
运动锻炼调节miRNA对多种类型癌症的改善及机制的研究进展
2023-02-21 13:18:08上官若男张晓波孙景权
中国体育科技 2023年1期
陈 敏,上官若男,张晓波,孙景权*
世界卫生组织国际癌症研究机构调查数据显示,2020年全球新增癌症人数约1 929万人,中国新增癌症患者占比23.7%,成为癌症新增人数最多的国家(Sung et al.,2021)。国家癌症中心发布的2022年全国癌症统计数据显示,2016年恶性肿瘤发病约406.4万人,死亡约241.4万人,平均每天超过1.1万人被确诊(Zheng et al.,2022)。癌症是一种局部病变的全身性疾病,造成局部组织的发育衰竭并形成肿瘤。畸变的蛋白分子通过恶性循环进行大量增殖复制,造成基因突变并形成致癌物,致癌物随即阻碍干细胞分化,导致机体局部生物信号细胞增殖和分化的紊乱,最终促进其发展(Lai,2019)。致癌物的增殖和扩散速度极快,现代医药的作用有限,迫切需要发现和开发新的诊疗方法。microRNA(miRNA)是多种癌症诊断和预后标志物,其异常表达是值得关注的话题。miRNA是一种小型非编码RNA,调节基因转录后的表达。在过去10年里,miRNA作为肿瘤抑制因子或致癌基因引起广泛关注,其与肿瘤的发展、转移和各种癌症疗法的内在抗性有关(Jiang et al.,2018;Zhang et al.,2019a),在癌症中主要作为促癌因子和抑癌因子发挥作用。一般情况下,癌基因miR‑NA在肿瘤样本中上调,而抑癌基因miRNA下调。实际上,现在已知的miRNA能够介导许多与癌症进展、转移(沈克,2014;阳德全 等,2020)和治疗耐药性(吕腾,2019)有关的生物学途径,提示其可能是优化癌症诊疗的工具。
miRNA在健康个体中一直处于稳定状态,但包括生活方式在内的外部因素能够影响其表达(Gomes et al.,2014)。越来越多的研究表明,癌症中抑癌miRNA的表达明显受限(谷士海 等,2020;王书廷 等,2020;Wang et al.,2019),而运动锻炼作为调节miRNA表达的有效途径,能够明显降低癌症风险和减缓癌症进程(Mooren et al.,2014)。目前,关于运动锻炼调节miRNA改善的癌症类型及其背后的可能机制尚未完全明晰。本研究分析运动锻炼诱导的相关miRNA变化与癌症相关的miRNA表达之间的潜在联系,总结运动锻炼诱导miRNA可能改善的癌症类型,并梳理运动锻炼诱导miRNA改善相关癌症的信号通路。
1 运动锻炼降低癌症的风险和发展
运动锻炼能够减少癌症的发生,并抑制肿瘤生长。流行病学研究认为,全球25%的癌症发生与超重和静态生活方式有关,而运动锻炼则能够通过多种途径降低癌症的患病风险(张雪 等,2019),包括运动介导的代谢激素水平(Maddocks et al.,2013)、炎症反应(Hojman,2017)、免疫细胞功能(Idorn et al.,2017)变化以及miRNA水平调节(Gomes et al.,2014)等。研究表明,运动锻炼对癌症及其治疗有直接影响,是预防癌症的潜在干预手段(Chris‑tensen et al.,2018)。运动锻炼与乳腺癌、结直肠癌以及前列腺癌的发病率之间存在显著负相关(Ashcraft et al.,2016)。在小鼠模型中,自主运动能够导致小鼠肿瘤的发生率和生长率降低60%以上(Idorn et al.,2017)。此外,临床研究表明,随着运动时间的推移,体内会发生一系列与癌症相关的生物学及表观遗传学的变化(Thomas et al.,2017)。对26名久坐不动的中年男性进行12周的耐力和力量综合训练后,其骨骼肌中191个基因存在差异表达(Pourteymour et al.,2017);而由运动锻炼诱导的抑癌基因的差异表达,能够有效抑制肿瘤细胞的侵袭、转移及自我更新等(Isanejad et al.,2016);而运动锻炼作为一种预防和改善癌症的干预手段,在临床上得到越来越多的应用。
2 运动锻炼诱导miRNA出现适应性变化
miRNA广泛存在于人体的组织和器官中,并以非常稳定的形式释放到外周血液循环(Redova et al.,2013),其含量因运动训练刺激而改变。因此,miRNA作为一种对运动锻炼有刺激性反应的潜在生物标志物受到关注。2013年1月至2020年1月,在中国知网、PubMed、Science Direct等数据库中,以“exercise”“miRNA”“cancer”“mecha‑nism”为关键词检索相关文献。其中,检索到关于运动锻炼影响健康个体血液中miRNA含量变化(至少4周运动干预)的研究文献共9篇(表1)。关于运动锻炼引起miRNA含量变化的研究尚处于初步阶段,相关研究数量有限,且大部分研究的样本量较小,因此在筛选文献时未设置最小样本阈值。人体血液中miRNA含量受相关疾病影响,因此,为排除运动以外因素对miRNA含量的影响,所纳入的9项研究的受试者均为健康个体。
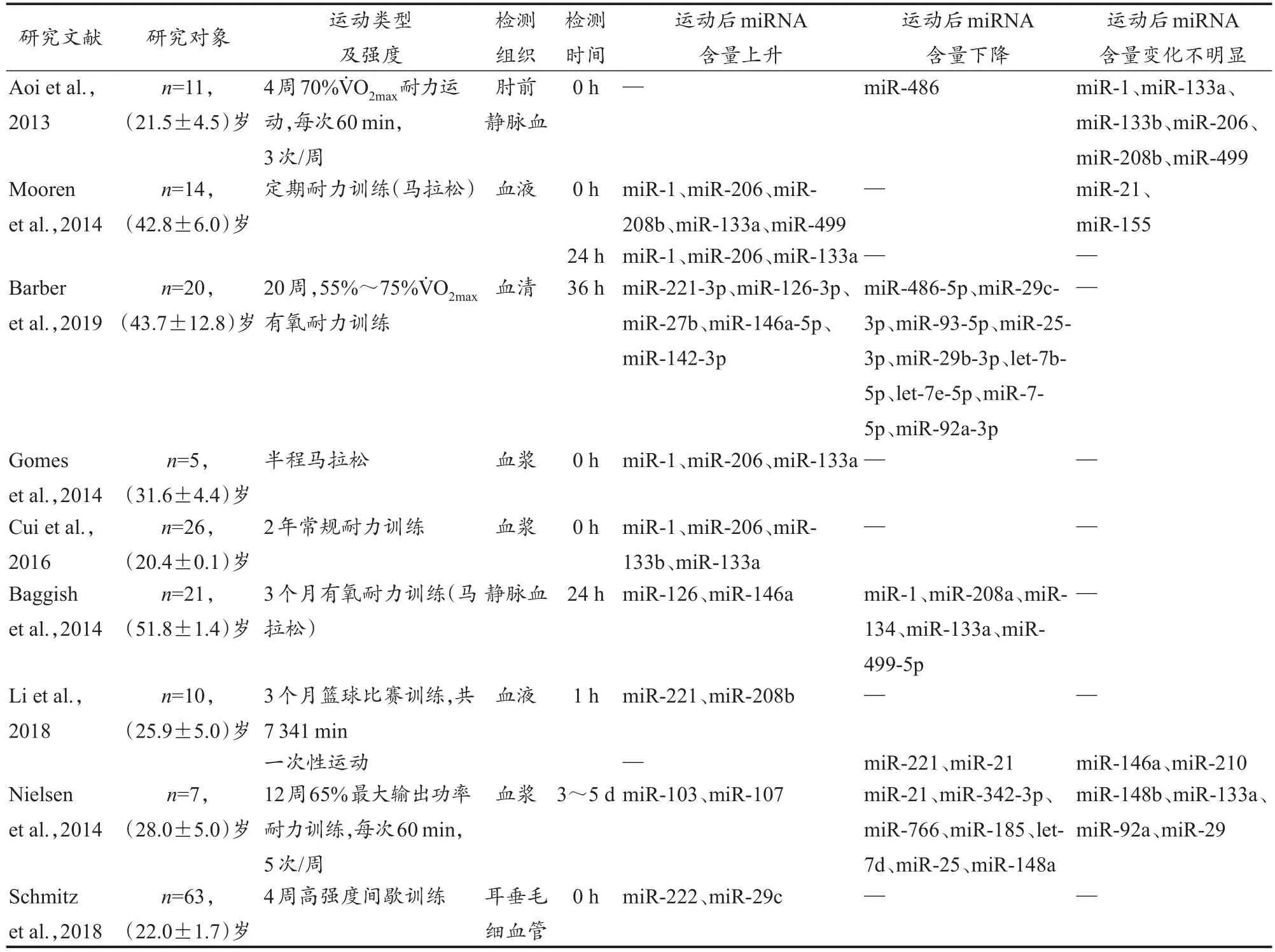
表1 运动锻炼诱导miRNA的适应性变化Table1 Adaptive Changes of Exercise-Induced miRNA
综合相关研究发现,经过长期运动锻炼后,血浆中miR-1、miR-133a和miR-206的含量明显上升,且这种变化可持续至运动后 24 h(Cui et al.,2016;Gomes et al.,2014;Mooren et al.,2014);而 miR-146a和 miR-126在血浆中含量上升的状态可持续36 h甚至更久(Barber et al.,2019)。Nielsen等(2014)研究表明,12周耐力运动后3~5 d仍可以检测出部分miRNA的含量变化。有研究表明,经过长期运动锻炼后,miR-486在血浆中的含量呈下降趋势(Aoi et al.,2013;Barber et al.,2019)。总体而言,长期的运动锻炼会导致血浆中miRNA的含量变化,且运动锻炼对miRNA的影响具有一定时效性。
3 运动锻炼可能通过调节miRNA改善多种类型癌症
3.1 miRNA参与癌症发生和发展
miRNA是一类具有调控功能的内源性非编码RNA,大小约20~25个核苷酸,通过对调节mRNA转录的基因进行切割、失稳或抑制,进而调控相关蛋白表达。据预测,miRNA能够调控55%以上的蛋白质编码基因(Friedman et al.,2009),并参与细胞发育、增殖、新陈代谢和信号转导等过程。miRNA与mRNA只有部分碱基互补,因此单个miRNA可能靶向多个不同的mRNA(Zhu et al.,2018)。同样,单个mRNA可能包含多个不同的miRNA结合位点,从而形成复杂的调控网络(Zhu et al.,2018)。通过这种作用模式,miRNA直接或间接调控癌症相关的mRNA,进而广泛参与癌症的发生和发展(Yuan et al.,2016)。在实体和血液肿瘤中,已发现大量miRNA含量发生改变,并且其表达与诊断时癌症的严重程度和分期有关(Tutar et al.,2015)。
相关研究表明,miRNA对癌症的发生和进展具有双重效用,其靶点是肿瘤抑制因子或癌基因(Priyadarshini et al.,2013)。因此,特定miRNA的表达能够下调癌基因或重新表达肿瘤抑制基因,进而形成肿瘤抑制(Song et al.,2016)。如在肝癌中,miR-206通过靶向c-Met基因的3’-UTR导致c-Met基因沉默,从而阻断CDK6的细胞周期进程,预防肝癌发生和发展(Umeh-Garcia et al.,2018)。miRNA是癌症发展中的诊断标记物,如在乳腺癌中,已经发现miR-1、miR-133a、miR-126等是早期癌症发生的生物标记物(Muti et al.,2014)。在癌症患者中,miRNA失调将会影响癌细胞增殖、分化和侵袭等(Park et al.,2018)。基于此,调控miRNA对癌症患者的康复极为重要。
3.2 运动锻炼诱导的miRNA可能改善多种类型癌症
运动锻炼能够诱导基因的差异表达(Ornish et al.,2008),其中就包括miRNA,且不同运动类型对miRNA的含量产生不同影响。近年来,运动锻炼诱导miRNA的含量变化引起广大学者的关注。相关研究发现,12周高强度间歇训练后,乳腺癌患者血清中抑癌miRNA含量明显增加,促癌miRNA含量明显减少(Isanejad et al.,2016)。同时,运动锻炼对miRNA的调节能够抑制相关促癌因子,并有效抑制肿瘤细胞自我更新、侵袭和转移等,降低罹患癌症的风险(Mooren et al.,2014)。可见,运动锻炼可能通过调节miRNA的含量变化,进而影响癌症的发生与发展。
目前,越来越多的研究表明,长期运动锻炼可诱导相关miRNA的含量变化,且这些miRNA作为抑癌基因在改善癌症相关信号通路中发挥重要作用(Alhasan,2019;Cui et al.,2013;Wang et al.,2019)。由此推测,运动锻炼可能通过诱导miRNA含量变化改善相关癌症。虽然长期运动锻炼后miR-486含量下降,但有研究表明,较低含量的miR-486表达并不会增加癌症不良预后的风险(Jiang et al.,2018)。因此,结合表1的分析结果,将运动锻炼诱导miRNA改善的癌症类型进行汇总(图1),有6种癌症能够同时被运动诱导的miRNA改善。可见,运动锻炼通过调节在多种类型癌症中发挥抑癌因子作用的miRNA(miR-1、miR-126、miR-133a、miR-146a、miR-206)含量,可能对结直肠癌、肺癌、前列腺癌、乳腺癌、肝癌和胃癌具有良好的改善作用。
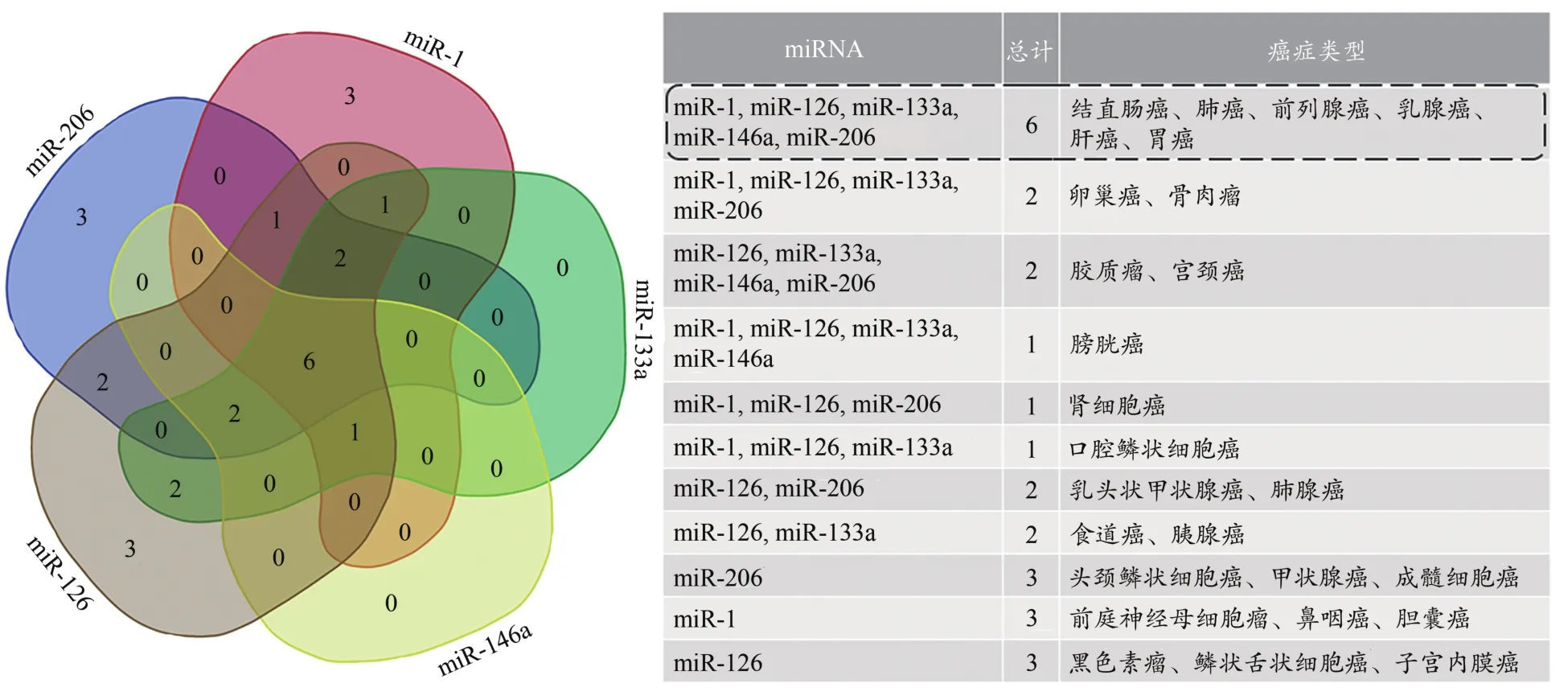
图1 长期运动锻炼诱导miRNA改善的癌症类型Figure 1. Cancer Types of Long-Term Exercise Improved by Regulating miRNA
4 运动锻炼调节miRNA改善癌症的可能机制
miRNA已被证实能够抑制癌症相关重要基因的表达(Cui et al.,2013),能够调控大约30%人类癌基因和肿瘤抑制基因(Areeb et al.,2015)。miRNA调控靶基因的主要机制是通过与mRNA 3’非翻译区(3’-untranslated region,3’-UTR)部分碱基互补序列相结合,诱导mRNA降解或抑制mRNA翻译(Zhu et al.,2018),阻滞细胞周期进程,进而有效靶向肿瘤干细胞并阻碍其转移。如miR-206针对c-Met基因的 3’-UTR进行沉默(Wang et al.,2019),进而将细胞周期阻滞在G0/G1期(Zhou et al.,2019);miR-126的表达与细胞增殖减少和癌细胞在G1期阻滞有关(Al‑hasan,2019)。运动锻炼能够调节相关miRNA含量,从而达到改善癌症的效果,而miRNA改善癌症主要通过调控相关因子。鉴于此,在分子水平上了解运动锻炼诱导miR‑NA干预癌细胞的相关信号通路十分重要,既有利于了解运动锻炼通过调节miRNA改善癌症的机制,亦有助于将运动锻炼应用于癌症的临床控制。
4.1 运动锻炼通过调节miRNA改善乳腺癌的可能机制
乳腺癌是全世界女性最常见的恶性肿瘤之一。在乳腺癌相关研究中,miR-206、miR-133a、miR-126、miR-1和miR-146a作为抑癌因子在组织中的表达显著降低(Al‑hasan,2019;Cui et al.,2013;Long et al.,2019),而运动锻炼能够显著增加miR-1、miR-126、miR-133a、miR-146a、miR-206的含量。事实上,经过一段时间的运动锻炼后,miR-206在乳腺肿瘤组织中表达增加,并减少肿瘤组织生长(Isanejad et al.,2016)。癌症干细胞作为能够自我更新并介导肿瘤生成和转移的癌细胞亚群,在肿瘤生成和转移中的作用至关重要。有研究发现,在乳腺癌干细胞(breast cancer stem cells,BCSCs)中,T-Box转录因子 3(T-Box tran‑scription factor 3,TBX3)过表达,参与其增殖、迁移和调控(Amir et al.,2016),并调节TGF-β1的促迁移能力,这是BCSCs扩增的必要和充分条件(Li et al.,2013)。然而,TBX3在乳腺癌细胞中受运动锻炼调节的miR-206抑制,且这种抑制能够消除肿瘤侵袭表型(Amir et al.,2016)。其主要机理是,运动锻炼调节的miR-206抑制TBX3转录因子,降低TGF-β促迁移能力,进而抑制BCSCs迁移。同理,受运动锻炼调节的miR-1通过与卷曲蛋白7(frizzled class receptor 7,FZD7)结合,阻滞Wnt/β-catenin信号转导,进而抑制BCSCs增殖和迁移(Liu et al.,2015)。
在肿瘤细胞中,过表达的miR-133a使细胞周期阻滞在G2/S期,抑制DNA新合成(Cui et al.,2013)。而有研究发现,miR-133a含量受运动锻炼的调节(Cui et al.,2016;Gomes et al.,2014;Mooren et al.,2014)。生物信息学预测表明,表皮生长因子受体(epidermal growth factor receptor,EGFR)是miR-133a潜在靶点。基因分析显示,miR-133a与EGFR mRNA上的3’-UTR结合(不与突变的3’-UTR结合),从而下调蛋白表达水平,因此EGFR的蛋白表达随miR-133a表达的增加而减弱;另外,在乳腺癌细胞中,受运动锻炼调节的miR-133a能够抑制磷酸化Akt蛋白(phospho-pro‑tein kinese B,p-Akt)水平(Cui et al.,2013)。磷脂酰肌醇3-激酶(phosphatidylinositol 3-kinase,PI3K)和Akt是EGFR下游重要信号通路分子,激活的EGFR通过该通路将信号传递给PI3K蛋白,进而导致Akt蛋白磷酸化。因此,miR-133a在运动锻炼中表达增加,然后主动转运到肿瘤组织,而过表达的miR-133a通过靶向EGFR阻止PI3K的信号传导,进而抑制p-Akt蛋白水平,从而调节乳腺癌细胞的周期和增殖(Cui et al.,2013)。
运动锻炼能够增加miR-126和miR-146a的含量,miR-126过表达与乳腺癌细胞增殖减少及G1期阻滞有关(Al‑hasan,2019)。Alhasan(2019)研究发现,血管内皮生长因子A(vascular endothelial growth factor A,VEGF-A)通过促进血管内皮细胞增殖和肿瘤生长参与肿瘤转移,且在乳腺癌细胞中是miR-126的直接靶点。因此,过表达的miR-126通过直接调控VEGF-A抑制乳腺组织中癌细胞的生长和迁移。研究表明,miR-146a直接与白细胞介素-1受体相关激酶 1(interleukin-1 receptor-associated kinase 1,IRAK1)的3’-UTR结合,从而发挥肿瘤抑制作用(Long et al.,2019)。
4.2 运动锻炼通过调节miRNA改善前列腺癌的可能机制
前列腺癌被认为是最常见的癌症之一。研究发现,miR-206过表达能够改善前列腺癌(Wang et al.,2018),而运动锻炼能够增加血液循环中miR-206的含量。miR-206上调能够抑制前列腺癌细胞增殖、迁移,侵袭并诱导G0/G1阻滞,而前列腺癌组织中miR-206的表达明显降低(Wang et al.,2018)。膜联蛋白下调或功能障碍在恶性肿瘤的发生、发展中起重要作用,miR-206通过与膜联蛋白A2(annexin A2,ANXA2)mRNA的3’-UTR结合,调节上皮-间充质转化(epithelial-mesenchymal transition,EMT)信号(Yang et al.,2018)。由此,运动锻炼后,miR-206能够介导ANXA2,从而抑制前列腺癌细胞的侵袭和转移。
同样,miR-1含量也受到运动锻炼调节。miR-1通过阻滞G0/G1期进程抑制细胞生长,并且miR-1过表达能够从功能上抑制前列腺癌细胞的活性和增殖,miR-1靶向c-Met,显著降低p-Akt和p-mTOR的蛋白水平,从而阻滞Akt/mTOR信号通路以减少前列腺癌细胞的存活和增殖(Gao et al.,2019)。Hazar-Rethinam等(2011)研究表明,E2F家族成员E2F5和CDK家族成员PFTK1参与前列腺癌细胞周期的调控,而miR-1可直接调控这2个基因,进而抑制前列腺癌细胞增殖。
在前列腺癌组织中miR-133a-3p含量降低,尤其在骨转移性前列腺癌中。Tang等(2018)发现,miR-133a-3p通过直接靶向EGFR、成纤维细胞生长因子受体1、IGF-1R和肝细胞生长因子受体(mesenchymal-epithelial transition factor,MET)等多种细胞因子受体,阻滞PI3K/Akt信号通路,进而降低前列腺癌骨转移的发生率。
荧光素酶实验研究表明,ADAM金属蛋白酶结构域9(a disintegrin and a metalloprotease 9,ADAM9)参与多种癌症的发生和发展,是miR-126的靶基因(Hua et al.,2018)。目前,已证实miR-126能够降低ADAM9蛋白的表达,进而抑制前列腺癌进程(Hua et al.,2018),而运动锻炼能够增加miR-126含量。由此,运动锻炼可能通过调节miR-126来抑制前列腺癌的进程。
Xu等(2015)通过力学分析发现,miR-146a靶向Rho相关的含有卷曲螺旋蛋白激酶1(rho associated coiled coil containing protein kinase 1,ROCK1)的3’-UTR,抑制ROCK1的基础表达水平,同时miR-146a过表达能够降低(天冬氨酸特异性半胱氨酸蛋白酶3,cysteinyl aspartate specific protein‑ase,Caspase3)活性。因此,运动锻炼增加的miR-146a通过调节ROCK/Caspase3通路,在促进前列腺癌细胞凋亡过程中发挥重要作用(Xu et al.,2015)。
4.3 运动锻炼通过调节miRNA改善肝癌的可能机制
肝细胞癌是最常见的原发性肝脏恶性肿瘤之一。越来越多的证据表明,miRNA失调与包括肝癌在内的多种肿瘤的发生密切相关(Chen et al.,2016)。有研究发现,在27例人体肝癌组织中,miR-206的表达低于癌细胞旁正常组织(Wang et al.,2019),而运动锻炼能够上调miR-206。人类肝癌组织中c-Met水平与miR-206表达呈负相关,且miR-206针对c-Met基因的3’-UTR进行沉默,但c-Met表达的恢复可逆转miR-206对肝癌的抑制作用(Wang et al.,2019)。因此,miR-206主要通过靶向c-Met/PI3K/Akt/mTOR通路减少肝细胞生长因子诱导细胞EMT和血管生成,进而阻滞肝癌细胞增殖和细胞周期(Chen et al.,2016)。
内皮素-1(endothelin-1,ET-1)是一种强有力的促有丝分裂原,通过western blot和real-time PCR均检测到ET-1在人肝癌组织中高表达,而miR-1在肝癌细胞系中表达降低(Li et al.,2012)。荧光素酶报告分析表明,miR-1通过与ET-1的3’-UTR碱基互补结合位点配对来抑制ET-1表达,进而抑制肝癌细胞增殖(Li et al.,2012)。
功能研究表明,过表达的miR-133a通过抑制细胞增殖、迁移、侵袭和克隆形成,诱导细胞凋亡并阻滞细胞周期于G0/G1期,从而抑制体内肿瘤生长(Zhang et al.,2019b)。有研究证实,IGF-1R是肝癌细胞中miR-133a的直接靶点,miR-133a的过表达通过阻滞IGF-1R及其下游PI3K/Akt信号通路,进而抑制肝癌生长(Zhang et al.,2019b)。运动锻炼后,miR-133a含量显著增加,由此,miR-133a可能是运动锻炼影响肝癌进展的有效途径。
Polo样激酶 4(Polo-like kinase 4,PLK-4)属于致癌基因。生物信息学分析表明,PLK-4具有调节ATR/CHEK1通路参与肝癌的致癌作用。miR-126/PLK-4信号轴主要通过调节细胞增殖和细胞周期来抑制肿瘤的发生和发展。在此过程中,ATR/CHEK1通路受到显著抑制(Bao et al.,2018)。因此,受到运动锻炼调节的miR-126进一步负调控PLK-4的ATR/CHEK1通路,影响肝癌发展。
TNF受体相关因子6(TNF receptor associated factor 6,TRAF6)是肿瘤坏死因子受体相关因子家族的成员,其在许多癌症中是一种普遍扩增的癌基因(林根 等,2015)。研究表明,TRAF6在肝癌组织和细胞中表达上调,而受运动锻炼影响的miR-146a通过下调TRAF6抑制肝癌细胞增殖、侵袭和肿瘤生长(Zu et al.,2016)。
4.4 运动锻炼通过调节miRNA改善胃癌的可能机制
胃癌是世界上第四大常见癌症。与运动锻炼相关的miR-206能够抑制c-Met在胃癌中的表达,且在肿瘤中过表达的c-Met具有显著抑制肿瘤发展的作用(Zheng et al.,2015)。机理研究表明,配对盒基因3(paired box 3,PAX3)在胃癌组织中表达显著升高,能够促进胃癌细胞转移和侵袭(Zhang et al.,2015)。MET在人类多种癌症中上调,并在癌症发展中起重要作用(Han et al.,2015)。胃癌中,PAX3和MET水平呈正相关,且PAX3的功能是通过上调下游目标MET实现的(Zhang et al.,2015)。研究发现,PAX3是miR-206靶基因,由此,受运动锻炼调节的miR-206通过抑制PAX3的表达影响MET信号通路,从而抑制胃癌细胞转移(Zhang et al.,2015)。此外,Han等(2015)研究发现,miR-1通过靶向MET抑制胃癌细胞增殖和迁移。
Erb-B2受体酪氨酸激酶2(Erb-B2 receptor tyrosine ki‑nase 2, ERBB2)是一种跨膜酪氨酸激酶受体,属于EGFR家族,过表达的ERBB2能够促进细胞增殖,抑制细胞凋亡,导致细胞不受控或过度生长,其在人类肿瘤发生、发展中起重要作用(Li et al.,2017)。研究发现,miR-133a通过下调ERBB2及其下游信号分子p-Erk1/2和p-Akt的表达来抑制胃癌细胞增殖(Li et al.,2017)。而由于运动锻炼促使miR-133a含量增加,长期有氧耐力运动可能通过调节miR-133a影响胃癌进展。
双荧光素酶实验证实,miR-126通过靶定IGF-1R的3’-UTR,进而抑制IGF-1R表达,导致胃癌细胞增殖和侵袭降低(王宏 等,2019)。转化生长因子β激活激酶1(trans‑forming growth factor β-activated kinase 1,Tak1)是丝裂原活化蛋白激酶家族的成员,在NF-κB信号通路激活中发挥持久作用。大多数组织中,Tak1丢失使细胞对凋亡或死亡敏感(Mihaly et al.,2014)。此外,有研究发现,胃癌细胞中miR-146a表达与Tak1表达呈负相关(Chen et al.,2017)。因此,由运动锻炼诱导的miR-146a能够靶向Tak1,抑制NF-κB信号通路并降低Bcl2表达,进而抑制胃癌细胞增殖和迁移。
4.5 运动锻炼通过调节miRNA改善结直肠癌的可能机制
在结直肠癌中,miR-206作为肿瘤抑制因子,将结直肠癌细胞阻滞在G0/G1期,并加速细胞凋亡(Ren et al.,2016)。研究发现,miR-206过表达能够下调结直肠癌细胞中编导跨膜蛋白1(transmembrane 4 L six family member 1,TM4SF1),从而降低Akt和Erk磷酸化水平,影响结直肠癌细胞增殖、侵袭和转移(Park et al.,2018)。因此,miR-206作为运动锻炼和结直肠癌之间的潜在介质可能发挥重要作用。
过表达的miR-1模拟物能够显著降低肿瘤糖酵解,包括乳酸的产生和葡萄糖的摄取等,并最终抑制肿瘤增殖(Xu et al.,2017)。其主要机制是,miR-1通过与缺氧诱导因子-1α(hypoxia inducible factor 1α,HIF-1α)的3'-UTR结合,介导HIF-1α耗竭以抑制肿瘤糖酵解(Xu et al.,2017)。miR-1显著抑制Smad3与HIF-1α之间的相互作用,这种相互作用归因于HIF-1α减少,从而抑制Smad3激活,降低Warburg效应中代谢酶(如HK2和MCT4)的表达,进而最终抑制肿瘤增殖(Xu et al.,2017)。因此,运动锻炼可能通过显著增加miR-1改善结直肠癌。
研究发现,SUMO特异性蛋白酶1(SUMO-specific protease 1,SENP1)基因表达失调在多种癌症中均有表现,是癌症进展中关键的癌基因(Zhou et al.,2018)。荧光素酶基因实验表明,miR-133a-3p能够通过与其3'-UTR结合来调节SENP1的表达,导致SENP1下调和CDK抑制剂(如 p16、p19、p21和p27)上调(Zhou et al.,2018),从而在结直肠癌细胞增殖和调控细胞周期中发挥作用。
另外,miR-126和miR-146a在结直肠癌中含量下降(Bleau et al.,2018;Yuan et al.,2016)。趋化因子(C-X-C基序)受体4(C-X-C motifchemokine receptor 4,CXCR4)是7种跨膜G蛋白偶联受体成员,对多种细胞动员、迁移、增殖和存活起至关重要的作用,且CXCR4在各种癌症类型中高度表达,被认为是表达最广泛的癌症相关趋化因子受体(Chatterjee et al.,2014)。在结直肠癌中,miR-126通过CXCR4失活RhoA信号通路发挥肿瘤抑制作用(Yuan et al.,2016)。研究发现,miR-146a能够阻止 c-Met的翻译,miR-146a的过表达在肿瘤转移克隆体中导致恶性肿瘤减少,并在一定程度上消除原发性肿瘤(Bleau et al.,2018)。由此,运动锻炼可能通过调节miR-126和miR-146a改善结直肠癌。
4.6 运动锻炼通过调节miRNA改善肺癌的可能机制
非小细胞肺癌(non-small cell lung cancer,NSCLC)是全球癌症相关死亡的主要原因,血管生成是NSCLC的主要标志。研究发现,miR-206在NSCLC中含量显著下降(Xue et al.,2016)。在NSCLC中,miR-206通过抑制14-3-3ζ/STAT3/HIF-1α/VEGF通路降低NSCLC的血管生成能力(Xue et al.,2016)。具体机制是 ,14-3-3ζ与磷酸化的STAT3结合,提高HIF-1α表达,增强HIF-1α向VEGF启动子的募集,导致14-3-3ζ增强血管生成。然而,在NSCLC细胞异种移植模型中,miR-206过表达或14-3-3ζ表达减少均抑制STAT3/HIF-1α/VEGF通路,并降低肿瘤的生长和血管生成(Xue et al.,2016)。在另一条通路中,miR-206通过抑制c-Met和Bcl2在NSCLC中的表达,在肿瘤中发挥肿瘤抑制作用(Sun et al.,2015)。另外,miR-1和miR-206均抑制c-Met下游的Akt和Erk通路,并阻断肝细胞生长因子诱导的EMT(Jiao et al.,2018)。由此,miR-1和miR-206可能介导运动锻炼调节NSCLC的相关信号通路。
在NSCLC患者中发现,miR-133a下调与NSCLC患者的预后不良相关(Shen et al.,2019)。YES原癌基因1(YES proto-oncogene 1,YES1)也被确认为是miR-133a的直接靶标,且YES1下调能够抑制NSCLC细胞增殖(Shen et al.,2019)。因此,运动锻炼诱导的miR-133a可能通过靶向YES1,抑制NSCLC中的细胞增殖。
PTEN/PI3K/Akt通路被认为是调控细胞增殖、生长、代谢和凋亡等生物学过程的重要信号通路(Song et al.,2016)。miR-126上调导致PIK3R2、PI3K和p-Akt表达下降,PTEN表达升高(Song et al.,2016);相反,miR-126下调,这3种蛋白表达量增加。双荧光素酶基因检测结果表明,PIK3R2是miR-126的靶基因(Song et al.,2016)。因此,在NSCLC细胞中过表达的miR-126能够降低PIK3R2的表达,影响PTEN/PI3K/Akt信号通路,从而抑制肿瘤细胞增殖、迁移和侵袭(Song et al.,2016)。
在NSCLC中,miR-146a-5p通过靶向调节细胞周期因子(CCND1和CCND2)mRNA的3’-UTR位点,在mRNA和蛋白质水平上均降低细胞周期因子的表达,从而导致细胞周期停滞在G0/G1期,进而抑制NSCLC细胞系中的细胞增殖和细胞周期进程(Li et al.,2016)。由此,运动锻炼可能通过诱导miR-146a抑制NSCLC细胞增殖。
综上所述,运动锻炼可能通过调节miR-1、miR-126、miR-133a、miR-146a、miR-206a含量,对乳腺癌、前列腺癌、肝癌、胃癌、结直肠癌、肺癌具有良好的改善作用。根据miRNA的生理生化特性,miRNA能够调控mRNA转录的基因进行切割、失稳或翻译抑制,进一步抑制相关基因表达。miRNA调控癌基因的具体机制为miRNA通过与mRNA的3’-UTR部分碱基互补配对,诱导mRNA翻译、降解和沉默,导致细胞有丝分裂G0/S期停滞,阻断癌细胞新蛋白质形成和DNA复制,从根源上切断癌细胞复制和转移。运动锻炼通过调节miRNA改善相关癌症的进程中,运动锻炼诱导的miRNA主要通过调节c-Met/Akt/mTOR、c-Met/Akt/Erk和c-Met/PI3K/Akt信号通路阻滞细胞有丝分裂G0/S期,进而有效控制癌细胞更新、侵袭和迁移(图2)。
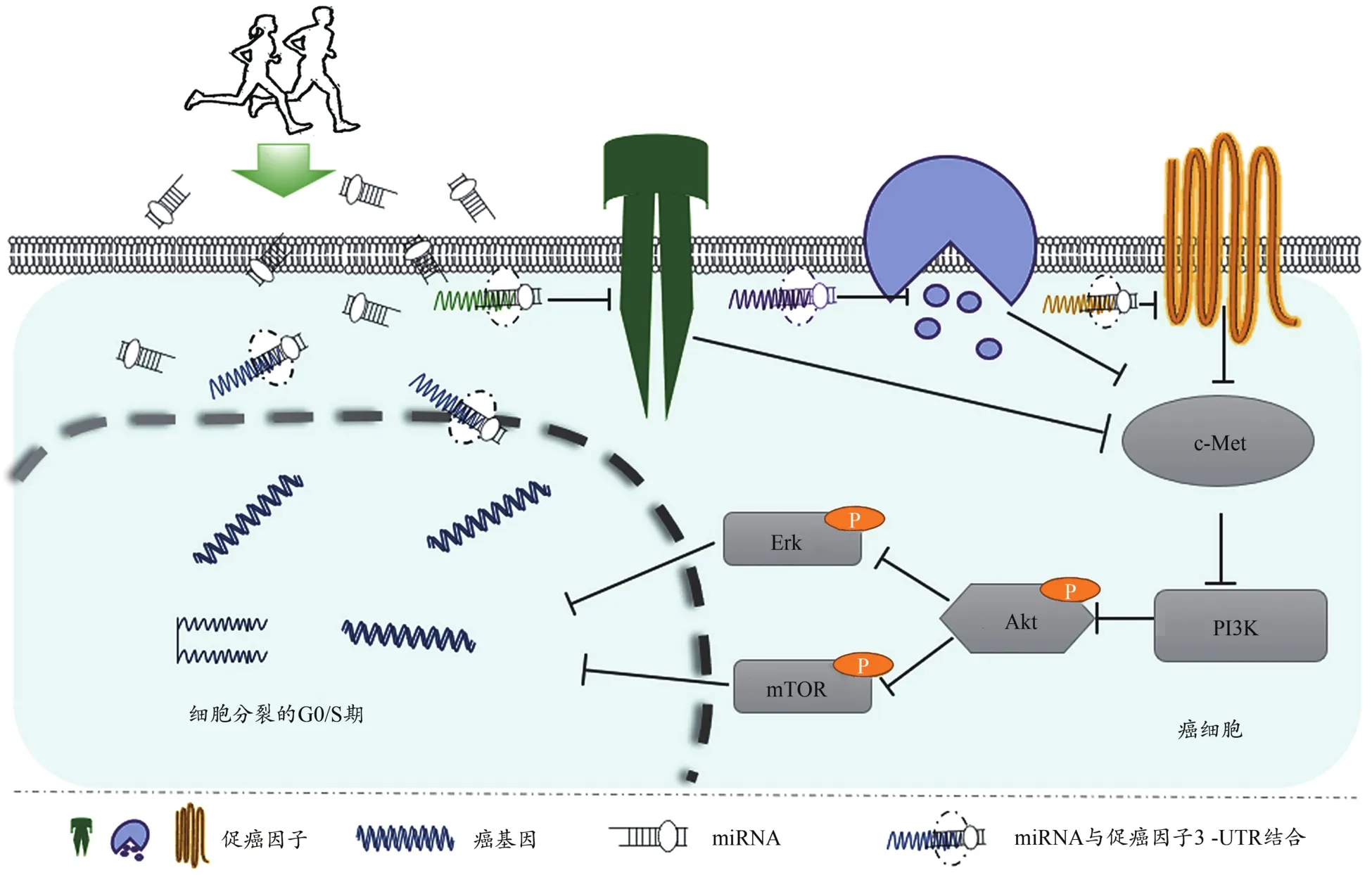
图2 运动锻炼通过调节miRNA改善癌症的机制Figure 2. The Mechanism of Physical Exercise Improving Cancer by Regulating miRNA
5 结论
运动锻炼能够调节血浆中运动敏感型miRNA的含量,可能对乳腺癌、前列腺癌、肝癌、胃癌、结直肠癌、肺癌具有良好的改善作用,且运动锻炼诱导的相关miRNA可能通过调节相关信号通路阻滞细胞有丝分裂的G0/S期,从而有效控制癌细胞更新、侵袭和迁移。目前,有关运动锻炼对miRNA表达的影响受到样本量小的限制,且未对miRNA或身体活动进行量化。更重要的是,鲜见运动锻炼诱导miRNA改善癌症的针对性研究,未来需要进一步探讨其具体机制。
猜你喜欢
家庭医学(下半月)(2020年3期)2020-05-30 12:42:02
家庭医学(下半月)(2020年3期)2020-05-30 12:42:00
家庭医学(下半月)(2020年3期)2020-05-30 12:42:00
家庭医学(下半月)(2020年1期)2020-05-11 02:05:32
天津医科大学学报(2019年3期)2019-08-13 06:53:08
中国生殖健康(2019年7期)2019-01-06 09:27:34
海峡姐妹(2018年7期)2018-07-27 02:30:36
特别健康(2018年4期)2018-07-03 00:38:08
特别健康(2018年2期)2018-06-29 06:13:42
肿瘤预防与治疗(2015年1期)2015-09-26 07:26:20
