量介于2.06×107~3.30×108CFU·g-1之间[24];青藏高原北麓河不同类型高寒草地生态系统细菌数量介于0.40×107~4.60×107CFU·g-1之间[25];祁连山中段黑河上游不同海拔细菌数量介于4.60×107~3.70×108CFU·g-1之间[26];天祝高寒草甸不同干扰生境下的细菌数量介于1.95×106~7.31×106CFU·g-1之间[27];而在若尔盖亚高原区,细菌数量仅介于1.40×105~2.85×105CFU·g-1之间[20].马仑亚高山草甸不同海拔、不同深度土壤可培养放线菌数量次之,介于5.6×107~15.63×108CFU·g-1之间,与细菌一样,也明显高于其他相似高寒草甸生态系统.如三江源地区不同退化程度高寒草地土壤放线菌数量介于6.68×104~19.88×104CFU·g-1之间[28];青藏高原海北高寒草甸生态系统土壤放线菌数量介于1.23×104~8.22×104CFU·g-1之间[29];西藏地区土壤放线菌数量仅介于0.1×103~5.22×105CFU·g-1之间[30].马仑亚高山草甸不同海拔、不同深度土壤可培养真菌数量最少,介于6×103~8.42×105CFU·g-1之间,同样高于相似的高寒草甸生态系统.如三江源区不同类型高寒草甸土壤真菌数量介于1.2×103~4.3×103CFU·g-1之间[31];天祝高寒草原不同生境土壤真菌数量介于2.35×104~4.55×104CFU·g-1之间[27].究其原因,可能是由于青藏高原高寒草甸生态系统所处海拔更高,环境更恶劣,土壤在长期低温胁迫下容易冻结,通透力差,从而对土壤微生物的生长繁殖具有较强的抑制作用[3].而马仑亚高山草甸地处华北地区,相对于高寒冷环境而言,土壤环境较好,有利于土壤微生物的大量生长繁殖.
3.2 可培养微生物数量与土壤深度、海拔高度关系
本研究发现土壤深度对可培养微生物数量影响显著.在本研究中,同一海拔高度土壤可培养细菌和真菌数量均表现为0~10 cm显著高于20~30 cm,与相关学者在青藏高原、东北黑土地以及内蒙古荒漠草原的研究结论一致[32-34].可培养放线菌数量变化特征略有不同.由此可见,不同类型土壤微生物随着土壤深度的变化表现出不同的变化趋势.究其原因,可能是由于0~10 cm 土壤有机质含量较高,能提供微生物生长发育所需的各类营养元素,因而更适合土壤微生物的生长发育.因此0~10 cm土壤细菌和真菌数量高于20~30 cm,而放线菌则更能适应相对贫瘠的环境[35-36].
其次,本研究发现0~10 cm和20~30 cm两个深度不同类型土壤微生物数量随海拔高度的升高变化趋势也不同.具体表现为细菌数量随海拔高度的升高均呈下降的趋势.与本研究结果一致,张丹丹等在珠穆朗玛峰不同海拔梯度上同样发现土壤细菌丰度随海拔增加而降低,且与海拔高度存在显著负相关关系[37].但是在疏勒河上游土壤可培养细菌数量随着海拔高度的升高却呈现先下降后上升的趋势[38].本研究放线菌数量随海拔高度的升高呈现先下降后上升的变化趋势,而在甘南高寒草甸区及青海高寒草甸区,相关学者发现随着海拔高度的升高放线菌数量呈现下降的趋势[29,39],表明微生物的环境异质性很强.本研究真菌数量随海拔高度的升高呈上升趋势,但是在色季拉山研究者发现海拔越高,真菌数量越少[40].综上所述,不同区域、不同类型、不同深度土壤微生物数量随海拔高度变化趋势不同,可能是由于地理环境的区域差异性,造成不同区域的地理环境因子不同,这些环境因子通过影响植被特征与土壤特征进而影响土壤微生物数量[5].
3.3 可培养微生物与环境因子的关系
本研究表明,制约不同海拔、不同类型土壤微生物数量的关键环境因子不同.细菌数量受ST显著影响.在南方红壤丘陵区,土壤细菌也受到ST的显著影响[41].类似地,在东北寒温带落叶松天然林区域,气候变化导致的温度变化对土壤细菌群落存在显著影响[42].相关学者在江西官山不同海拔梯度的研究同样表明ST对细菌影响显著[43],可能是由于高海拔地区低温胁迫造成的[44].作为仅次于细菌的第二大类微生物,放线菌也是生态环境中一类非常重要的微生物群类.本研究中TDS与ST是影响放线菌数量的主要环境因子.类似地,李博文[39]在甘南高寒草甸区发现ST是限制放线菌生长的关键环境因子.也有研究表明该类型微生物数量往往受到土壤pH值、温度、湿度、海拔和区域小气候等局部因素的共同影响[45-47].本研究中土壤真菌数量相对较少,且受土壤pH值影响显著,与前人研究结果一致[48-49].此外,Liu等[50]研究表明pH值也是影响中国西南地区不同海拔土壤真菌多样性的重要环境因子之一,且土壤pH值越高,土壤真菌多样性指数越高.类似地,相关学者研究表明长白山不同海拔土壤真菌的丰富度与土壤pH值有很强的相关性[45].当然,也有学者研究表明土壤真菌数量及多样性主要受土壤含水量[51]、土壤温度[52]等环境因子的影响.综上所述,土壤微生物与环境因子之间的关系十分复杂,不同区域之间具有区域差异性,同一区域不同类型微生物之间也有差异性,有待进一步深入研究.
4 结论
研究区不同海拔、不同深度可培养细菌数量最多,介于2.13×109~16.53×109CFU·g-1之间;放线菌次之,介于5.6×107~15.63×108CFU·g-1之间;真菌最少,介于6×103~8.42×105CFU·g-1之间.
不同海拔、不同深度土壤可培养微生物数量明显不同.具体表现为同一海拔0~10 cm土壤细菌、真菌数量均显著高于20~30 cm(P<0.05),放线菌略有不同.不同海拔0~10 cm与20~30 cm土壤细菌数量均表现为随着海拔高度的升高而显著下降(P<0.05).土壤放线菌数量均表现为随着海拔高度的升高呈现先下降后上升的显著趋势(P<0.05).而土壤真菌数量均表现为随着海拔高度的升高而显著上升(P<0.05).
限制不同类型土壤微生物数量的关键环境因子不同.细菌数量主要受ST影响;放线菌数量主要受TDS影响;真菌数量则主要受土壤pH值影响.
猜你喜欢
儿童故事画报·自然探秘(2024年5期)2024-05-22 22:26:58
疯狂英语·新读写(2023年4期)2023-05-10 10:44:22
青海草业(2022年2期)2022-07-23 09:34:58
国际呼吸杂志(2019年22期)2019-12-09 09:20:36
活力(2019年21期)2019-04-01 12:17:10
绿色科技(2017年6期)2017-04-20 08:13:04
河北林业科技(2016年5期)2016-11-08 03:13:30
西南农业学报(2016年5期)2016-05-17 05:42:44
西藏大学学报(自然科学版)(2015年1期)2015-12-25 11:43:44
应用海洋学学报(2014年4期)2014-11-22 07:43:54
