
TTN基因突变致儿童扩张型心肌病的研究进展
2023-02-17 09:35郑奎娄美娜综述张英谦审校
中国当代儿科杂志 2023年2期
郑奎 娄美娜 综述 张英谦 审校
(1.河北省儿童医院心内科/河北省小儿心血管重点实验室,河北石家庄 050031;2.河北医科大学研究生学院,河北石家庄 050017)
扩张型心肌病(dilated cardiomyopathy,DCM)是儿童心肌病中最常见的类型,占50%~60%。儿童DCM的病因复杂,常导致患儿心力衰竭或心源性猝死,其显著的特点是进展快和病死率高[1-2]。其中遗传病因在儿童DCM中占有重要地位,30%~40%的患儿与遗传相关[1-2]。编码肌联蛋白(titin)的TTN基因截断突变(TTN-truncating variant,TTNtv)是成人DCM最常见的遗传突变,在家族性DCM中约占25%,散发性DCM病例中约占18%[3]。由于成人DCM发病率高,关于成人DCM的遗传研究已取得了一定的进展[4],但关于儿童的DCM遗传研究比较落后,其治疗也多借鉴于成人DCM的治疗方案。多项研究报道TTNtv导致的成人DCM预后良好,对治疗有良好的反应,但对于儿童患者来说多数预后不良,而且容易发生恶性心律失常和心源性猝死[1,5]。当前已有基因治疗用于杜氏肌营养不良症[6]。本文总结了TTN基因突变与儿童DCM发病的关系、基因型-表型关系及相关治疗研究进展,为DCM患儿的预后评估、个体化管理和精准治疗,包括未来基因治疗的研究,提供参考。
1 肌联蛋白与肌节
肌联蛋白由TTN基因编码,位于2号染色体q31区段,包含364个外显子,广泛的mRNA剪接可产生多种肌联蛋白亚型,与心脏相关的亚型主要为N2B(3 MDa)和N2BA(3.2~3.8 MDa)[4]。此外,还包括少量novex-3亚型,以及最近新发现的C-端Cronos亚型[7-8]。肌节是横纹肌收缩的最基本单位。肌联蛋白是肌节的重要组成部分,主要由4个亚基组成,分别对应肌节的4个结构域:Z盘、I带、A带和M线,从Z盘到M线纵向横跨半个肌节,对肌节的组装、机械感应和信号传递发挥着重要作用,同时决定着肌原纤维的延伸性和被动张力[9]。肌联蛋白的末端被T-cap3锚定在Z盘N端区域,为肌节的机械传感发挥着重要作用[10]。I带区域毗邻Z盘,具有高度弹性特征,提供类似“弹簧样”功能,主要由免疫球蛋白样结构域、PEVK区(富含脯氨酸、谷氨酸、缬氨酸和赖氨酸残基)和N2B独特序列(N2B-us)组成[10]。A带区域内主要由相对稳定的纤维连接蛋白3型和免疫球蛋白样超重复序列组成,主要参与调节粗肌丝的长度。A带区域毗邻并嵌入M带的羧基末端丝氨酸-苏氨酸激酶结构域,参与许多信号通路的传导,调节肌节主动收缩力[10-11]。
2 TTNtv可能导致DCM发病的分子机制
当前国内外关于TTNtv导致儿童DCM发病机制的研究报道较少,大多是关于成人DCM的遗传研究。有研究报道在成人DCM中18%~25%的患者与TTNtv相关,此外2%~3%的健康人群携带TTNtv[3-4,8,11]。不同队列研究报道儿童 DCM 人群中TTNtv的发生率有差异,有研究表明DCM患儿中很少发生TTNtv[12]。而Khan等[13]对106例儿童DCM遗传病因分析发现约9%的患儿携带TTN基因突变,并发现DCM患儿的发病年龄与TTNtv检出率密切相关,青少年(26%)明显高于婴幼儿(6%)。Wang等[1]对我国46例DCM患儿进行遗传病因研究,显示约13%的患儿与TTNtv相关。TTNtv导致DCM的确切机制尚未得到明确的证实,其分子发病机制也一直存在争议。在具有TTNtv遗传性家族史的DCM患者中,遵循常染色体显性遗传,但同时明显存在不完全和年龄依赖的外显率[4,11]。肌联蛋白可通过多种机制调节其弹性及被动张力,当前认为包括TTN截断发生的位置、肌联蛋白亚型转录、翻译后修饰(氧化、磷酸化等)、单倍体剂量不足、环境因素影响等与DCM的发生、发展密切相关。此外也有研究认为TTNtv可直接产生异常的肌联蛋白,并通过一种“毒肽”机制对肌节和心肌细胞造成直接损害,但当前认为“毒肽”模型机制尚未在人类心脏研究中得到证实[4,11,14],下文将进行简要介绍。
2.1 TTNtv发生的位置与其致病性相关
DCM最常见的遗传病因是TTNtv,其多为杂合子变异,包括错义突变、框移突变、剪切突变、无义突变、重复突变和拷贝数变异等,这些突变预测可能改变肌联蛋白的氨基酸序列[4,15]。此外,有报道显示纯合子或非典型剪接TTN基因突变也可致儿童严重的DCM表型[8]。基因型-表型分析研究表明DCM的表型及其严重性与TTNtv发生的位置高度相关[15]。多项研究表明TTNtv多发生于A带区域,少部分TTNtv发生于I带、Z盘和M线区域,并且发生于A带或M线区域的TTNtv患儿常表现为更严重的DCM,预后差[14-16]。A带区域含有高比例的不对称外显子,TTNtv可能会改变翻译阅读框,使其更容易产生异常的肌联蛋白[16]。有超过1/3的TTNtv可进行选择性剪接,特别是发生于I带区域的外显子[16]。选择性剪切补救机制,可减轻TTNtv产生的影响,因此发生于I带区域的DCM表型相对较轻[16]。Z盘区域肌联蛋白表达减少可能导致Z盘结构破坏和功能受损[17]。研究认为Z盘区域TTNtv的致病性较低,可能是与Z盘TTNtv外显子的组成性表达较低相关[18]。Biquand等[19]研究显示对发生于M线区域Mex5外显子(编码插入序列7)缺失的小鼠可导致严重的DCM表型,其心室进行性扩张,心室肌伴有广泛的纤维化,同时还伴有兴奋-收缩偶联蛋白的异常表达。携带位于M线区域的TTNtv患儿常表现为早期危及生命的DCM表型,多数患儿病程早期即发生死亡[8,15]。儿童DCM早期发病或预后不良是否与TTNtv多发生于A带、M线区域或同时合并其他致病基因相关有待进一步研究。
2.2 肌联蛋白亚型的转录调控可能与DCM相关
调控肌联蛋白亚型转录可能是治疗收缩和/或舒张功能障碍性DCM的潜在靶点。在健康成年人的心肌组织中(主要为左心室)肌联蛋白N2BA亚型占30%~40%,N2B亚型占60%~70%,同时在正常衰老过程中此比例也保持相对恒定[7-8]。N2B亚型和N2BA亚型的正常比例,决定着肌联蛋白的延伸性和被动张力,如N2BA亚型比例增多则肌原纤维被动张力降低,其心室收缩功能障碍;N2B亚型占比增多则肌原纤维变得僵硬,其心室舒张功能降低[4,7-8]。有研究发现在终末期DCM患者中,其肌联蛋白亚型主要为较长、较柔软的N2BA亚型[11]。广泛的mRNA剪接可产生不同类型的肌联蛋白亚型,N2BA亚型的表达可由剪接因子RNA结合基序蛋白-20(RNA binding motif protein 20,RBM20) 调控[11]。RBM20是一种RNA剪接因子,参与mRNA成熟的过程,可与剪接位点附近的内含子、U1和U2小核核糖蛋白(snRNP)结合位点结合,调控肌联蛋白亚型转换[11]。RBM20缺失可使肌联蛋白N2BA亚型的表达显著升高,致使肌原纤维主动和被动张力降低,从而导致心室扩张[11]。对于舒张性心力衰竭,有研究表明包括地高辛和洋地黄在内的强心苷,可以有效降低RBM20水平,可使肌联蛋白更倾向于N2BA亚型表达,以改善其舒张功能[11]。有研究发现胰岛素和三碘甲腺原氨酸都能调节RBM20的表达和活性通路[7]。最近有学者对大鼠进行研究,发现三碘甲腺原氨酸或胰岛素可上调RBM20表达和激活雷帕霉素靶蛋白(mechanistic target of rapamycin,mTOR)激酶轴,从而使肌联蛋白倾向于N2B亚型表达[11,20]。另一种可以转录修饰肌联蛋白的分子是RNA结合基序蛋白24。有研究表明,在小鼠动物模型中敲除RNA结合基序蛋白24可导致DCM[11]。该研究也显示,微RNA能够通过与mRNA结合,使mRNA靶向降解来调控肌联蛋白的转录,并在携带TTNtv的DCM小鼠模型中证实了一种微RNA(miR-208b)显著表达[11]。但miR-208b对肌联蛋白亚型调控的确切机制尚未不明确。
2.3 肌联蛋白弹簧元件的翻译后修饰
除了转录修饰外,肌联蛋白也可通过翻译后磷酸化、氧化、乙酰化等修饰改变其肌原纤维延伸性和被动张力[4,7]。肌联蛋白最常见的翻译后修饰是磷酸化[7]。有研究发现携带TTNtv的DCM患者,其左心室中的肌钙蛋白I和肌联蛋白结合蛋白-C(myosin binding protein-C,MyBP-C)的磷酸化水平显著降低,肌联蛋白磷酸化水平的降低可加重心室功能进一步恶化,最终导致心力衰竭[17]。基于蛋白质组学分析,预测肌联蛋白上有数百个磷酸化位点,但当前认为只有少数几个磷酸位点与肌联蛋白的结构和功能相关,其大多位于I带区域,主要包括PEVK和N2Bus区域[4,8,10]。I带区域内的PEVK元件含有丰富的脯氨酸、谷氨酸、缬氨酸和赖氨酸,具有高度的可扩展性。当前认为N2Bus元件也具有许多磷酸位点,由几种不同的激酶介导,包括蛋白激酶A、蛋白激酶G、细胞外信号调节激酶2和钙调素激酶Ⅱδ等[8]。差异性磷酸化已被证明可直接改变心肌细胞的长度和张力,激活PEVK元件可增强心肌细胞被动张力有助于心室收缩,避免心室进一步扩张;然而激活N2Bus元件的磷酸化可使心肌纤维变长、被动张力降低,加重心室扩张[8]。Hidalgo等[21]通过激活蛋白激酶C磷酸化PEVK元件,发现小鼠和猪的心肌细胞被动张力可增加20%~30%。通过调控肌联蛋白上的磷酸位点有望治疗DCM,但其具体机制仍需进一步研究。
2.4 单倍体剂量不足
当前关于单倍体剂量不足的机制导致DCM仍存在一定争论。单倍体剂量不足动物模型显示TTNtv并不直接损伤肌节,而是通过增加代谢应激,从而对心肌细胞和心室功能产生慢性毒害作用[22]。TTNtv导致产生异常的mRNA转录本,致使无意义mRNA衰减增加,由此TTN等位基因表达减少,基因表达减少可能增加代谢应激,代谢的改变可导致mTOR复合物-1信号通路激活,从而致使异常肌联蛋白合成和自噬相关的致病反应,最终导致DCM表型[4,11]。有研究对TTNtv大鼠单倍体剂量不足建模研究,并用RNA-seq研究揭示了TTNtv等位基因介导的无意义mRNA衰减增加表明单倍体剂量不足[22]。除单倍体剂量不足外,有研究还发现TTNtv导致大鼠心肌耗氧率降低,活性氧水平升高,线粒体蛋白泛素化水平升高,提示TTNtv导致线粒体功能障碍[23-24]。此外还有研究发现TTNtv导致DCM患者的iPSC心肌细胞对异丙肾上腺素、Ca2+和血管紧张素Ⅱ等反应减弱[11]。总之,由TTNtv导致的DCM中可观察到心肌代谢、线粒体功能和钙离子处理等异常,这些病理性改变是直接来自TTNtv,还是机体代偿性改变带来的病理变化也有待研究证实。
2.5 TTNtv导致的DCM与环境因素相关
TTNtv导致DCM的发病年龄常见于0~40岁,但当与年龄、性别、药物、酒精、妊娠或获得性疾病等风险因素相结合时,可加重DCM[11]。发病年龄越小,其预后多不良,对传统的治疗疗效欠佳[1]。化疗药物也可诱发、加重TTN基因突变相关的儿童DCM[11]。最近在蒽环类化疗药物诱导的心肌病患者中发现了TTNtv,同时还观察到TTNtv可能与化疗药物诱导的严重心肌病相关[11,25]。酒精是一种常见的环境风险因素,与TTNtv导致的DCM预后不良相关,在酗酒伴TTNtv的DCM患者中可以观察到其左心射血分数下降得更显著[26]。同时TTNtv还与围生期心肌病的风险增加有关[11]。TTNtv的外显率还与性别相关。有研究显示男性DCM发病的中位年龄为28岁,女性为56岁,携带TTNtv的女性DCM患者比男性患者预后好[27]。雌激素是否对TTNtv导致的DCM具有保护作用,从而导致了性别相关的差异,有待进一步研究。
3 TTNtv可导致的临床表型
儿童DCM具有显著的遗传异质性,患儿同时可携带多个基因变异,受环境因素的影响及发病年龄依赖的外显率等特点,可使患儿的临床表型时有重叠[2,11]。编码肌联蛋白的基因突变与多种骨骼肌病和心肌病相关,其中最常见的表型为DCM,其相关临床表型见表1[8,11,25]。TTNtv携带者的发病年龄与其临床表型的严重程度密切相关,对于儿童时期发病的TTNtv患者常表现为危及生命的 DCM表型,预后不良[1,13]。成人TTNtv携带者通常会在中年或晚年才表现出DCM相关临床症状,但有证据表明TTNtv携带者在出现DCM相关症状前,心脏MRI可检测到其心功能障碍和/或偏心性心室重构[11]。此外,包括肥厚型心肌病(hypertrophic cardiomyopathy,HCM)、限制型心肌病(restrictive cardiomyopathy,RCM)、致心律失常型右心室心肌病( arrhythmogenic right ventricular cardiomyopathy,ARVC)和围生期心肌病等也有报道[11,25]。在 HCM 中,TTNtv占比约为 1.3%~4%,但当前对RCM、ARVC携带TTNtv报道得较少[8]。也有研究报道TTNtv可能与约16%的家族性房颤和约 5% 的早发性房颤相关[28]。Enriquez等[29]也研究发现携带TTNtv的DCM患者中,约65%的患者发生过快速性房性心律失常,并且多数患者早期只表现为快速性房性心律失常,随着病程进展才出现DCM相关表现;随访2年并发现约53%的患者发生过室性心动过速复发,约27%的患者需要心脏移植、机械循环支持或死亡。特发性/家族性DCM中携带TTNtv可作为预测早期发生心律失常的独立危险因素,携带TTNtv的DCM患者有增加发生室性心律失常的风险,同时还应考虑到与年龄依赖相关的外显率,并建议对未受影响的携带者进行长期随访[30]。
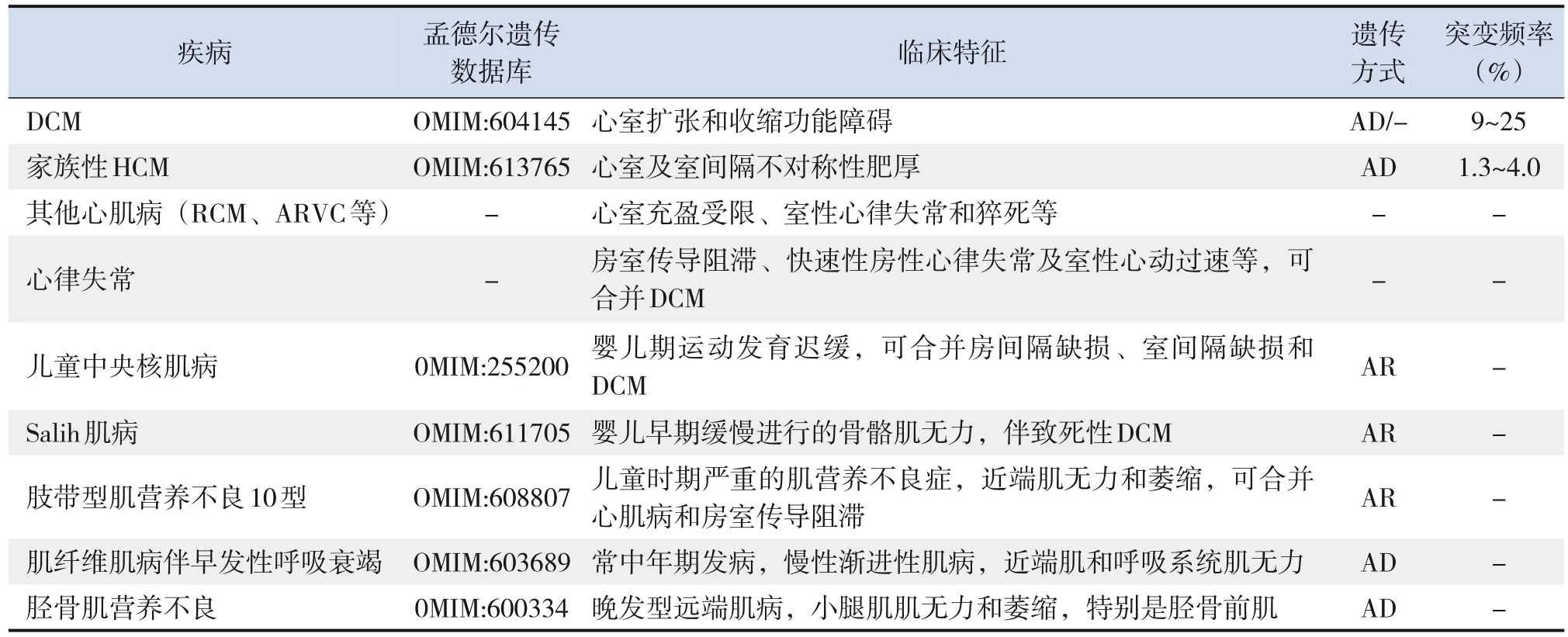
表1 与TTN基因变异相关的临床表型/疾病
4 基因治疗有望治愈TTNtv相关的DCM
当前儿童DCM的治疗方法多为纠正紊乱的继发性病理生理改变,包括神经体液过度激活和心室重构。对于基因治疗,反义寡核苷酸(antisense oligonucleotide,AON)介导的外显子跳跃疗法可能是治愈TTNtv导致DCM的潜在手段。在AON介导的这种治疗方法中,单链寡核苷酸被设计为结合突变内含子-外显子边界前mRNA,或阻断外显子剪接基因序列,其目标是通过选择性剪接跳过包含移码突变的外显子,从而恢复阅读框防止翻译过程中肌联蛋白的早期终止[11,31-32]。美国食品药品监督管理局已经批准了一种外显子跳跃疗法用于杜氏肌营养不良症[6],希望类似的基因治疗策略也可用于治疗TTNtv导致的DCM。AON介导的外显子跳跃疗法治疗DCM的可行性已经在小鼠模型中得到了验证,该方法可使肌联蛋白表达正常,并可挽救TTN基因突变导致DCM表型,但目前尚不清楚这种治疗策略是否能够逆转已经存在的DCM表型[32]。此外还应该特别关注外显子跳跃治疗也可能会发生脱靶效应[10]。此外,包括mTOR抑制剂雷帕霉素和人诱导多能干细胞来源的心肌细胞(human induced pluripotent stem cell-derived cardiomyocyte,hIPSC-CM)治疗TTNtv相关DCM也有研究报道[11],但当前认为hIPSC-CM技术存在一定局限性,其产生的心肌细胞相对不成熟,常表达较柔软的N2BA亚型,其功能效应也有待进一步验证。总之AON介导的外显子跳跃疗法为治愈DCM带来了新希望,未来外显子跳跃疗法很有可能成为突破传统DCM的治疗手段。
综上所述,TTNtv是DCM最常见的遗传突变,尽管当前对TTNtv导致DCM的发病机制存在较大争议,但研究了解TTN基因突变相关儿童DCM的分子发病机制,有助于这部分患儿的预后评估及精准治疗。TTNtv导致儿童DCM的遗传分子机制与成人是否存在差异,也有待进一步研究。此外,临床工作中如何精准评估TTNtv的致病性也是当前所要面临的重大挑战。随着基因检测成本的持续下降,基因诊断、精准治疗将会是未来医学发展的大趋势。进一步深入研究TTNtv导致DCM和心律失常的遗传分子机制,可为精准治疗TTN基因突变相关的DCM患者带来新希望。
猜你喜欢
电子科技大学学报(2022年5期)2022-10-29
中国生殖健康(2020年4期)2021-01-18
中国临床医学影像杂志(2019年1期)2019-04-25
中国生殖健康(2018年4期)2018-11-06
现代园艺(2017年21期)2018-01-03
中国中医药信息杂志(2016年5期)2016-12-01
中国康复理论与实践(2015年10期)2015-12-24
医学研究杂志(2015年5期)2015-06-10
国际心血管病杂志(2015年5期)2015-02-27
现代检验医学杂志(2015年5期)2015-02-06
