
豆米轮作对春玉米土壤固氮细菌群落结构及多样性的影响
2023-02-14 07:00提俊阳张玉芹杨恒山张瑞富邰继承萨如拉韩镁琪
中国农学通报 2023年2期
提俊阳,张玉芹,杨恒山,张瑞富,邰继承,萨如拉,韩镁琪
(内蒙古民族大学农学院/内蒙古自治区饲用作物工程技术研究中心,内蒙古通辽 028000)
0 引言
内蒙古自治区是中国重要的粮食主产区,玉米作为主要粮食作物大面积连年种植,有些区域已经20年甚至30余年连作,种植结构单一、化肥过量与利用效率低。玉米替代种植可以调整农业生产结构,但由于过分强调比较效益,人们放弃了合理轮作体系,形成一种掠夺式的种植结构[1],玉米长期连作下导致土壤物理性状恶化[2]、有机质含量降低、同一耕层深度内的单一养分消耗较多,土壤养分失衡,出现缺肥和偏肥现象[3]。与此同时,土壤微生物的群落结构和多样性发生改变,导致玉米生理病害、微生物病害和营养失衡频繁发生[4-5],产量多年一直停滞甚至下降,影响粮食持续增产。因此,解决玉米连作障碍,提升土壤肥力已成为本地区农业可持续发展中亟待解决的重要问题。优化耕作制度以最大限度地发挥植物-土壤反馈的积极作用[6],科学合理的轮作模式可缓解连作引起的土壤环境恶化。研究表明,豆科与禾本科作物轮作中,禾本科作物吸收的氮量有5%~34%来自于前茬豆科作物根瘤所固定的氮素[7],其增产效果已得到广泛认可[8]。轮作能够影响土壤微生物群落生长代谢和群落结构[9-10],研究表明,玉米-玉米-大豆轮作能够显著提高土壤细菌的多个多样性指数[11];相较于玉米连作,玉米-大豆轮作能够提高总细菌和固氮菌的多样性并改变了细菌群落结构[12]。在内蒙古大豆主产区建立了玉米-大豆轮作的种植模式[1],其中呼伦贝尔市大力推行大豆-玉米、大豆-玉米-玉米轮作模式。前人从产量和效益方面也做了研究[13-14],发现在旱涝年份,轮作对产量影响效果大于常年[13]。免耕秸秆覆盖结合燕麦-大豆-玉米轮作模式增产效果较好[15];在旱作区,通过实施多年免耕轮作可实现作物的增产增收[16]。前人对内蒙古作物轮作农田系统的研究主要集中在轮作周期土壤物理化学质量,土壤微生物生物量碳、氮、磷含量及土壤酶活性的季节变化、年际变化和空间变异规律的分析方面;针对大豆玉米轮作模式下土壤固氮细菌群落结构及多样性的变化未见报道。本研究以连作玉米为对照,以nif-H基因为固氮菌的指示基因,利用高通量测序技术,分析大豆-玉米和大豆-玉米-玉米2种轮作模式下固氮细菌群落结构及多样性的变化,并分析其与土壤有机质和氮素养分之间的关系,明确不同轮作方式对土壤养分及固氮细菌的影响,以期为呼伦贝尔市及同类地区合理的推行米豆轮作提供理论依据。
1 材料与方法
1.1 试验区概况
试验于2018—2020年在内蒙古呼伦贝尔阿荣旗农业科技园区(47°56′54″—49°19′35″N,122°02′30″—124°05′40″E)进行。当地属温带大陆性半湿润气候,年平均降水量458.4 mm,年有效积温2394.1℃,无霜期90~130天。土壤类型为黑钙土,有机质31.51 g/kg,全氮1.84 g/kg,有效磷8.97 mg/kg,速效钾67.66 mg/kg。
1.2 试验设计
以玉米连作为对照(MMM),设大豆-玉米2年轮作周期(MSM)和大豆-玉米-玉米3年轮作周期(SMM)2个处理。大豆选用高产、宜机收品种‘蒙豆30’,采用垄上双行的种植方式,行距65 cm,株距10~15 cm,种植密度37.5万株/hm2,底施复合肥(26-15-10)315 kg/hm2;玉米品种选用‘A6565’,采用等行距65 cm种植,种植密度75000株/hm2,底施复合肥(26-15-10)450 kg/hm2,于玉米大喇叭口期追施尿素(46-0-0)150 kg/hm2;玉米、大豆均为4月下旬播种,10月初收获。每处理3次重复,小区面积5 m×8 m=40 m2。
1.3 土壤样品采集
于2020年(8月2日)春玉米吐丝期,各小区分别取0~20 cm和20~40 cm土层土壤,每个小区按S型分布取5个样点,将每小区相同土层5个样点的土混合成一个样品,除去碎石、根系及其他杂物后,充分混合后分成3份,1份装于已灭菌的5个10 mL试管并置于冰盒中,迅速带回实验室,保存于-80℃冰箱中,用于nif-H基因测序;第2份装于自封袋于-20℃冷冻保存,用于测定铵态氮和硝态氮;第3份风干后用于测定土壤有机质、全氮和碱解氮含量。具体土样名称编号见表1。

表1 不同处理不同土层土样编号
1.4 土壤养分测定
有机质采用重铬酸钾氧化-比色法(A590 nm)测定;土壤全氮采用H2SO4-H2O2消煮后用全自动离子分析仪(easychem,意大利SYSTEA)测定;土壤碱解氮采用碱扩散法测定;土壤铵态氮和硝态氮采用2 mol/L KCl溶液浸提后,用全自动离子分析仪(easychem,意大利SYSTEA)测定。
1.5 土壤DNA提取和固氮细菌nif-H基因PCR扩增
土壤样品送至北京奥维森生物技术公司进行nif-H基因提取及PCR扩增样品测序,每个试验样品取0.3 g土样,采用E.Z.N.A.Soil DNA Kit(Omega,CA,USA)试剂盒提取土壤DNA,选用正向引物(5′-AAA GGY GGWATC GGYAAR TCC ACC AC-3′)和反向引物(5′-TTG TTS GCS GCR TACATS GCCATCAT-3′)扩增固氮细菌nif-H基因。PCR反应体系包含XμL(30 ng)DNA样品、1 μL正向引物(5 uM)、1 μL反向引物(5 uM)、3 μL BSA(2 ng/μL)、12.5 μL 2xTaq Plus Master Mix、7.5-X μL dd H2O,总计25 μL;反应条件为94℃预变性5 min,94℃变性30 s,58℃退火30 s,72℃延伸60 s,35个循环,72℃延伸7 min,置于4℃冰箱内保存。PCR产物使用1%琼脂糖凝胶电泳检测扩增目的条带大小,并用Agencourt AMPure XP核酸纯化试剂盒纯化,使用Illumina Miseq高通量测序平台上机测序。
1.6 PCR扩增产物的质量控制
对扩增固氮细菌nif-H基因进行双端测序,截去条形码和引物序列后使用FLASH(V1.2.7)对每个样品的读长(reads)进行拼接;通过QIIME(V 1.7.0)过滤掉低质量的拼接序列,并使用UCHIME算法去除嵌合体;将得到的有效序列通过Uparse C(V7.0.1001)在97%的相似性水平上进行OTUs聚类,对OTUs代表序列进行物种注释。
1.7 数据分析
利用Excel 2019、DPS(V9.5)对不同处理不同土层间土壤养分、在97%一致性阈值下的α样性指数(Chao1、ACE、Shannon、Simpson)进行多重比较(LSD法,P<0.05为具有显著性差异),并用R软件(V2.15.3)对不同菌门、属绘制稀释曲线、柱状图。
2 结果与分析
2.1 不同土层土壤养分含量
如表2所示,与MMM相比,两种轮作模式0~20、20~40 cm土层土壤有机质、全氮、碱解氮、铵态氮和硝态氮均显著增加;MSM土壤有机质0~20 cm土层显著高于SMM,碱解氮和硝态氮2个土层均显著高于SMM,全氮和氨态氮差异不显著。说明轮作可增加土壤有机质和氮素养分含量。

表2 不同轮作模式土壤样品养分含量
2.2 测序结果质量分析
使用IlluminaMiseq技术对固氮微生物nif-H基因测序分析,经进一步去除嵌合体、短序列等,6个样品共得到原始序列256133条,经进一步去除嵌合体、短序列后得到优质序列244033条。如图1所示,序列数量达到20000时稀释性曲线均趋于平坦,说明此测序深度获得序列数据量可以反映土壤样品固氮微生物信息。
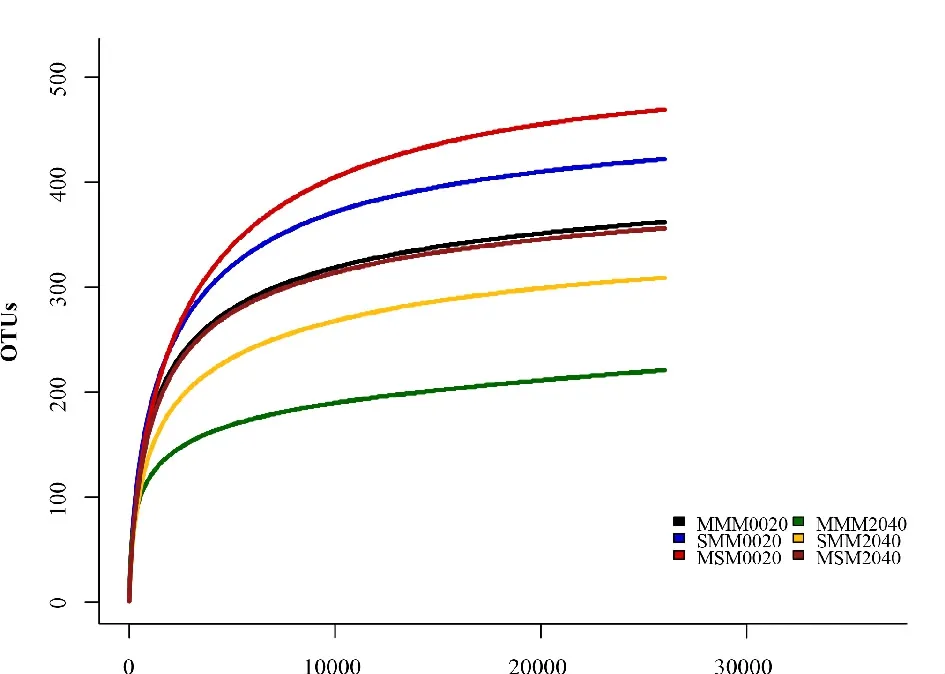
图1 不同轮作模式土壤样品稀释曲线
2.3 不同轮作模式固氮细菌的多样性
从表3可知,相较于MMM、MSM和SMM固氮细菌Chao1指数和ACE指数在0~20 cm土层和20~40 cm土层均显著增加,轮作1年周期MSM高于2年周期SMM;Shannon多样性和Simpson多样性在0~20 cm土层MSM与SMM间差异不显著,但均显著高于MMM,20~40 cm土层MSM显著高于MMM。
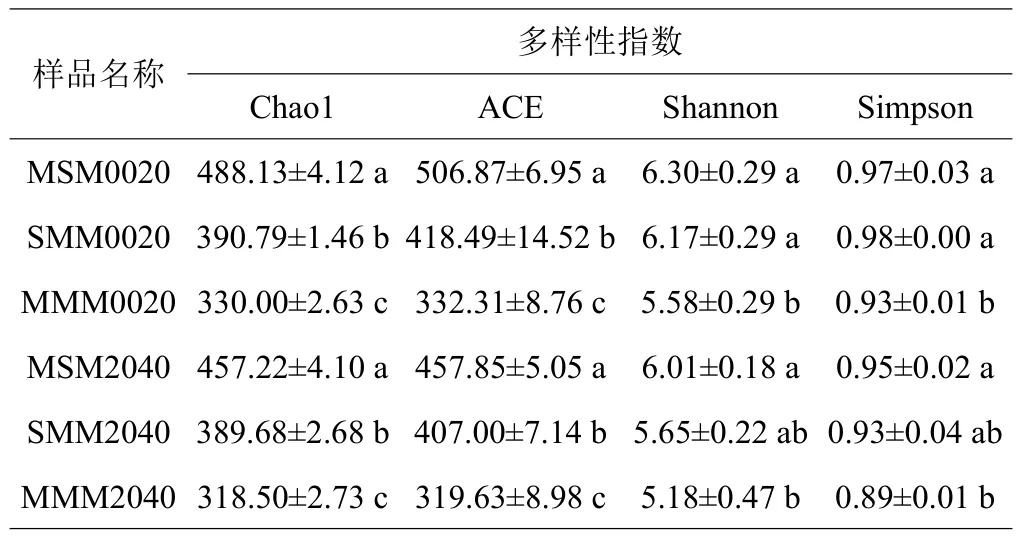
表3 固氮细菌多样性指数
2.4 土壤固氮细菌群落结构分析
通过对样品获得的OTUs进行归类,得到15个门、23个纲、56个目、89个科和163个属。由图2可知,MMM、MSM、SMM各组样品优势菌门主要分布在变形菌门(Proteobacteria)、疣微菌门(Verrucomicrobia)和放线菌门(Actinobacteria),这3个门的OTUs丰度占所有固氮菌门的87.68%~91.5%。其中,他们的丰度分别为81.59%~88.0%、0.2%~5.2%和1.13%~2.9%。MSM和SMM的0~20 cm和20~40 cm土层变形菌门相对丰度相较于MMM均有所提高(0.17%、0.17%;3.57%、1.06%),MSM较SMM变形菌门0~20 cm土层相对丰度没有显著差异,20~40 cm土层提高2.5%。
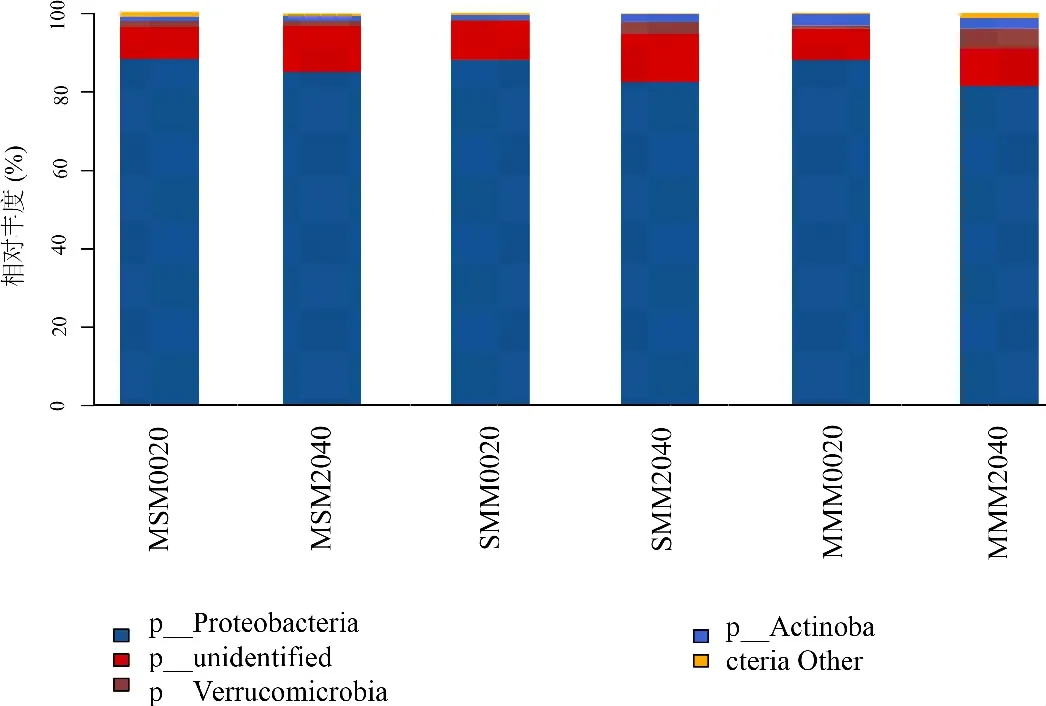
图2 不同轮作方式土壤在门分类水平上nif-H基因的分布
从图3可知,各组样品优势菌属包括12个属,他们分别是慢生根瘤菌属(Bradyrhizobium)、斯科曼氏球菌属(Skermanella)、伪食酸菌属(Pseudacidovorax)、地杆菌属(Geobacter)、厌氧粘杆菌属(Anaeromyxobacter)、固氮螺菌属(Azospirillum)、甲基孢囊菌属(Methylocystis)、脱硫菌属(Desulfarculus)、脱氯单胞菌属(Dechloromonas)、弗兰克氏菌属(Frankia)、固氮弧菌属(Azoarcus)和纤毛菌属(Leptothrix),占所有菌属的73.39%~82.21%。与MMM相比,MSM和SMM的0~20 cm土层慢生根瘤菌属(8.18%、6.36%)、未知菌属(2.87%、4.72%)、弗兰克氏菌属(0.02%、0.07%)和纤毛菌属(0.02%、1.30%)相对丰度均有所提高;20~40 cm土层慢生根瘤菌属(10.24%、9.73%)、伪食酸菌属(5.64%、2.32%)、地杆菌属(0.66%、1.74%)和固氮弧菌属(2.08%、0.38%)相对丰度均有所提高。MSM与SMM相比,0~20、20~40 cm土层慢生根瘤菌属(1.83%、0.52%)、甲基孢囊菌属(4.76%、2.84%)和固氮弧菌属(0.46%、1.70%)均有所提高。
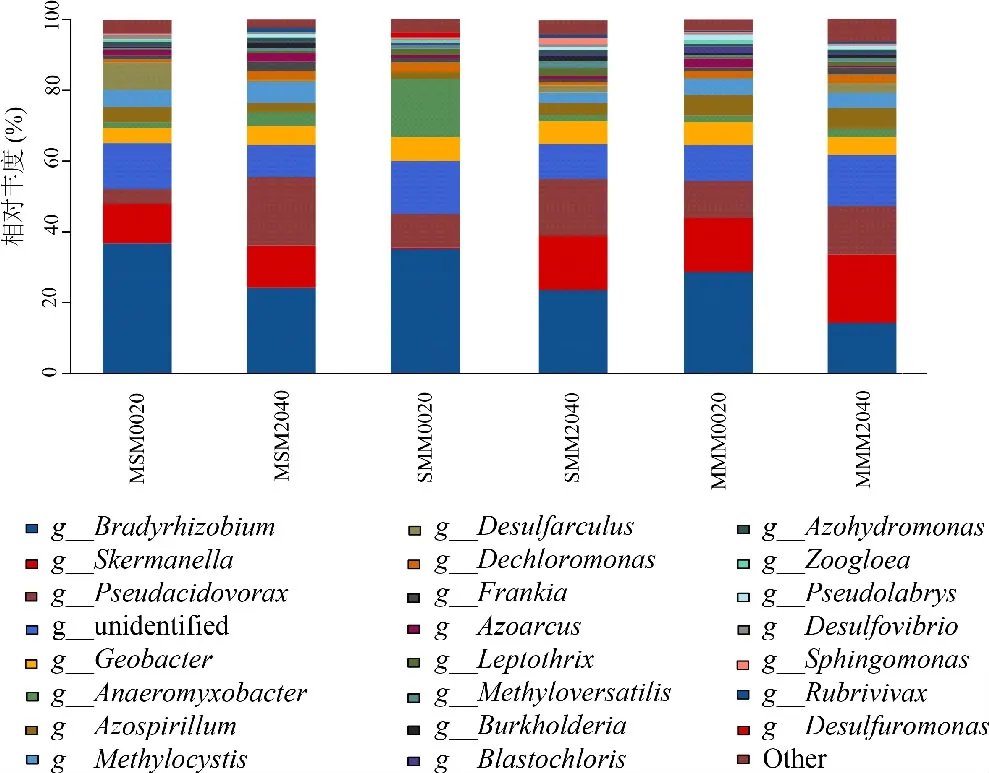
图3 不同轮作方式土壤在属分类水平上nif H基因的分布
2.5 土壤固氮细菌多样性指数和优势菌属与土壤养分的相关性分析
从表4可知,Chao1、ACE指数与各养分指标均呈正相关,OM达到显著水平,AN、NO3--N达到极显著水平;Shannon、Simpson指数与AN、NO3--N呈正相关,与OM、TN、NH4+-N呈负相关。

表4 土壤固氮细菌多样性指数与土壤养分的相关性分析
从表5所示,慢生根瘤菌属(Bradyrhizobium)与各养分指标呈正相关,OM、TN、NH4+-N达到极显著水平;斯科曼氏球菌属(Skermanella)与各养分指标呈负相关;伪食酸菌属(Pseudacidovorax)与各养分指标呈负相关,TN、NH4+-N达到极显著水平;地杆菌属(Geobacter)与NH4+-N呈正相关,与其他养分指标呈负相关;厌氧粘杆菌属(Anaeromyxobacter)与各养分指标呈正相关;固氮螺菌属(Azospirillum)与各养分指标呈负相关;甲基孢囊菌属(Methylocystis)与OM、TN、NH4+-N呈负相关,与AN、NO3--N呈正相关;脱硫菌属(Desulfarculus)与各养分指标呈正相关;脱氯单胞菌属(Dechloromonas)与各养分指标呈负相关;弗兰克氏菌属(Frankia)与AN呈正相关,与其他各养分指标呈负相关;固氮弧菌属(Azoarcus)与各养分指标呈正相关;纤毛菌属(Leptothrix)与各养分指标呈负相关。

表5 土壤固氮细菌优势菌属与土壤养分的相关性分析
3 结论
相较于玉米连作,大豆-玉米-玉米轮作和大豆-玉米轮作明显提高土壤养分和有机质含量,增加了土壤固氮细菌丰度和多样性,提高了变形菌门和慢生根瘤菌属的相对丰度。2种轮作模式下,固氮细菌大豆-玉米2年轮作周期下丰度较高。固氮细菌丰度指数与有机质、碱解氮和硝态氮呈显著正相关,有机质、全氮和铵态氮对优势菌属影响较大。
4 讨论
4.1 大豆-玉米轮作对玉米土壤固氮细菌丰度及多样性的影响
土壤养分的差异是影响土壤微生物群落丰度和多样性的主要原因之一[17]。作物轮作多样性和作物残茬影响作物活力和相关微生物[18],大豆玉米轮作模式通过轮换种植不同作物,不同作物对土壤养分偏耗不同,不同作物残茬改变了土壤养分,对土壤微生物丰度和多样性影响显著[19]。与营养元素相比,微生物组更好地预测作物生物量和产量[18];本研究发现,大豆-玉米和大豆-玉米-玉米轮作模式下固氮细菌的多样性和丰度均改变,0~20 cm土层MSM和SMM丰度指标Chao1和ACE与多样性指标Shannon和Simpson均显著高于MMM;2种轮作模式相比较,MSM丰度指标Chao1和ACE显著高于SMM。作物影响土壤生物群落的主要手段是通过有机物的输入,大豆和固氮细菌之间的相互作用,促进土壤碳储存[6];土壤碳和氮等养分是土壤固氮细菌丰度及群落改变的主要影响因素[20],固氮细菌所需能量主要源于土壤有机质,MSM土壤固氮细菌丰度显著高于SMM,这可能由于MSM的有机质养分含量高,为固氮菌生长提供较多的碳源;而SMM和MMM土壤养分含量低,限制了固氮细菌的生长。此外,固氮细菌多样性与作物残茬和根系分泌物关系密切[21],根系分泌物显著增加了土壤中与氮循环生物过程相关的基因,如固氮等[22],MSM土壤固氮细菌的丰富度和多样性较高,可能由于根系分泌物和残茬较多,根系分泌物诱导了更高的养分矿化,以帮助植物调节微生物的募集,以满足更快生长时期对养分的需求[22]。同时,后茬作物玉米轮作两年实际上又造成了禾本科作物的连作也是其原因之一。相关性分析表明,有机质、碱解氮、硝态氮对固氮细菌丰度指数的提高影响显著,Chao1、ACE指数与有机质、碱解氮、硝态氮达到显著正相关。HU等[23]研究表明,土壤pH、全氮和硝态氮是调控固氮菌网络的主要因子。因此,土壤养分可利用性提高,有利于固氮细菌生长繁殖,土壤微生物多样性升高[24]。
4.2 大豆-玉米轮作对固氮细菌群落结构的影响
作物轮作能使土壤中聚集多种作物残茬和作物根系分泌物,使土壤内微生物生长和繁殖所需的营养成分均衡,从而改变土壤微生物的群落结构。研究表明,不同轮作模式下土壤固氮细菌的群落结构有所不同[25-26],轮作方式、种植年限等因素会导致研究结果产生差异[27-28]。本研究中,2种轮作模式土壤固氮优势门和属类群相似,以变形菌门和慢生根瘤菌属为主,相较于MMM、MSM和SMM 2个土层的变形菌门相对丰度较高,慢生根瘤菌属也较高,两种轮作方式下,MSM高于SMM。豆科作物与禾本科作物轮作可以提高土壤有机碳含量[29],MSM土壤有机质含量高和玉米生长旺盛根系分泌物较多,它们共同作用可能促进变形菌门菌群生长,导致变形菌门相对丰度显著高于SMM和MMM。HU等[23]研究了连续35年单施或配施含氮、磷或钾的无机肥对东北黑土固氮微生物群落丰度和组成的影响,发现施氮肥处理固氮菌群落以α-变形菌纲(属水平上的慢生根瘤菌)为主;本研究中慢生根瘤菌属相对丰度MSM最高,可能由于其前茬种植大豆,慢生根瘤菌属是大豆进行根瘤固氮的主要菌属[30],而大豆残茬的存在导致次年土壤内慢生根瘤菌属相对丰度较高。最近的研究表明凋落物的输入类型可以强烈地影响土壤生物群落分解有机化合物的能力,并导致微生物群落向特定凋落物类型分化[6];土壤有机物质输入的类型、速率和时间是土壤微生物群落的重要驱动力[6];大豆玉米轮作农田系统中作物秸秆类型多样化,大豆和玉米秸秆分解速率不同,还入土壤的时间不同,这可能是轮作微生物多样性不同于玉米连作的原因。
植物根系的位置是控制细菌群落的主要因素,其次是发育阶段和土壤类型[31];本研究中MSM和SMM对不同土层固氮菌的影响有所不同,0~20 cm土层弗兰克氏菌属相对丰度有所提高,20~40 cm土层中均提高了伪食酸菌属和固氮弧菌属相对丰度。弗兰克氏菌是一种能与非豆科植物共生形成根瘤并实现生物固氮的放线菌[32],其固氮效率远远高于豆科植物根瘤菌,还能够促进宿主植物对旱寒等各种不同生境的适应性[33],在重金属污染土壤复垦中弗兰克氏菌与其宿主的共生过程中发挥关键作用[34-35];伪食酸菌具有固氮作用[36]。因此,大豆玉米轮作增加了土壤中固氮效率高多功能的有益微生物类群。研究表明,变形菌门的细菌大多属于异养型微生物,其中包含许多与碳循环有关的细菌,可用来指示土壤肥力状况[37];慢生根瘤菌在多种环境中普遍存在,土壤中处于优势地位[38],慢生根瘤菌与丛枝菌根真菌协同作用显著提高了结瘤、固氮、营养、种子产量和生物量[39];本研究中,优势菌属与养分的相关性分析表明,慢生根瘤菌属、厌氧粘杆菌属、脱硫菌属、固氮弧菌属与有机质、全氮、碱解氮、铵态氮和硝态氮呈正相关,其中慢生根瘤菌属与有机质、全氮、铵态氮呈极显著正相关,说明微生物与土壤养分之间的正相关关系表明豆米轮作土壤碳、氮养分的增加仅对共生固氮菌慢生根瘤菌属有促进作用。轮作条件下固氮细菌和参与碳循环细菌之间的耦合关系值得进一步研究。
猜你喜欢
今日农业(2021年11期)2021-11-27
生态科学(2021年5期)2021-11-13
园艺与种苗(2021年1期)2021-03-16
西藏农业科技(2019年3期)2019-11-04
中国化肥信息(2018年12期)2018-03-01
新农业(2017年24期)2018-01-30
——固氮微生物
新疆农垦科技(2016年3期)2016-02-20
江西农业大学学报(2013年3期)2013-11-05
植物营养与肥料学报(2012年5期)2012-10-26
植物营养与肥料学报(2012年1期)2012-10-26
