
白芥子诱导的鸡内源性芥子碱降解菌的筛选、鉴定及酶活测定
2023-02-16 07:18:40范景胜邹成义郑钰嘉倪青松汪林书
中国饲料 2023年3期
邓 卉,余 丹,范景胜,屈 东,邹成义,李 斌,殷 勤,郑钰嘉,倪青松,汪林书
(四川省畜牧科学研究院,动物遗传育种四川省重点实验室,四川成都 610066)
我国优质饲料原料高度依赖进口,2020年进口大豆超1亿t,在新冠疫情和中美贸易战的双重压力下,原料价格进一步飞涨。由此,《国务院办公厅关于促进畜牧业高质量发展的意见》(国办发〔2020〕31号)中明确提出促进菜籽粕等非粮饲料资源高效利用,促进豆粕减量替代。芥子碱作为菜籽粕、白芥子等十字花科类植物原料的重要抗营养因子,极大限制了该类原料在工业饲料中的应用。利用微生物自身分泌的酶降解芥子碱的生物技术方法是目前最安全、最有效的手段之一,本研究通过白芥子诱导试验,从鸡的盲肠中筛选出一株能有效降解芥子碱的细菌,经鉴定为大肠杆菌,并进一步考察了该菌及其功能酶漆酶在不同培养时间对芥子碱的降解效果,为高效利用十字花科类非粮型饲料原料提供技术支撑。
1 材料与方法
1.1 培养基 芥子碱液体培养基,由(NH4)2SO4、NaCl、K2HPO4、MgSO4.7H2O、KCl、FeSO4、芥子碱硫氰酸盐和蒸馏水组成;伊红美蓝固体培养基;LB液体培养基;芥子碱+LB液体培养基(芥子碱:LB=3:2)。
1.2 菌株筛选与鉴定
1.2.1 动物试验日粮及采样 在藏香鸡基础日粮中添加20%白芥子饲喂28 d后,无菌条件下分别采集鸡空肠、回肠、盲肠内容物存放于无菌生理盐水中,待分离培养目标菌株。基础日粮组成及营养水平见表1。
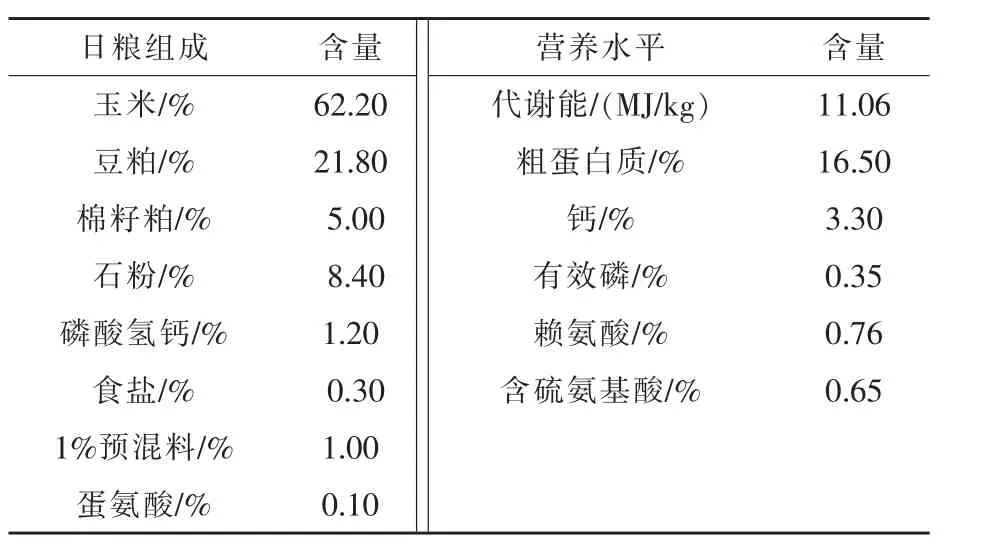
表1 基础日粮组成及营养水平
1.2.2 分离培养目标菌株 将采集的鸡空肠、回肠、盲肠内容物分别接种于芥子碱+LB液体培养基中,37℃、180 r/min摇床振荡培养48 h。取芥子碱+LB培养液在伊红美蓝固体培养基上划线,37℃恒温培养24 h。挑取伊红美蓝培养基上单个菌落到芥子碱液体培养基中,37℃、180 r/min摇床振荡培养72 h。再取芥子碱培养液在伊红美蓝固体培养基上划线,37℃恒温培养36 h。最后挑取伊红美蓝培养基单个菌落到LB液体培养基中,于37℃富集培养12 h,将获得的纯化菌株冻存-20℃冰箱备用。
1.2.3 菌株种属鉴定 形态学鉴定:采用形态学鉴定法观察伊红美蓝培养基上菌株的培养特征和菌落形态,并结合革兰氏染色法在100×显微镜下观察菌株染色形态特征。参照《伯杰细菌鉴定手册》初步鉴定其种属。
分子生物学鉴定:提取获得菌株基因组DNA,以通用引物进行PCR扩增。引物序列为27F:AGTTTGATCMTGGCTCAG;1492R:GGTTACCTTG TTACGACTT。PCR反应条件为:98℃3 min;98℃10 s,55℃15 s,72℃10~15 s/kb,39个循环;72℃5 min。扩增产物进行琼脂糖凝胶电泳分离、检测,切胶纯化后送至北京擎科生物科技有限公司测序,所得16SrDNA基因序列在GenBank里进行序列比对,下载GenBank中同源性较高序列,并利用MEGA-X软件的Neighbor-Joining法构建系统发育树。
1.3 测定获得菌株降解芥子碱效果 将获得菌株接种到芥子碱液体培养基中,空气摇床37℃培养72 h。从第0 h开始,每隔8 h测定培养液中芥子碱的含量,比较获得菌株在不同培养时间对芥子碱的降解效果。设置3个重复,结果取平均值。
采用紫外分光光度法测定芥子碱含量:精密称取芥子碱硫氰酸盐对照品(含量≥98.0%)37.0 mg,溶于10 mL蒸馏水中制成溶液。分别精密吸取上述溶液3、4、5、6、8、10、15、20μL,用水稀释至3 mL,摇匀后即得浓度分别为0.0037、0.0049、0.0062、0.0074、0.0099、0.0123、0.0185、0.0247 mg/mL的芥子碱对照品溶液,以蒸馏水为空白对照,在326 nm波长处测定吸光度。以吸光度A为纵坐标,浓度C为横坐标绘制标准曲线并进行线性回归分析。取样品培养液15μL加蒸馏水稀释至3 mL,摇匀后在326 nm波长下测定吸光度,根据标准曲线计算出培养液芥子碱含量。
1.4 测定获得菌株漆酶活力 以2,2'-连氮基-双(3-乙基苯并二氢噻唑啉-6-磺酸)(ABTS)为底物,分别在第8、16、24、32、40、48 h采用可见分光光度计在420 nm下测定培养液漆酶活力。以每毫克蛋白每分钟氧化1 nmoL底物ABTS所需的酶量为一个酶活力单位(U)。
1.5 统计分析 运用Excel对相关数据绘制回归曲线、建立回归方程以及确立复相关指数R2,再采用SPSS 17.0软件对回归方程进行方差分析(ANOVA),以P<0.05作为差异显著的判断标准,以P<0.01作为差异极显著的判断标准。
2 结果与分析
2.1 菌株筛选 初步筛选结果显示,在肠道不同位置取样会影响目标菌的培养,本试验条件下,盲肠中培养出的目标菌明显多于空肠和回肠(表2)。

表2 鸡肠道不同取样位置条件下目标菌的培养情况
选择筛选数最多的盲肠段目标菌继续培养观察,发现上述盲肠中筛选出的9株菌均能够利用芥子碱作为营养物质生长,肉眼观察其中1株目标菌在芥子碱培养液中浑浊度明显高于其他8株,并且在伊红美蓝培养基上的生长情况明显优于其他8株,将该菌株命名为SDB2(Sinapine-degrading bacteria 2)。
2.2 SDB2种属鉴定
2.2.1 形态学鉴定 在伊红美蓝培养基上,菌株SDB2长满整个平板,菌落呈圆形、突起、边缘规则的形状且带有翠绿色金属光泽,初步判定为典型的大肠杆菌菌落。采用革兰氏染色法对单菌落染色后在100×显微镜下观察后判定该菌为革兰氏阴性短杆菌。
2.2.2 分子生物学鉴定 由图1可知,菌株SDB2属于Escherichia sp,273-c菌株。因此将SDB2菌株鉴定为变形菌门;γ-变形菌纲;肠杆菌目;肠杆菌科;大肠埃希菌。

图1 基于16S rDNA序列构建SDB2菌株的系统发育树
2.3 SDB2降解芥子碱效果研究 由图2可知,吸光度A与浓度C建立的线性回归方程为A=54.1106C+0.0329,复相关指数R2=0.9989。
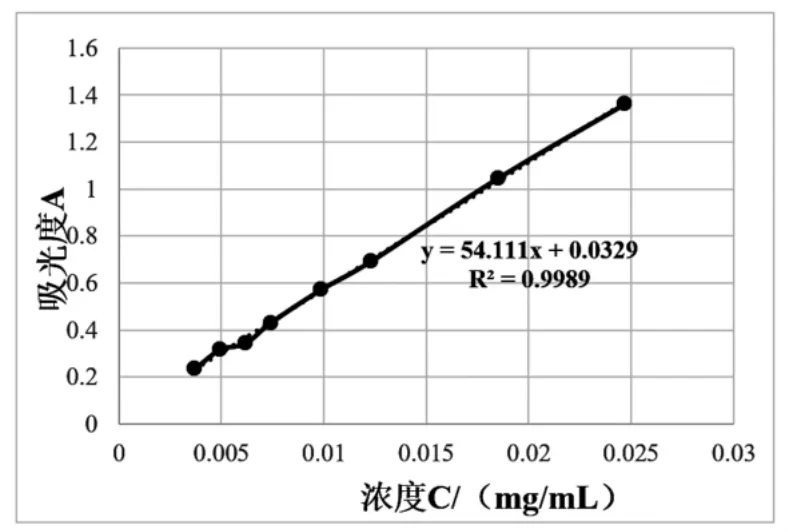
图2 芥子碱对照品标准曲线
由表3可知,芥子碱浓度随培养时间增加,总体上呈降低趋势,第72 h浓度最低,降低幅度达50%。
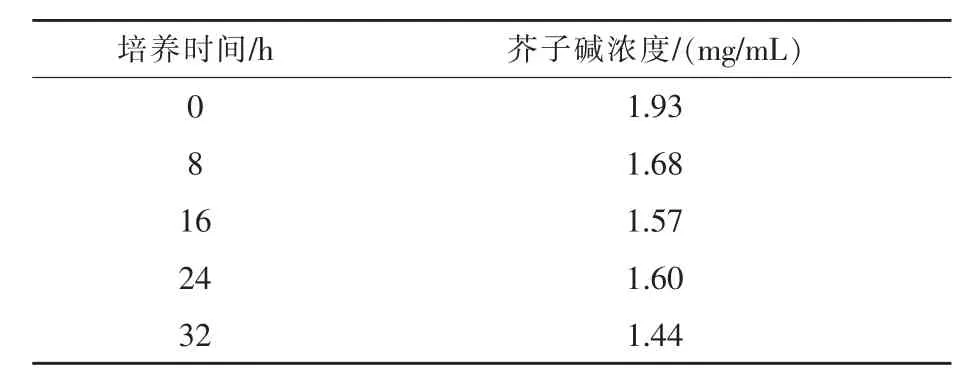
表3 SDB2菌株不同培养时间对芥子碱浓度的影响
进一步对芥子碱浓度与培养时间建立回归方程并做方差分析,结果如图3和表4所示。芥子碱浓度与SDB2菌株培养时间存在极显著的线性回归关系(P<0.01),复相关指数R2=0.9368,芥子碱浓度随培养时间增加而降低。
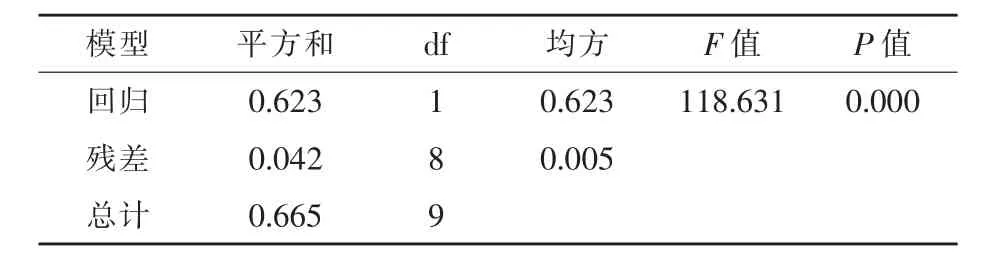
表4 芥子碱浓度与SDB2菌株培养时间建立回归方程的方差分析
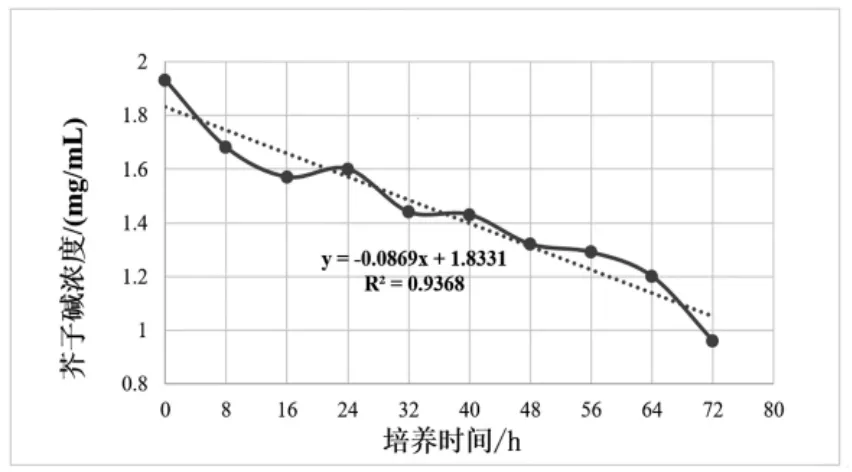
图3 芥子碱浓度与SDB2菌株培养时间的线性回归关系
2.4 SDB2漆酶活力 分别在第8、16、24、32、40、48 h测定芥子碱培养液的漆酶酶活,结果由表5可知,SDB2菌株漆酶活力第8 h最高,达7.38 U/mL,之后漆酶活力逐渐降低,第48 h最低。
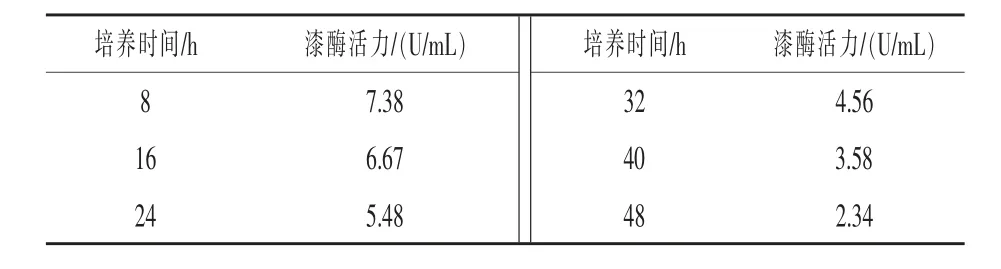
表5 不同培养时间对SDB2菌株漆酶活力的影响
进一步对漆酶活力与培养时间两者建立回归方程并做方差分析,结果如图4和表6所示。漆酶活力与培养时间存在极显著的线性回归关系(P<0.01),复相关指数R2=0.9957,漆酶活力随SDB2菌株培养时间增加而降低。
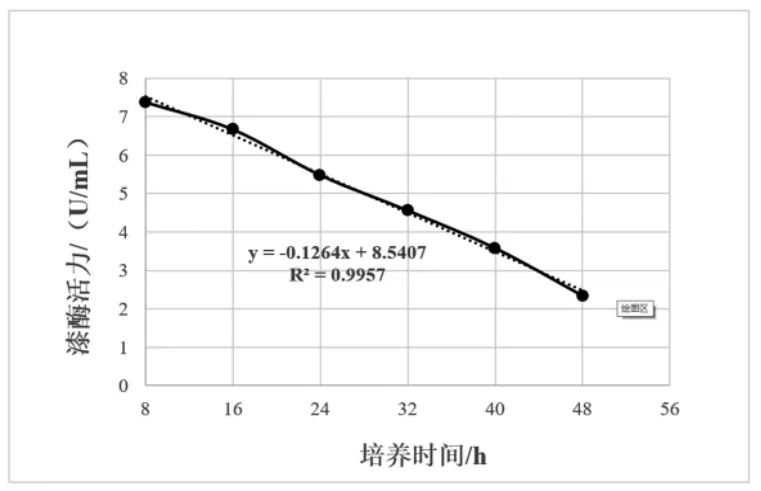
图4 SDB2菌株漆酶活力与培养时间的线性回归关系
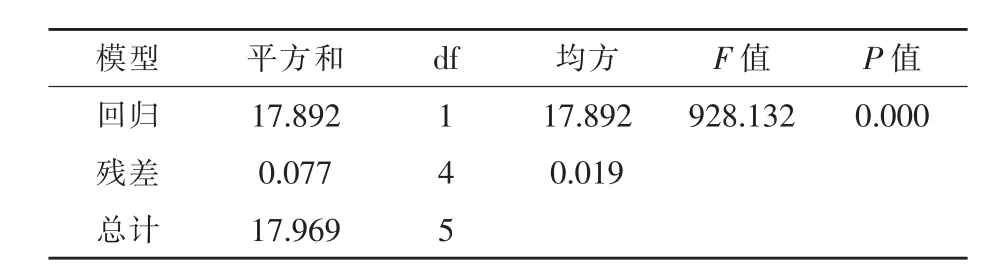
表6 SDB2菌株漆酶活力与培养时间建立回归方程的方差分析
对漆酶活力与芥子碱浓度两者建立回归方程并做方差分析,结果如图5和表7所示。漆酶活力与芥子碱浓度存在极显著的线性回归关系(P<0.01),复相关指数R2=0.9630,漆酶活力与芥子碱浓度呈正相关,芥子碱浓度高,则漆酶活力高,反之芥子碱浓度低,则漆酶活力低。

图5 漆酶活力与芥子碱浓度的线性回归关系
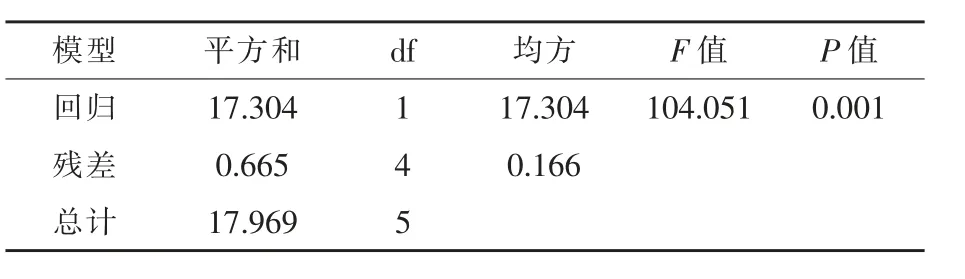
表7 漆酶活力与芥子碱浓度建立回归方程的方差分析
3 结论与讨论
本研究通过白芥子诱导试验,从鸡盲肠中筛选出一株能有效降解芥子碱的菌株SDB2,鉴定为大肠杆菌。将该菌株接种到芥子碱液体培养基中能有效降解芥子碱。芥子碱浓度与培养时间存在极显著的线性回归关系,芥子碱浓度随培养时间增加而降低,芥子碱在72 h内的降解率可达50%;SDB2功能酶漆酶活力与培养时间存在极显著的线性回归关系,漆酶活力随培养时间增加而降低;漆酶活力与芥子碱浓度存在极显著的线性回归关系,漆酶活力与芥子碱浓度呈正相关,芥子碱浓度高,则漆酶活力高,反之芥子碱浓度低,则漆酶活力低。
前人为降低菜籽粕、白芥子等十字花科类植物原料中芥子碱的含量也做了一些尝试,Lucht(1998)采用化学法使芥子碱含量降低至检测限以下,但菜籽粕中赖氨酸的含量下降了20%;Thiyam等(2006)用含水甲醇、乙醇等作为溶剂去提取芥子碱,存在溶剂损耗大、污染环境等问题,不利于生产应用;Mailer等(2009)和Mittasch等(2010)开展了利用转基因技术特异性地沉默芥子碱在植物中生物合成代谢,从而降低芥子碱含量的品种选育研究,但至今尚无商品化品种出现,并且此举会使芥子碱在十字花科类植物所发挥的生理代谢调控、提高植物抗病性和改善营养品质的重要功能丧失,无法统筹考虑芥子碱在生长植物中的营养功能角色和饲料原料中的抗营养因子角色。
采用生物技术方法降解芥子碱是目前最先进、最安全的技术手段之一,其原理是利用微生物自身分泌的功能性生物酶破坏芥子碱的分子结构,从而达到降解芥子碱的目的。芥子碱属于简单酚类物质,有关研究报道,多酚氧化酶、酪氨酸酶、阿魏酸酯酶、β-葡萄糖苷酶等能有效降解芥子碱(柯木根等,2007)。Lacki等(1996)发现,采用白腐真菌分泌的酶(多酚氧化酶为主)在水溶液中可酶解98%以上的芥子碱和芥子酸;钮琰星(2013)报道,蘑菇中提取的酪氨酸酶是一种一元酚氧化酶,能在30~40℃水解芥子碱,最终分解成苯醌或者其他衍生物,并在反应过程中发现有色物质的产生;Qiao等(2002)发现,阿魏酸酯酶可在50~60℃水解芥子碱,但不能继续分解其降解产物芥子酸;周浩宇等(2011)发现,β-葡萄糖苷酶对芥子碱具有一定的降解作用,降解率达17%。而漆酶作为一种多酚氧化酶,多存在于微生物中,其催化范围广,可催化氧化邻、对苯二酚等多元酚及其衍生物,催化氧化底物类型已达250个,但目前关于漆酶降解芥子碱的研究鲜见报道。Yu等(2016)发现,筛选出的芥子碱降解菌YD-1和YD-2的胞外产物含有蛋白酶、淀粉酶和脲酶;余丹等(2020)发现,筛选出的芥子碱降解菌SDB1具有漆酶、淀粉酶、多酚氧化酶、乙酰胆碱酯酶和脂肪酶的活性,不具有β-葡萄糖苷酶的活性。本研究采用鸡盲肠中筛选出的优势大肠杆菌SDB2去降解芥子碱,结果表明降解效果非常显著,同时也发现SDB2对芥子碱的降解与其所含漆酶的活性密切相关,漆酶活性与芥子碱浓度呈正相关,提示SDB2以芥子碱为营养物质分泌漆酶,芥子碱浓度高呈正营养状态时,则漆酶活性高,而随着漆酶对芥子碱的降解,引起芥子碱浓度降低呈现负营养状态,因此漆酶活性降低。余丹等(2020)筛选出的SDB1属于肺炎克雷伯氏菌属,该研究表明在培养液中分别添加0.40 g/L和0.80 g/L的芥子碱均可以提高SDB1漆酶活力,而本研究在培养过程中没有添加芥子碱作为营养补充,结合此前研究结果推测此为引起漆酶活力下降的原因,可为后续研究方法提供参考依据。
综上所述,为提高菜籽粕等非粮饲料资源的高效利用,本文采用安全有效的微生物发酵酶解技术去降解芥子碱等抗营养因子,在新冠疫情和中美贸易的双重压力下,具有十分重要的意义和广阔的应用前景。
猜你喜欢
现代畜牧科技(2021年5期)2021-07-20 08:07:48
临床与实验病理学杂志(2021年5期)2021-06-17 12:57:46
世界最新医学信息文摘(2020年87期)2020-12-10 04:47:46
现代检验医学杂志(2019年6期)2020-01-17 02:16:58
故事会(2019年1期)2019-01-11 01:18:42
新世纪智能(语文备考)(2018年11期)2018-12-29 12:30:56
宝藏(2017年2期)2017-03-20 13:16:28
小小说大世界(2016年2期)2016-03-07 12:15:11
动物营养学报(2015年9期)2016-01-07 11:29:38
中国组织化学与细胞化学杂志(2013年4期)2013-01-04 03:05:06
