
物理处理对黄粉虫蛋白理化及消化性质的影响
2023-02-13 07:25王金梅江轶群穆利霞骆兆娇孟赫诚
现代食品科技 2023年1期
王金梅,江轶群,穆利霞,骆兆娇,孟赫诚*
(1.华南理工大学食品科学与工程学院,广东广州 510640)(2.广东省农业科学院蚕业与农产品加工研究所,广东省农产品加工重点实验室,农业农村部功能食品重点实验室,广东广州 510610)
黄粉虫(Tenebrio molitor)是粉虱的幼虫,蛋白含量高(48%~66%,干基),且富含必须氨基酸,是一种良好的可食性昆虫蛋白资源[1]。黄粉虫蛋白(TPI)常采用碱溶酸沉法进行提取,目前的研究大多集中于提取工艺的优化,评价黄粉虫蛋白的乳化性和起泡性等功能性质,以期为黄粉虫蛋白应用于食品体系提供理论依据。Jiang等[2]采用盐溶/盐析辅助碱溶酸沉法提取TPI,发现(NH4)2SO4盐析处理可显著提高蛋白的溶解性、乳化活性和泡沫稳定性。Zhao等[3]报道两次碱提可有效提高TPI提取率和蛋白含量,并发现提取过程中物料比、温度、盐浓度、pH等因素均影响了TPI的流变学特性。然而,关于不同加工处理对TPI结构和功能性质影响的研究仍较为有限,且未见关于 TPI体外消化性的研究,限制了TPI在食品工业的应用。
热处理是食品工业中常见的操作单元,如热杀菌等。热处理可诱导蛋白质去折叠,内部疏水基团暴露并发生热聚集,影响其在界面上的吸附和界面膜的形成,从而影响其界面性质[4]。对大豆分离蛋白进行热处理可获得具有高表面疏水性的柔性的蛋白结构,显著提高其在油水和气水界面的表面活性,改善起泡性和泡沫稳定性[5]。Lee等[6]发现随着加热温度和加热时间的提高,TPI的表面疏水性和聚集程度不断提高,乳化性能和起泡性降低但凝胶强度增强。热处理对蛋白体外消化性的影响与蛋白种类和热处理强度密切相关。Ren等[7]认为热处理不影响大豆分离蛋白在体外模拟胃中的水解,但显著改善了其在体外模拟肠道中的水解速率,且高温处理(121 ℃)的蛋白去折叠程度更高,暴露了更多的酶结合位点,故其消化性的改善效果明显高于巴氏杀菌(70 ℃)处理的样品。Liu等[8]则认为热处理促进了大米蛋白分子间二硫键的形成和疏水相互作用,提高了其抗消化性。
动态高压微射流是一种新型均质技术,流体在高压状态下经受高强剪切力、冲击力和空穴效应,可能改变大分子的结构及其与水分子间的相互作用,进而影响蛋白的功能性和消化性[9]。微射流处理(120 MPa)下,大豆分离蛋白发生部分去折叠和结构重排,乳化活性和乳化稳定性得以改善[10]。微射流处理对蛋白聚集体的解聚作用也可用以提升难溶蛋白的溶解度[11]。Chen等[12]认为微射流处理可改变β-乳球蛋白的构象和聚集状态,从而改善其胃肠道消化性。本文以碱溶酸沉法提取的TPI为原料,探究了热处理和动态高压微射流处理对TPI的理化、功能和体外消化性质的影响,为昆虫蛋白作为食品蛋白资源应用于蛋白饮料、乳液、泡沫食品的可行性提供理论依据。
1 材料与方法
1.1 原料
脱脂黄粉虫(Dumas法测定其蛋白含量为73.30%±0.15%,N×5.71,干基),广东泽和诚生物科技有限公司,粉碎60目备用。福林酚试剂盒购于北京鼎国公司。SDS-PAGE电泳试剂盒购于北京索莱宝公司。胃蛋白酶(效价1:10 000)和胰蛋白酶(效价1:250)购于美国Sigma公司,猪胆盐提取物购于北京索莱宝公司。其他化学试剂均为分析纯。
1.2 主要仪器设备
DELTA冷冻干燥机,德国 Christ公司;Dumas定氮仪,德国Elementer公司;Zeta电位及纳米粒度分析,英国Malvern公司;UVA2300紫外可见分光光度计,德国Christ公司;RW20电动搅拌机,德国IKA公司;PYCZ-30垂直板电泳仪,北京六一仪器厂。
1.3 试验方法
1.3.1 黄粉虫蛋白的制备
将黄粉虫以1:20的料液比加入去离子水,调pH值12.0,充分搅拌2 h后8 000 r/min离心20 min,上清液调pH值4.5后8 000 r/min离心20 min,蛋白沉淀水洗、复溶,调pH值7.0,4 ℃透析48 h后冷冻干燥,制得天然黄粉虫蛋白NTPI。
热处理:将NTPI溶于10 mmol/L磷酸缓冲液(pH值7.0),配置成1%(m/m)的NTPI溶液,室温搅拌2 h后密封于带盖瓶分别在90 ℃和120 ℃进行热处理,处理时间均为15 min。其中,90 ℃热处理采用水浴方式,120 ℃热处理采用高压蒸汽灭菌方式进行。热处理后,样品迅速冰浴冷却制得HTPI-90和HTPI-120。
动态高压微射流处理:将NTPI溶于10 mmol/L磷酸缓冲液(pH值7.0),配置成1%(m/m)的NTPI溶液,室温搅拌2 h后分别在50 MPa和100 MPa的压力下经动态高压微射流循环处理三次,制得DTPI-50和DTPI-100。
1.3.2 天然黄粉虫蛋白在不同溶液中溶解度
将NTPI溶于10 mmol/L磷酸缓冲液(pH值7.0),配置成1 mg/mL的NTPI溶液,分别加入10 mmol/L DTT、8 mol/L尿素、m=1% SDS、m=1% SDS+8 mol/L尿素、m=1% SDS+8 mol/L尿素+10 mmol/L DTT。水化过夜后8 000 r/min离心20 min,采用Lowry法[13]测定上清液中的蛋白含量。以牛血清白蛋白为标准蛋白制作标曲,上清液的蛋白总含量与样品蛋白总含量的比值即为溶解度。
1.3.3 十二烷基硫酸钠聚丙烯酰胺凝胶电泳(SDS-PAGE)
电泳分析参考Laemmli[14]的方法进行。分别向天然和物理处理后的TPI溶液中加入还原型上样缓冲液[0.125 mol/L Tris-HCl缓冲液,包含1% SDS(m/V)、2%β-巯基乙醇(2-ME,V/V)、5%甘油(V/V)和0.025%溴酚蓝(m/V)]后涡旋,热水煮沸5 min,10 000 r/min离心5 min。配制浓度为12.5%的分离胶与5%的浓缩胶,取10 μL样品上清上样,在恒压条件下进行电泳。其中样品在浓缩胶中时电压为40 V,进入分离胶后将电压调至120 V。用考马斯亮蓝R250配制的染色液对胶片染色 30~45 min,然后用脱色液(乙醇:乙酸:水=100:100:800,V/V/V)对胶片进行脱色。
1.3.4 巯基和二硫键
游离巯基依据 Beveridge等[15]方法测定,分别向2 mL天然和物理处理后的蛋白溶液加入 10 mL Tris-Gly-8M Urea缓冲液,8 000 r/min离心20 min,取上清进行检测。游离巯基含量(SHF,μmol/g protein):取2 mL上清液,加入80 μL Ellman's试剂(4 mg/mL DTNB),立即混匀,静置15 min后在412 nm波长下测定吸光值(A1)。总巯基含量(SHT,μmol/g protein):取2 mL上清液,加入0.2%β-巯基乙醇处理2 h后,再加入4 mL TCA溶液(12%,m/V),8 000 r/min离心20 min,洗涤沉淀后复溶,取2 mL测吸光值(A2)。巯基及二硫键(SS,μmol/g protein)的计算公式如下:
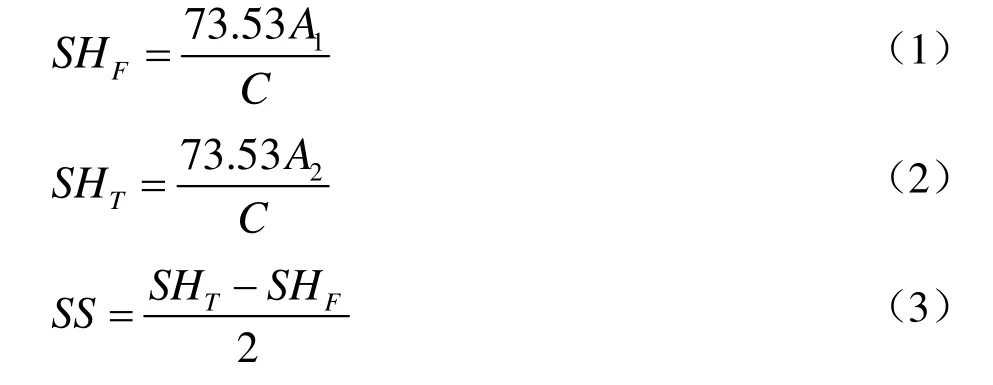
式中:
73.53=106/(1.36×104),1.36×104为 Ellman's试剂的摩尔消光系数;
C——蛋白浓度,mg/mL。
1.3.5 黄粉虫蛋白在不同pH值下的溶解度
天然和物理处理后的TPI溶液分别调pH值至2.0、3.0、4.0、5.0、6.0、7.0、8.0 和9.0。8 000 r/min 离心20 min后,采用Lowry法[13]测定上清液中的蛋白含量。以牛血清白蛋白为标准蛋白制作标曲,上清液的蛋白总含量与样品蛋白总含量的比值即为溶解度。
1.3.6 乳化性
采用经典的比浊法[16],以乳化活性指数(Emulsifying Activity Index,EAI,单位m2/g)和乳化稳定指数(Emulsifying Stability Index,ESI,单位min)表征TPI的乳化性。将TPI充分溶解于磷酸缓冲液(pH值7.0)中,与大豆油以4:1的体积比混合,15 000 r/min均质1 min,迅速从乳液底部吸取50 μL加入5 mL SDS溶液(0.1%,m/V)中并涡旋,在500 nm下测其吸光值,记为A0;10 min后再次吸取50 μL加入5 mL SDS溶液(0.1%,m/V)中,涡旋后测其吸光值,记为A10,以0.1%的SDS溶液作为空白对照。
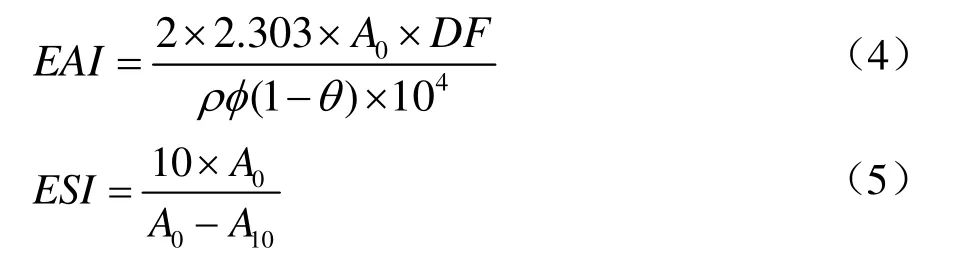
式中:
DF——稀释因子;
ρ——蛋白浓度,g/mL;
φ——光程,φ=0.01;
θ——油相的体积分数,θ=0.25。
1.3.7 起泡性
以起泡能力(Foaming Capacity,FC)和泡沫稳定性(Foaming Stability,FS)表征 TPI的起泡性。将10 mL TPI溶液(1%,m/V)用均质机以10 000 r/min的速率剪切2 min后迅速转入50 mL量筒中记录搅打后样品体积,记为V1;静置30 min后再次读取样品体积,记为V2。
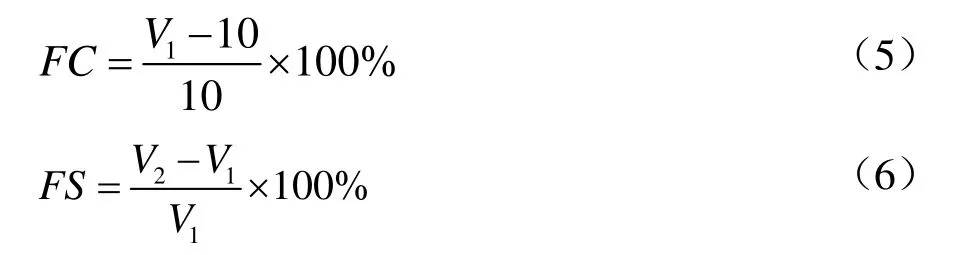
1.3.8 体外消化性
TPI的体外消化实验根据 Nunes[17]等的体外消化模型进行。调节TPI溶液(1%,m/V)的pH值至2.0,于37 ℃摇床中预热搅拌10 min后加入胃蛋白酶进行酶解,酶解1 h后调节pH值至7.0以终止模拟胃消化过程。向消化液中加入猪胆盐提取物和胰蛋白酶酶解,酶解2 h后对消化液进行加热处理以终止模拟肠消化。期间分别于0、10、30、60、90、120、150和180 min取样,同时对体外消化过程中的亚基降解情况进行SDS-PAGE的分析,具体方法见 1.3.3,并采用TCA-NSI法测定氮释放量。
1.3.9 数据分析
实验数据为三次测定平均值,结果以“平均值(mean)±标准偏差(SD)”表示。组间的显著性差异和相关性分析采用SPSS 19.0软件的ANOVA分析方法,p<0.05为显著性差异。
2 结果与讨论
2.1 天然黄粉虫蛋白在不同溶液中的溶解度
天然黄粉虫蛋白在不同溶液中的溶解度见图1。由图可知,NTPI在pH值7.0的磷酸缓冲液中溶解度较低(22.49%)。加入可裂解蛋白质分子间二硫键的强还原剂DTT后,其溶解度略有提升,说明二硫键参与了TPI高级结构的形成。尿素和SDS的添加使NTPI溶解度分别显著提升至75.41%和96.20%,表明氢键和疏水相互作用是NTPI高级结构形成过程中的主要作用力。当以上三种试剂全部添加时,NTPI可完全溶解。二硫键、氢键及疏水相互作用均参与TPI高级结构的形成,其中,疏水相互作用在稳定TPI结构中占主导地位。
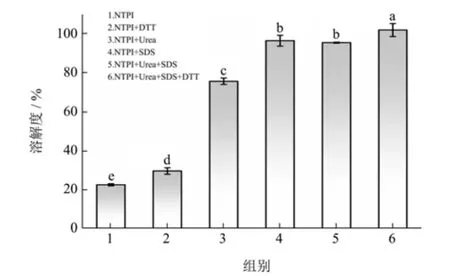
图1 天然黄粉虫蛋白在不同溶液中的溶解度Fig.1 Solubility of NTPI in different solutions
2.2 物理处理对黄粉虫蛋白理化性质的影响
天然和物理处理后黄粉虫蛋白的SDS-PAGE图谱见图2。由图可知,NTPI在分子量75 ku附近出现两条清晰的蛋白条带,可能与黄粉虫中调控脂肪形成的蛋白有关[18]。在43 ku和59 ku附近出现的条带可能来自昆虫自身的黑化抑制蛋白(43 ku)[19]、胰蛋白酶(59 ku)[20]和β-糖苷酶(59 ku)[21]。在14~32 ku之间出现的蛋白条带可能来源于黄粉虫的表皮蛋白[22]。在分子量小于14 ku范围内的条带可能来自黄粉虫的抗冻蛋白[1]。

图2 物理处理后黄粉虫蛋白的SDS-PAGE图谱Fig.2 SDS-PAGE profiles of TPI after physical treatments
中性条件下的热处理可能导致蛋白结构的展开和聚集行为的发生[4]。HTPI-90在分离胶顶端颜色加深,说明在90 ℃热处理形成了聚集体,这可能是由于TPI发生了部分去折叠,内部疏水基团暴露,发生疏水性聚集。而当加热温度提升至120 ℃时,HTPI-120分离胶中大量特征条带明显变浅甚至完全消失,而在点样孔处和分离胶顶端均出现深色条带,意味着120 ℃热处理诱导TPI亚基发生了更强烈的聚集。Lee等[6]以荧光法监测热处理过程中TPI的疏水性聚集,发现TPI的聚集程度随着温度的升高而提高。DTPI-50与NTPI的SDS-PAGE图谱基本一致,DTPI-100在点样孔处出现颜色较浅的条带,说明说明动态高压微射流处理并不影响TPI的亚基组成,100 MPa处理可形成少量TPI聚集体。Fan等[11]采用高压微射流在40~120 MPa下处理燕窝蛋白,其SDS-PAGE图谱与对照样品差异较小,说明微射流处理不改变蛋白的初级结构。
游离巯基是影响蛋白质界面和乳化行为的重要参数之一,见图3。NTPI的游离巯基和总巯基含量分别8.58 μmol/g 和 31.66 μmol/g,说明 NTPI中的大部分巯基均以二硫键形式存在。90 ℃热处理后,HTPI-90的游离巯基含量略有下降(7.99 μmol/g),二硫键含量略有提升。蛋白质在热处理时二硫键可断裂生成游离巯基,冷却过程中游离巯基重排可形成二硫键,巯基-二硫键的互换反应导致了HTPI-90游离巯基含量的变化。当热处理温度提升至120 ℃时,HTPI-120游离巯基含量提升8.41 μmol/g,二硫键含量从11.77 μmol/g降低至11.17 μmol/g。结合图2可知,HTPI-120发生了强烈的疏水聚集,在聚集过程中游离巯基可能被包埋入聚集体内部,无法与氧气或其他游离巯基接触,从而抑制了二硫键的生成。
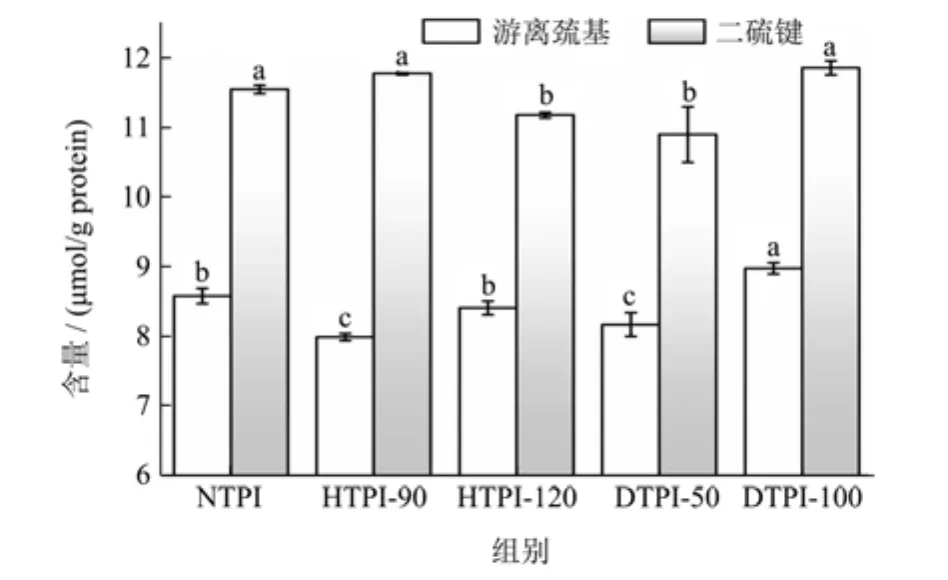
图3 物理处理对TPI游离巯基及二硫键含量的影响Fig.3 Effects of physical treatments on contents of free sulfhydryl group and disulfide bond in TPI
动态高压微射流作为一种高压剪切方式,可显著影响蛋白的高级结构,导致蛋白构象的改变,蛋白游离巯基的变化为其表现形式之一,且这些变化与压力水平并不一定具有直接相关性[9]。50 MPa动态微射流处理后,DTPI-50的游离巯基和二硫键含量均有所下降(8.17 μmol/g和 10.89 μmol/g),而当压力增至100 MPa时,DTPI-100的游离巯基和二硫键含量分别提升至8.98 μmol/g和11.85 μmol/g。整体而言,热处理和动态高压微射流处理对 TPI游离巯基和二硫键含量的改变较为有限,说明HTPI和DTPI的聚集体可能以疏水性聚集为主,而非以二硫键交联形成的共价聚集。
2.3 物理处理对黄粉虫蛋白溶解度的影响
商业化的蛋白产品生产过程中通常包括热处理等一系列的物理处理,可能导致蛋白的变性、聚集,影响蛋白的溶解度。图4为天然和物理处理TPI的溶解度-pH曲线,显示了天然和物理处理TPI溶解度具有pH依赖性,呈现U型趋势,且所有蛋白的NPI的pI均约为4.5,说明本文中的物理处理不会造成TPI的pI偏移。NTPI的溶解度较低,在pH值2.0~9.0间的溶解度均低于30%。热处理后的HTPI的溶解度略低于NTPI,且HTPI-90和HTPI-120无明显差异。蛋白经过高于变性温度的热处理后易发生高度去折叠,疏水基团暴露,带电氨基酸在蛋白分子表面重排,蛋白分子表面电荷变化,同时蛋白与水分子间的相互作用减弱,导致蛋白溶解度的下降。Lee等[6]对黄粉虫蛋白进行55~95 ℃热处理后,蛋白溶解度在pH值2.0~11.0也出现一定程度的下降。
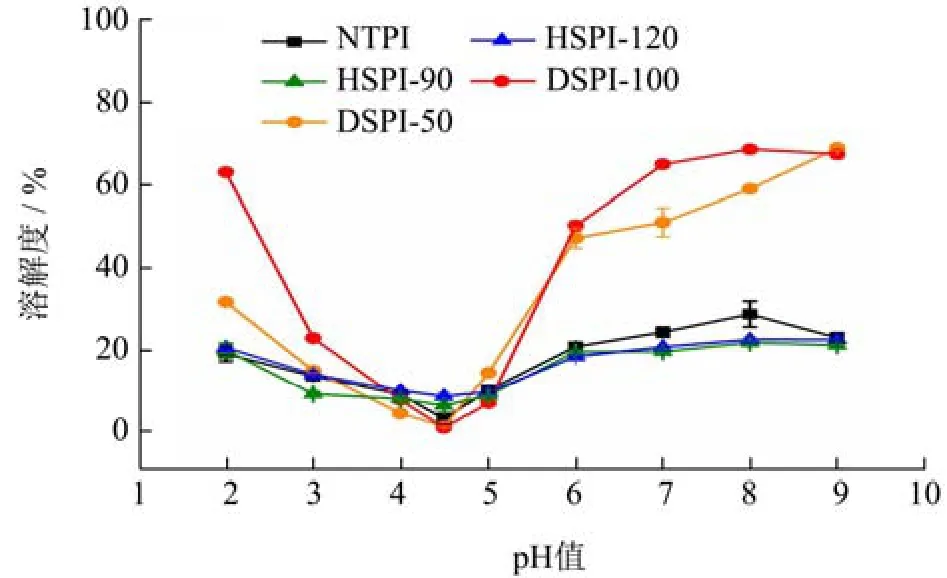
图4 物理处理对TPI溶解度的影响Fig.4 Effects of physical treatments on TPI solubility
动态高压微射流可显著提高TPI的溶解性,且溶解性随着处理压力的提高而增强。pH值7.0时,DTPI-50和DTPI-100的溶解度由24.05%分别提升至50.80%和64.81%,这可能是由于动态高压微射流的高剪切力、冲击力和空穴效应所致。微射流处理过程中产生强大的机械力和冲击力可破坏大分子的有序结构,同时空穴效应可破坏蛋白的氢键和疏水相互作用,加强蛋白与水分子间的相互作用,提升难溶蛋白的可溶性[9]。Fan[11]等对燕窝水提后的沉淀物进行高压微射流处理,发现蛋白的水溶性显著增强,且当处理压力在40~120 MPa之间时,溶解度与处理压力也呈正相关趋势。Shen[10]等也发现对大豆蛋白进行热处理会导致其在中性条件下溶解度的下降,但120 MPa的微射流处理可提高热处理大豆蛋白的溶解性至与天然大豆蛋白同一水平。
2.4 物理处理对黄粉虫蛋白乳化性和起泡性的影响
乳化活性(EAI)和乳化稳定性(ESI)是评价其蛋白乳化性能的重要指标,结果见表1。热处理可大幅提升 TPI的乳化活性,其中 HTPI-90的 EAI值由14.82 m2/g提升至22.12 m2/g,HTPI-120的EAI值较HTPI-90略有提升但无显著性差异。热处理虽然降低了TPI的溶解性(见图4),但显著改善了其乳化活性,说明HTPI的疏水性聚集可能在其乳化活性的提升中发挥主要作用。高温条件下TPI的结构充分展开,暴露出更多疏水基团,具有更良好的两亲性,故更易吸附于油滴表面。Shen等[10]也认为热处理对球蛋白乳化活性的改善与其表面疏水性的提高有直接相关性。但Lee等[6]发现热处理降低了TPI的乳化活性,这可能是由于其热处理强度不同,所得HTPI的结构与本文有显著差异所致。普遍认为,适宜的热变性和去折叠才能提升球蛋白的乳化活性,过度加热可能破坏蛋白的乳化性。HTPI的乳化稳定性显著低于NTPI,可能是由于热处理提升了TPI的表面疏水性,促进了相邻液滴表面蛋白分子间的相互作用,导致液滴的絮凝和聚结。高压微射流处理虽然大幅提升了TPI的溶解性(见图4),但对TPI的乳化活性与乳化稳定性无显著影响,这可能是由于DTPI的构象变化对其乳化性产生了负面影响。蛋白经微射流处理后,可能具有更稳定的三级结构[10],而乳化活性与三级结构的柔性程度呈正相关[23],故DTPI的乳化性并无显著改善。
表1也反映了物理处理对TPI起泡能力(FC)及起泡稳定性(FS)的影响。通常而言,蛋白的FC受蛋白内在性质如尺寸、表面电荷、表面疏水性和结构柔性等因素的影响。HTPI的FC值从74.18%提升至80.77%,且不同温度处理后的样品无差异,这可能是因为热处理后蛋白的构象变化导致其内部疏水基团暴露,分子柔性增强,提升了HTPI在界面的吸附效率[5]。DTPI在中性条件下溶解度提升(见图4),说明蛋白分子间的静电斥力增强,且DTPI对蛋白具有一定的解聚作用[11],可生成较小尺寸的蛋白结构,但 DTPI与NTPI的FC并无显著差异,这可能是由于微射流处理后的蛋白呈现较稳定的三级结构[10],不利于其 FC的改善。不同物理处理均提升了TPI的FS,且高压动
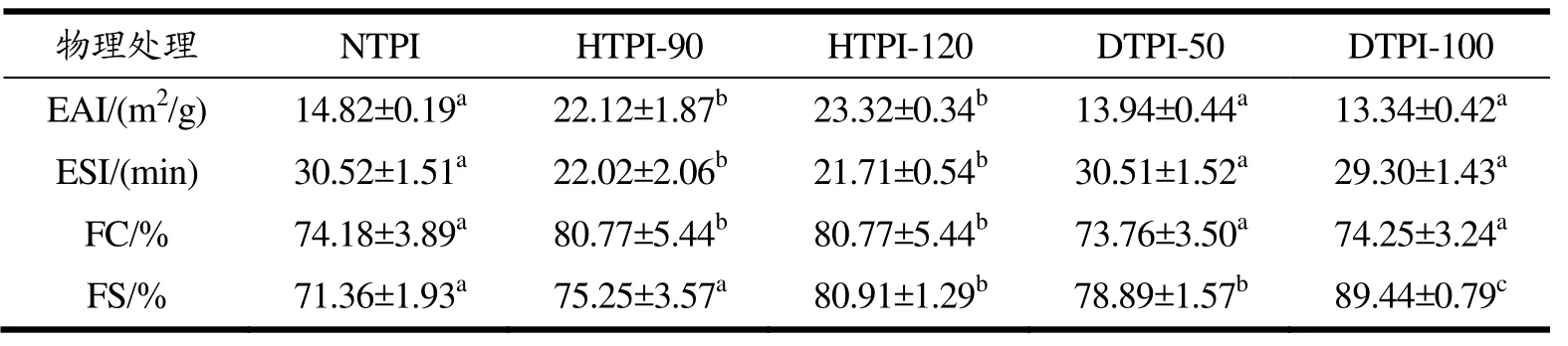
表1 物理处理对TPI乳化性和起泡性的影响Table 1 Effects of physical treatments on the emulsification and foamabilityof TPI
态微射流处理对 FS的改善效果优于热处理。泡沫的不稳定性主要由排液、聚结和奥氏熟化等原因所致[24],提升蛋白分子间的静电斥力和界面膜的粘弹性有利于提升泡沫的稳定性。HTPI-120的FC值高于HTPI-90(80.91% vs 75.25%),这可能是由于120 ℃处理后蛋白的聚集状态更有利于其凝胶状界面膜的形成,提供了有效的空间位阻,避免了泡沫因聚结导致的破灭。课题组前期研究也发现对大豆分离蛋白进行亚临界水处理(121 ℃)后的 FS和 FC均优于传统热处理(90 ℃)[5]。DTPI-120的FS值最高(89.44%),可能归因于微射流处理提升了蛋白分子间的静电斥力,削弱了泡沫表面蛋白分子间的相互作用,降低了泡沫的聚结,从源头上避免了泡沫破灭。
2.5 物理处理对黄粉虫蛋白体外消化性影响
对天然和物理处理TPI进行模拟体外消化并监测消化过程中亚基的降解情况,见图5。所有样品在模拟胃消化10 min时的电泳条带均无明显变化,说明胃蛋白酶酶解10 min时对TPI亚基并无水解作用。NTPI在胃消化30 min时所有的亚基条带基本消失,在11 ku处出现深色条带,可能是亚基水解后的小分子产物。随着模拟肠消化的进行,NTPI的亚基进一步水解,且11 ku处条带的颜色逐渐变浅,说明胰蛋白酶对该水解产物具有一定的酶解作用。HTPI-90与 NTPI的SDS-PAGE图谱变化基本一致。HTPI-120在模拟胃消化30 min时,点样孔处的深色条带明显变浅,说明胃蛋白酶可水解该聚集体。体外消化结束后,除 11 ku处条带外,HTPI-120在浓缩胶顶端仍可见条带,说明120 ℃热处理形成的TPI聚集体具有较差的体外消化性。本课题组前期研究也报道了喷射蒸煮处理中的高温(120 ℃)导致天然米糠蛋白的热变性与聚集,引起其消化性的降低[25]。动态高压微射流处理后的 TPI表现出明显的抗消化性。模拟胃消化60 min时,DTPI在45 ku和17 ku附近仍有条带。模拟肠消化开始后,45 ku处条带消失,但体外消化结束后,17 ku处条带仍存在,这可能是由于微射流处理引起了TPI的疏水性聚集,增强了TPI表皮蛋白中部分亚基的抗消化性。
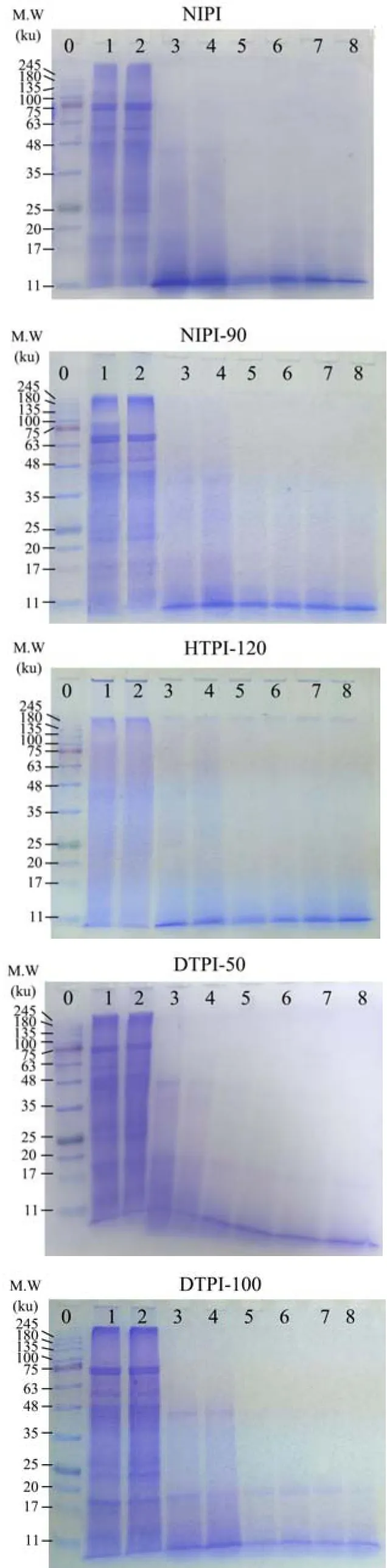
图5 物理处理后TPI体外消化过程的SDS-PAGE图谱Fig.5 SDS-PAGE profiles of TPI after physical treatments during in vitro digestion
蛋白体外消化过程中的可溶性氮释放的变化情况可补充SDS-PAGE结果反映蛋白的消化情况,见图6。在模拟胃消化10 min时,所有样品无明显氮释放,说明此时蛋白的水解度较低,这与SDS-PAGE图谱一致。当胃消化进行至30 min时,所有样品的氮释放量均急剧增加,随着消化的进行,氮释放量的增速逐渐减缓,直至体外消化结束时达到最大值。整体而言,NTPI的氮释放量最高,在3 h为39.53%。热处理降低了TPI的体外消化性,与 SDS-PAGE图谱一致(图5),但HTPI-90和HTPI-120的差异并不显著,二者在3 h时氮释放量分别为35.58%和33.90%。DTPI-50在整个消化过程中都表现出极强的抗消化性,其在3 h时氮释放量仅为29.02%。当处理压力增至100 MPa时,DTPI-100在体外消化60~180 min内的氮释放量均基本与NTPI持平,说明高压力下的微射流处理虽然降低了TPI中部分亚基的消化性(图5),但可能提升了TPI水解物的消化性,水解物被继续裂解成分子量更小的肽,难以染色或跑出分离胶,故其SDS-PAGE图谱中(图5)虽可见未降解亚基的条带,但其氮释放量仍可维持较高水平。
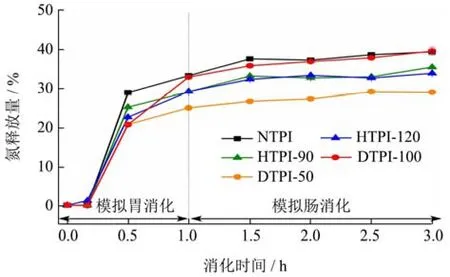
图6 TPI体外消化过程中氮释放量的变化Fig.6 Changes of nitrogen release during TPI in vitrodigestion
3 结论
碱溶酸沉法提取的TPI主要由疏水相互作用、氢键及二硫键稳定其高级结构,且疏水相互作用占主导地位。热处理和动态高压微射流处理均能诱导TPI大分子聚集体的形成,且以疏水性聚集为主,而非以二硫键交联形成的共价聚集。热处理TPI的溶解度整体略低于天然 TPI,而动态高压微射流可显著提升 TPI的溶解度。pH值7.0时,DTPI-50和DPI-100的溶解度由24.05%分别提升至50.80%和64.81%。热处理提升TPI的乳化活性、起泡能力和泡沫稳定性,但降低了其乳化稳定性;微射流处理对TPI的乳化性能和起泡能力均无显著影响,但可大幅提升其泡沫稳定性。热处理和动态高压微射流处理均导致了TPI的消化性能的降低。在体外模拟消化结束时,HTPI-90和HTPI-120的氮释放量由 NTPI的 39.53%分别降至35.58%和33.90%,DTPI-50和DPI-100的SDS-PAGE图谱中均可见未被完全降解的亚基条带。热处理和微射流处理对TPI进行改性后,黄粉虫蛋白作为食品配料可能可以用于蛋白饮料、乳液和泡沫食品中。
猜你喜欢
食品科学技术学报(2022年5期)2022-10-11
广州化学(2022年4期)2022-09-01
环境卫生工程(2021年2期)2021-06-09
生物技术通报(2021年12期)2021-02-10
工业水处理(2020年10期)2020-10-22
绿色科技(2019年10期)2019-06-17
分析化学(2019年4期)2019-05-13
广西林业科学(2019年1期)2019-04-26
中国林副特产(2017年1期)2017-02-21
中国饲料(2016年17期)2016-12-01
