
植物体细胞胚胎发生的分子机制
2023-02-10 01:26:34孙思敏孙伟男杨细燕张献龙
作物学报 2023年2期
韩 贝 孙思敏 孙伟男 杨细燕 张献龙
綜述
植物体细胞胚胎发生的分子机制
韩 贝 孙思敏 孙伟男 杨细燕*张献龙
华中农业大学作物遗传改良全国重点实验室, 湖北武汉 430070
植物细胞的全能性是指每个细胞均具有该植物的全部遗传信息, 其离体组织或细胞在适当培养条件下具有发育成完整植株的潜能。植物体细胞胚胎发生是最能体现植物细胞全能性的一种方式, 其在人工种子、单倍体育种、无性繁殖和种质保存等领域具有广阔的应用前景, 其发生的机制也是基础研究领域的热点。近年来, 随着技术的进步及研究的深入, 植物体细胞胚胎发生的分子调控机制取得了重要进展。植物体细胞胚胎发生是一系列基因在时空顺序上表达调控的结果。本文系统综述了体细胞胚胎发生过程中激素及逆境胁迫信号转导、胚胎发育相关转录因子、胞外蛋白和表观遗传调控的作用, 并对本领域未来的研究重点及方向进行了展望。
体细胞胚胎发生; 激素及逆境胁迫信号转导; 转录因子; 表观遗传修饰
植物体细胞胚胎发生是指由体细胞经过离体培养产生胚状体的过程, 该过程可直接从外植体表皮、亚表皮、悬浮细胞、原生质体发生, 也可以从脱分化的外植体形成愈伤组织的外部或内部产生。从体细胞向胚性细胞转变是体细胞胚胎发生的前提, 该过程中离体的植物细胞经历脱分化形成愈伤组织, 然后脱分化状态的愈伤组织和细胞经历再分化过程, 再度分化成不同类型的细胞、组织和器官, 甚至最终再生成完整的植株。这一过程涉及细胞的重编程、激活细胞周期、细胞分化及器官发育过程, 受到众多转录因子、激素信号传导途径及表观遗传修饰等构成的复杂网络调控。本研究就体细胞胚胎发生过程中关键基因的功能及调控机制、表观遗传机制及关键基因的应用等进行综述, 为深入解析体细胞胚胎发生的分子机制及细胞全能性的研究提供理论依据。
1 激素信号转导与植物体细胞胚胎发生
在体细胞胚胎发生过程中, 通过不同激素的配合使用可以有效调控体细胞胚胎发生的各个发育阶段。比如, 通过调节生长素(auxin)与细胞分裂素(cytokinin, CTK)比率促进脱分化和愈伤增殖, 通过添加低浓度的乙烯(ethylene)促进体细胞胚胎发生, 通过赤霉素(gibberellins, GAs)调控胚性培养物到球形胚的转化, 通过添加脱落酸(abscisic acid, ABA)来提高体细胞胚的质量等。外源激素对体细胞胚胎的作用主要是通过胞外或胞内的激素受体将外界刺激信号转到核内, 从而调控基因的表达, 启动发育程序。
1.1 生长素信号传导及体细胞胚胎发生
生长素是诱导植物体细胞胚胎发生过程的重要植物生长调节剂。其中2,4-D在植物体细胞胚胎发生中的应用较为广泛。一定浓度的生长素能够促进愈伤组织的诱导和增殖。在棉花体细胞到胚性细胞的转变中, 伴随着内源生长素水平的升高, 表现为生长素信号途径的激活[1]。与生长素合成、极性运输和响应相关的基因被认为是植物体细胞胚胎发生的关键基因。目前已从不同植物中已解析出多个受生长素诱导表达的基因, 这些基因包括(Gretchen Hagen 3)、(Pin-formed)、生长素/吲哚乙酸蛋白基因(auxin/indole-3-acetic acid,)、生长素响应因子基因(auxin response factors,)和(small auxin-up RNAs)。生长素信号通路依赖生长素响应因子, 尤其是(jumonji C domain-containing protein 30)与和相互作用并直接与(lateral organ boundaries domain)家族基因(,等)的瞬时元件结合激活它们的表达,家族基因诱导的表达, 促进愈伤组织的增殖(图1)[2-4]。另外, 在体细胞胚胎发生过程中, 外源生长素能激活胚胎发育相关转录因子调控网络, 转录因子BABY BOOM (BBM)和LEC1- ABI3-FUS3-LEC2 (LAFL)复合物是体细胞胚胎发生的主要调节因子。BBM编码AIL (aintegumenta-like)并直接调节所有LAFL基因[5]。同时LAFL基因也可以激活生长素合成和运输进而调控体细胞胚胎发生。在体细胞胚胎发生过程中, 在SAM区域的较高浓度的生长素会诱导(WUSCHEL)基因的表达, 进而激活PIN1蛋白的极性定位, 促进体细胞胚胎的生长和发育[6]。
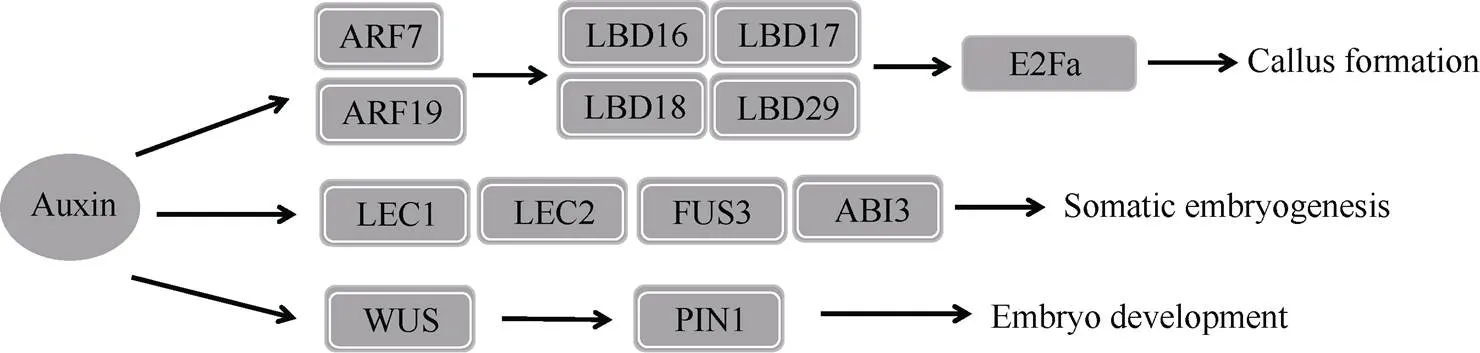
图1 生长素信号传导及体细胞胚胎发生
1.2 细胞分裂素信号与愈伤组织的增殖
在植物体细胞胚胎发生过程中, 大部分情况下生长素不是独立发挥作用, 只有当生长素和细胞分裂素维持在合适的比例时, 才能促进愈伤组织的产生[7]。细胞分裂素信号转导是一个“磷酸接力传递”的过程。细胞分裂素首先结合受体组氨酸激酶(histidine kinases, HK)并使其磷酸化, 将磷酸基团转移给胞质中的磷酸转运蛋白(histidine-phosphotransfer protein, HP), 磷酸化的HP进入细胞核并将磷酸基团转移到A型和B型反应调节因子(Arabidopsis response regulator, ARR)上, 进而进行信号的传递和激活。拟南芥中超表达促使细胞周期相关基因的表达, 从而促进愈伤组织的分裂与增殖[8]。细胞分裂素信号的抑制基因和的超表达会抑制拟南芥根系顶端分生组织的起始和拟南芥的体细胞胚胎发生, 在拟南芥细胞分裂素受体基因和双突变体中, 再生体细胞胚的根尖和茎尖发育畸形[9]。
1.3 乙烯信号与体细胞胚胎发生
植株的再生能力与乙烯的敏感性有一定的关联, 乙烯含量的升高及其信号途径的激活有利于体细胞胚胎发生[10]。拟南芥中的研究表明, 用乙烯合成抑制剂处理或超表达乙烯合成抑制基因(ethylene- overproducer 1)会降低外植体的再生能力, 乙烯不敏感突变体(、等)的再生能力下降, 而乙烯响应负相关因子突变体(、)的再生能力增强[11]。在大豆体细胞胚胎发生过程中, 添加一定浓度的乙烯合成前体1-氨基环丙烷-1-羧酸(1-aminocylopropane-1-carboxylic acid, ACC)显著促进大豆的体细胞胚胎发生效率, 而利用乙烯抑制剂氨基乙氧基乙烯甘氨酸(aminoethoxyvinylglycine, AVG)等处理则显著抑制大豆的体细胞胚胎发生效率[12]; 在棉花中, 乙烯及其信号对愈伤组织的增殖起着正向调控作用。超表达(squamosa- promoter binding protein-like 10)促使乙烯合成酶相关基因(1-aminocyclopropane carboxylic acid oxidases)的表达, 从而提高愈伤组织的增殖率, 乙烯合成抑制剂AVG抑制处理降低乙烯的含量并显著抑制愈伤组织的增殖; 而乙烯合成前体ACC处理则显著促进外植体愈伤组织的增殖[13]。
2 逆境胁迫信号与愈伤组织再生
理论上所有离体植物细胞在合适的外界环境下均可表现全能性。外界环境因素(特别是逆境因素)是体细胞胚胎发生的重要影响因素。逆境因子(包括机械损伤)在体细胞胚胎发生的几个主要阶段都起着重要的作用, 很多研究者都将逆境因子的调控作为优化体细胞胚胎发生体系的重要手段。在现有的诱导愈伤组织体系中, 培养基多采用MS培养基, 相对于维持植株正常生长或萌发胚生根成苗的含有逆境因子的低糖、低盐、低渗的SH培养基及1/2 MS培养基来说, MS培养基无机盐含量较高, 微量元素种类较全, 浓度也较高。另外, 很多物种, 包括胡萝卜、苜蓿、烟草等, 均有通过利用逆境处理来促进体细胞胚胎形成及发育的研究, 涉及到的逆境因素也多种多样, 主要有ABA处理、饥饿处理、渗透胁迫、高温处理等[14-15]。
机械损伤是愈伤组织诱导原初诱导触发因子[16]。再生的发生一般都是在创伤部位开始的。创伤可以诱导许多细胞反应, 包括激素的积累(茉莉酸)、Ca2+快速流入、活性氧(reactive oxygen species, ROS)爆发、细胞间远程通讯中断等。拟南芥(WIND1 WOUND INDUCED DEDIFFERENTIATION 1)及其同源物WIND2~WIND4是这一过程的中心调节因子。WIND1~WIND4是AP2/ERF转录因子家族成员, 在愈伤组织诱导过程中会在受伤处诱导迅速表达, 并促进细胞的脱分化及愈伤组织增殖。异位表达可使成熟的体细胞摆脱正常的分化程序而启动诱导在组织和结构上无定形的细胞增殖, 以及保持其脱分化状态[17]。转基因植株()细胞状态与2,4-D所诱导的愈伤组织细胞类似。WIND1蛋白是细胞分裂素应答的正调节因子, 主要通过对B型基因促进细胞分裂素信号传导并直接激活(enhancer of shoot regeneration1)从而促进愈伤组织的形成(图2)[17]。在损伤时也会诱导(sopentenyltransferase 3)、(lonely guy 1)、(lonely guy 4)和(lonely guy 5)并提高细胞分裂素水平, 进而通过(D-type cyclin 3;1)重新激活细胞周期[18]。其他(APETALA2/ethylene response factor)基因, 如和(PLETHORA3)、、也在伤口诱导的愈伤组织形成过程中上调表达, 并且还发现AP2/ERF和CTK介导的途径调控网络广泛串联[19]。
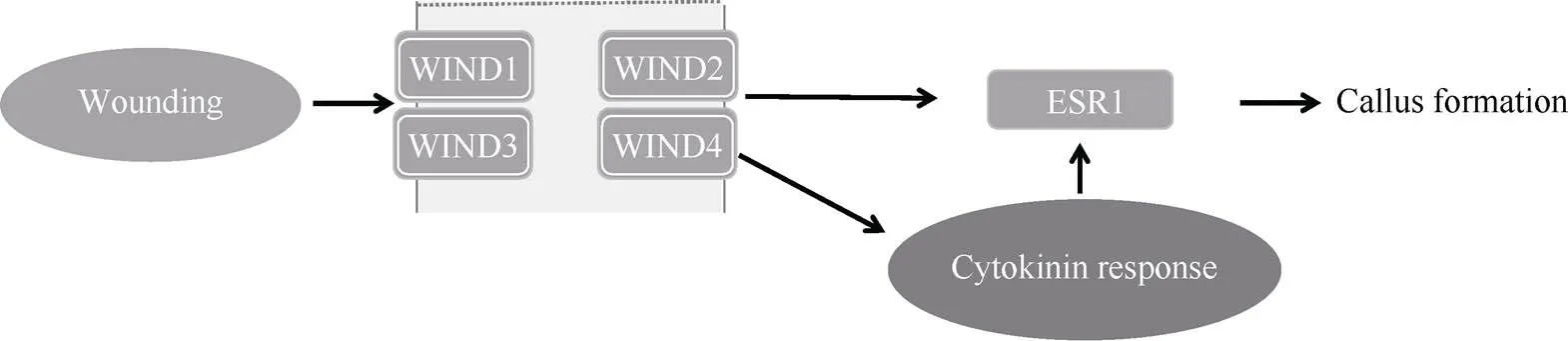
图2 机械损伤通过调节因子WINs调控愈伤组织形成
3 转录因子与体细胞胚胎发生
虽然体细胞胚胎发生过程受诸多外界环境因素的影响, 但归根结底是在各种因素的作用下, 体细胞中某些特异的基因启动表达, 从而使体细胞脱分化并再分化转变为胚性细胞。许多研究人员致力于解析体细胞胚胎发生过程的相关基因, 目前已鉴定和克隆了大量与体细胞胚胎发生相关的转录因子, 其中许多参与调节合子胚发生、分生组织分化和维持的转录因子都在体细胞胚胎发生过程中起着重要用。
3.1 核因子Y (nuclear factor Y, NF-Y)
核因子Y (nuclear factor Y, NF-Y), 又称CCAAT盒结合因子(CCAAT-binding factor, CBF)或亚铁血红素激活蛋白(heme activator protein, HAP), 是一类普遍存在于酵母、动物、植物等真核生物中的转录因子, 通常由3种不同亚基组成, 即NF-YA (CBF-B或HAP2)、NF-YB (CBF-A或HAP3)和NF-YC (CBF-C或HAP5) 3个家族[20]。植物NF-YB对胚胎发生具有重要作用[21]。拟南芥中NF-YB家族共有13个成员, 分为LEAFY COTYLEDON 1 (LEC1)类和非LEC1 2类蛋白, 其中LEC1类包括LEC1和LEC1-LIKE (L1L), 是植物胚胎形成的中枢调控因子[22]。在胚性细胞、体细胞胚和未成熟种子中高表达, 并可赋予体细胞向胚性细胞发育的能力, 在体细胞胚胎发育的早期能维持胚性细胞的命运, 目前在多个物种中将其作为体细胞胚胎发生的标记基因[23-24]。与类似, 在胚性愈伤组织、体细胞胚和未成熟种子中高表达, 而在非胚性愈伤组织、营养组织中表达量较低,的异位表达可以代替发挥作用, 在柑橘营养组织中异位表达会诱导出胚[25]。
3.2 B3结构域转录因子
B3结构域转录因子家族属于植物特异性转录因子家族。其中、(FUSCA3)和是含有B3结构域的与胚胎发育相关的调控因子, 是一类胚胎发育相关的标志基因, 与存在协同调控作用[26-27]。过量表达(或异位表达)基因调控、、、(abscisic acid-insensitive 3)和(wrinkeled 1)等转录因子基因的表达, 并促使体细胞向胚性细胞的转变, 从而使转基因植物具有了胚胎的特性[27]。与促进胚性细胞的形成不同,可直接诱导形成体细胞胚胎, 两者可能激活不同的调控路径[28]。同样,基因在顶端分生组织特异表达, 可以促使转基因植物在顶端分生组织产生体细胞胚[27]。、和的下调则显著抑制直接和间接体细胞胚胎发生[5]。、和同时受生长素的诱导表达, 而且也可以对生长素合成基因(YUCCA4)直接调控, 同时激活生长素响应基因的表达, 进而激活生长素信号传导途径[29]。
3.3 AP2/ERF结构域蛋白
AP2/ERF结构域蛋白是植物特异性转录因子家族, 它们参与许多发育过程的调节。有几个AP2/ ERF家族成员参与调控体细胞胚胎发生, 其中研究得最多的是BBM。最初是从甘蓝型油菜花粉体细胞胚中分离得到的, 并在花粉体细胞胚和合子胚中表达。在拟南芥、辣椒、毛白杨等物种中的研究表明基因是植物细胞全能性的关键调控因子, 在体细胞胚胎发生过程中该基因能促进细胞分裂和形态发生, 异位表达基因或者拟南芥基因能在没有外源植物生长调节剂或胁迫的情况下诱导体细胞胚胎的发生[30]。BBM能结合基因的启动子, 从而调控这些基因的表达, 进而调控(WUSCHEL-RELATED HOMEOBOX 2)和等早期胚胎发生基因; 反过来,的表达也受到LAFL蛋白质的调节, 它们形成交互网络共同促进体细胞胚胎发生[31-32]。另一个成员是拟南芥(EMBRYO MAKER)在胚发育的早期和成熟胚中表达, 异位表达促进拟南芥子叶体细胞胚胎发生的启动[33]。在机械损伤部分提到过的也是AP2/ERF转录因子家族中调控体细胞胚胎发生的成员,可以与途径相互作用,与单独激活或相比, 在外植体中按顺序依次激活和能诱导出更多的胚性愈伤[19]。
3.4 同源异形域转录因子
同源异形域(homeodomain, HD)是指真核生物中一个由约60个氨基酸组成的高度保守的具有转录因子特征的DNA结合域, 其最早是在果蝇体节发育中发现并命名的, 之后在高等植物中发现包含该类结构域的基因有重要作用。其中WOX转录因子家族在胚的发育、顶端分生组织建成和器官形成中发挥重要作用。是WOX家族中最早发现的成员, 在分生组织阶段该基因的表达受生长素和细胞分裂素的诱导, 调控分生组织维持的特性; 反之,基因调控体细胞胚胎发生过程中生长素依赖的营养组织向胚性组织转变[34-35]。超表达会诱导体细胞胚胎发生及茎尖和根尖器官发生, 异位表达基因可使那些难以诱导体细胞胚胎发生的顽拗型材料去分化产生不定芽和体细胞胚[35-36]。基因是细胞分裂素应答的负调节因子, 通过抑制A型基因的表达, 与细胞分裂素信号传导途径协同作用, 进而决定体细胞胚胎过程中某些细胞的命运[37]。另外由于可以穿过顶细胞层并激活其负调控因子(CLAVATA3)的转录, 这种WUS-CLV的反馈回路可以维持干细胞库的稳定和顶端分生组织的发育[34,36]。WOX家族其他成员也被发现具有调控体细胞胚胎发生的功能。在拟南芥中,是胚胎发育中细胞命运决定和顶端分化的调控因子, 包括顶端分生组织干细胞的启动, 发育后期还参与侧生器官的形成和分离[38]。在松树中被作为体细胞胚胎发生的标志基因[39]。(SHOOT MERISTEMLESS)是同源异性域转录因子KNOXI (KNOTTED1-like homeobox I)家族的成员, 与和共同作用保持顶端分生组织的干细胞微环境, 并调节细胞增殖和分化的平衡[40]。甘蓝型油菜中的异位表达可以调节对外源生长素的敏感性并促进拟南芥体细胞胚胎发生[41]。
4 体细胞胚胎发生过程中的信号转导
4.1 类受体激酶与体细胞胚胎发生
SERK (somatic embryogenesis receptor-like kinase)是体细胞胚胎发生过程中促进体细胞向胚性细胞转变的关键激酶。它首先是在研究胡萝卜体细胞向胚性细胞转变的过程中发现的, 其仅在胚性细胞的球形期表达, 而在非胚性细胞或其他时期的体细胞胚胎中均不表达[42]。SERK属于富含亮氨酸重复序列受体类似激酶(leucine-rich repeat receptor-like kinase, LRR-RLK)亚家族的成员, 通过识别分子信号进而介导其蛋白的LRR结构域与胞外蛋白结合, 诱导细胞内的信号级联放大。这些信号可识别不同的靶标, 并利用染色质重塑提高体细胞胚胎发生早期响应基因的表达(如和), 从而诱导组织或体细胞向胚性细胞发生转变[43]。异位表达拟南芥基因可促进其体细胞胚胎发生,该基因可作为体细胞胚胎发生过程中标记基因[44]。植物激素对SERK基因的表达和信号转导起重要作用。的表达受到生长素和细胞分裂素的协同调控,和则参与生长素特异性反应, 并且和与油菜素内酯信号传导有关[45]。也是属于LRR- RLK亚家族的成员, 虽与SERK同属一个家族, 但它们在体细胞胚胎发生中的功能却是相反的, 在芸薹属植物中,抑制体细胞胚胎发生转录因子(如)的表达进而抑制体细胞胚胎发生[38]。
4.2 钙信号与体细胞胚胎发生
Ca2+作为第二信使, 在植物生长发育及环境响应中起着重要的作用。有研究认为液泡Ca2+浓度增加是识别胚胎发生细胞的原初信号, 胚性细胞启动胚胎发育时伴随着较大的Ca2+浓度的波动, Ca2+的梯度调控胚胎发育阶段极性分化及器官建成[46]。较高浓度的Ca2+提高胚性愈伤组织的形成和胚胎发生的频率, 缺乏Ca2+会阻止体细胞胚的形成。
细胞Ca2+信号由Ca2+感受器进行感受并传递。植物中存在3种主要的Ca2+感受器。被熟知的是钙调素(calmodulin, CaM)和类钙调素蛋白(calmodulin- like proteins, CMLs)[47]。CaM受生长素诱导表达, 并在诱导体细胞胚胎发生后期(胚性愈伤到胚发育阶段)特异性表达, 一般定位于发育中胚胎的分生组织区域。CaM具有促进细胞增殖的作用; 采用钙调素抑制剂W7 [-(6-氨基己基)-5-氯-1-萘磺胺盐酸盐]处理则会抑制体细胞胚胎发生。钙依赖蛋白激酶(calcium-dependent protein kinase, CDPK)是第2类Ca2+感受器, 它包含一个C-末端CaM样结构域, 可以直接结合Ca2+并进行下游信号的传递, 阻断CDPK介导的信号通路抑制胚胎发生[48]。第3类Ca2+感受器是钙调磷酸酶B亚基类似蛋白(calcineurin B-like protein, CBL)家族, 与CaM不同, CBLs只在高等植物中存在, 并特异与蛋白质激酶CIPKs (CBL-interacting protein kinases)家族相互作用进行钙信号转导[49]。在棉花体细胞胚胎发生过程中发现和家族基因差异表达, 超表达基因影响棉花体细胞的脱分化过程, 并可能与生长素信号通路存在交叉互作[1]。
5 胞外蛋白与体细胞胚胎发生
为了在植物中诱导体细胞胚胎发生, 已经开发了多种诱导系统。这些系统中的分子有助于激发植物细胞的胚性潜能, 促进体细胞胚胎的发生。在此过程中也筛选到一些胚胎特异性基因作为区分胚性和非胚性细胞培养物的标记, 并发现了许多胞外蛋白也可作为胚胎发生潜力的标记物或调控体细胞胚胎发生的信号分子[47]。
5.1 阿拉伯半乳糖蛋白
阿拉伯半乳糖蛋白(arabinogalactan-proteins, AGPs)是一类富含羟脯氨酸或脯氨酸的糖蛋白, 广泛存在于植物的细胞壁、细胞膜和组织的细胞间隙, 并可以分泌到培养细胞的培养基中[50]。AGPs能促进多个物种的体细胞胚胎发生, 有些可作为细胞能否进行体细胞胚胎发生的早期标志。添加外源AGPs提高胡萝卜、仙客来、云杉、香蕉和小麦非胚性细胞系的胚性能力, 增加体细胞胚胎的数量[51-52]。去除细胞壁上的AGPs会使原生质体形成体细胞胚胎的能力下降, 而加入胞外AGPs则会逆转去除细胞壁的部分效果。添加从胡萝卜种子AGP中分离的可使失去体细胞胚胎发育能力的细胞系重新启动胚形成, 添加与AGP特异结合的抑制剂Yariv (1,3,5-三[4-β- D葡吡喃糖基-氧化苯基-偶氮基]-2,4,6-三羟基苯)导致细胞增殖受阻, 抑制体细胞胚胎发生[53-54]。AGPs调节体细胞胚胎发生的作用机制可能体现在两方面: (1) AGPs一般存在于胞外或膜的界面, 与细胞壁非共价结合, 能快速移动、降解, 可作为信号分子影响细胞识别及信号传递, 进而影响细胞增殖、分化, 导致发育轨迹的改变; (2) AGP可作为多种酶的底物, 裂解后产生不同种类的寡糖分子, 寡糖分子作为信号分子参与发育调控。有研究表明同时添加几丁质酶和AGP能进一步加快体细胞胚胎发生的频率, 几丁质酶能裂解AGP的寡糖基链, 产生信号分子, 促进体细胞胚胎的发生[55]。
5.2 脂质转运蛋白
脂质转运蛋白(lipid transfer protein, LTP)是一类分子量小于10 kD的小分子可溶性碱性蛋白[56]。在动物中, 有些LTP蛋白参与癌细胞增殖的调控。植物中LTP含量高, 占可溶性蛋白的4%左右。有些非特异性基因在幼嫩组织、营养生长时期的分生组织、幼胚中优势表达, 并在体细胞胚胎发生过程进行富集, 被认为是体细胞胚胎发生诱导的早期标记[58]。(extracellular protein 2)是胡萝卜胚性培养中第一个被分离和鉴定的编码LTP的基因, 其分泌到胞外,在原胚性细胞中高表达, 而在非胚性细胞中不表达[57-58]。棉花基因在下胚轴、非胚性细胞组织和植株中不表达, 但在胚性细胞、球形胚高量表达, 而在球后期体细胞胚胎中表达量又下降[59]。是葡萄体细胞胚胎发育过程中表皮原形成的标志, 异位表达导致胚胎严重畸形, 导致表皮层出现异常[60]。
6 体细胞胚胎发生的表观遗传调控
一般认为, 植物体细胞比动物体细胞具有更强的可塑性, 已分化的离体植物细胞在一定培养条件下可脱分化并获得全能性或亚全能性, 在这个过程中, 植物细胞必须获得改变细胞发育命运的潜能, 这一过程也伴随着染色质水平和基因表达水平的重编程, DNA甲基化、组蛋白去乙酰化/甲基化、miRNA等重要的表观调控因子也是影响体细胞胚胎发生的重要因素[61]。
6.1 DNA甲基化
植物体细胞胚胎发生过程受DNA甲基化的调控。一般来说, 非胚性愈伤组织基因组甲基化水平高, 而胚性愈伤组织的基因组甲基化水平较低, 这一现象在刺五加、甜菜、棉花、黑松等中都能观察到[62]。体细胞胚的发育过程也伴随着DNA甲基化水平的变化, 体细胞胚胎发育早期阶段较低, 随着体细胞胚胎的发育逐渐增加, 到子叶胚阶段达到最高值[63]。在拟南芥等中发现体细胞胚胎发生过程的调节需要一定水平的DNA甲基化维持, 甲基化抑制剂5-氮杂胞苷(5-azacitidine, 5-AzaC)处理诱导DNA胞嘧啶甲基化水平降低, 抑制非胚性愈伤组织的增殖或胚性愈伤组织的体细胞胚胎发生能力[64]。DNA甲基化可通过引起胚胎发生特定基因的启动进而影响体细胞胚胎发生。基因启动子在体细胞中被甲基化, 随着外源培养条件的施加, 其甲基化状态被消除, 促进体细胞向胚性细胞的转变及体细胞胚的产生, 随后在胚胎发育成熟后再次被甲基化[65]; 同样, 在拟南芥中, 5-AzaC处理显著抑制外植体的体细胞胚胎发生能力, 处理后体细胞胚胎发生调节基因、和等的表达也均显著降低, 但在DNA甲基化酶基因(和)突变体中这些基因明显上调表达并提高体细胞胚胎发生频率[64]。DNA甲基化受2,4-D的调控, 在诱导阶段, 高浓度的2,4-D通过DNA甲基化关闭原有分化细胞内基因的表达; 去除2,4-D后使DNA去甲基化。
6.2 组蛋白甲基化
在体细胞胚胎发生过程中, 发现DNA甲基化作用的同时也发现组蛋白甲基化对体细胞胚胎形成的影响。两个研究得较多的组蛋白甲基化修饰是关于组蛋白H3的第27位赖氨酸的三甲基化(H3K27me3)和第4位赖氨酸的三甲基化(H3K4me3), H3K27me3是动、植物发育重要基因的沉默基因转录标记物, 而H3K4me3是转录激活标记物。
组蛋白甲基化由组蛋白甲基转移酶完成。拟南芥中, 组蛋白甲基转移酶PRC2通过H3K27me3来抑制相关基因的表达, 从而促进细胞分化, 反之则引起细胞脱分化, 诱导体细胞胚胎发生[66]。在拟南芥中(Polycomb repressive complex 2)基因(CURLY LEAF,和SWINGER,)或(VERNALIZATION 2,和EMBRYONIC FLOWER 2,)双突变体在茎尖脱分化形成愈伤组织, 间接导致体细胞胚胎发生并形成异位根[67],与野生型相比,的突变体在营养组织中显示出更高的体细胞胚胎诱导能力[68]。大部分胚性相关基因、、和以及分生组织调节因子、和等基因的染色质区域都含有H3K27me3等甲基化位点[69]。而和与胚胎发生转录抑制因子和等互作, 并通过表观修饰抑制胚胎发生相关靶标基因的表达从而抑制愈伤组织的形成和体细胞胚胎发生。除了H3K27me3在体细胞胚胎发生发挥功能外, 在拟南芥还发现赖氨酸特异性去甲基酶LDL3可以在愈伤组织形成过程中特异性消除H3K4me2, 进一步使愈伤组织具有芽分化的能力[70]。
6.3 组蛋白去乙酰化
组蛋白去乙酰化也是一种与植物体细胞胚胎发生密切关联的组蛋白修饰。在激素诱导下组蛋白乙酰化水平和组蛋白去乙酰化酶(histone deacetylase, HDAC)的活性在体细胞胚胎发生过程中会发生明显变化。而且组蛋白H3和H4的乙酰化会促进体细胞胚胎发生。组蛋白乙酰化的水平和位置受到组蛋白乙酰转移酶(histone acetyltransferase, HAT)和组蛋白去乙酰化酶的严格控制, 并影响植物的许多发育过程[71]。HDAC抑制剂曲古抑菌素A (trichostatin A, TSA)处理小麦培养物可增加胚性愈伤诱导率和芽分化率[72]。HDAC双突变体或TSA处理诱导胚性标记基因、和的上调表达, 促进体细胞胚胎发生[73]。在拟南芥利用成熟的合子胚进行体外培养时, 经过TSA处理无需额外施加生长素便能够诱导体细胞胚形成, 这可能是由于TSA处理后生长素合成通路基因(、)、胚胎发育相关转录因子(、、)及胁迫响应因子/等显著上调表达引起的[74-75]。HDA6和HDA19分别特异性结合VAL1 (VP1/abscisic acid insensitive 3-like1)和VAL2 (VP1/abscisic acid insensitive 3-like2), 并通过表观修饰抑制胚胎发生相关靶标基因的表达从而抑制愈伤组织形成和体细胞胚胎发生[76]。(PICKLE)是拟南芥中主要的染色质重塑因子, 在抑制细胞过度增殖中起核心作用。其突变体在种子萌发后形成愈伤组织, 在突变体中和的H3K27me3水平下降导致甲基化抑制解除; 其另一突变体易诱导形成愈伤组织, 而该表型可被外施TSA模拟[77]。
6.4 miRNA的调控作用
植物的miRNA通过转录和转录后水平的基因沉默对植物细胞发育命运进行调控[78]。水稻中发现在分化的胚性愈伤比未分化的胚性愈伤中有更高的表达量; 而表达则相反[79]。柑橘和棉花等物种中也发现类似结果, 同时发现、、、、等也在体细胞胚胎发生过程中差异表达[78]。通过调控其靶基因的表达参与调控体细胞胚胎发生。在柑橘中超表达或敲除其2个靶基因和株系的体细胞胚的数量显著高于对照[80], 而且在体细胞胚胎发生的标志基因的超表达材料中可以发现miR156a在整个体细胞胚胎过程均显著上调[81]。是的另一个成员。棉花中对及其靶标研究发现其通过调控乙烯信号和类黄酮生物合成调控逆境下棉花体细胞胚胎发育[13]。miRNA的调控体细胞胚胎发生可能通过2个方面: (1) miRNA参与植物激素信号的转导。植物体细胞胚胎发生过程中多个激素信号途径相关基因被发现受miRNA调控; (2) 体细胞胚胎发生过程中一些重要转录因子可作为miRNA的直接靶标。miRNA可通过降解互补的mRNA或影响mRNA翻译降低蛋白水平。
7 关键基因在体细胞胚胎发生和遗传转化中的应用
随着研究的不断深入和新技术的涌现, 体细胞胚胎发生所涉及的激素信号转导, 基因转录调控、表观遗传修饰与代谢物动态变化等复杂分子生物学过程都在逐步深入地阐释。然而, 目前虽然通过对组织培养、农杆菌侵染等方法的优化, 拟南芥、杨树、水稻、玉米、棉花等植物的遗传转化体系得以成功建立, 但植物的高效遗传转化仍存在许多问题, 如基因型依赖性严重、转化效率不高。随着对体细胞胚胎发生机制深入研究, 一些调控体细胞胚胎发生的关键基因被发现并逐渐用于改进植物遗传转化体系和再生植株的效率, 从而实现植物的高效遗传转化[82]。
和是2个提高植物遗传转化效率的关键基因。在玉米中用和两个基因共转化未成熟胚时, 愈伤组织的比例显著提升, 且在多个不易转化的玉米自交系中取得较高的转化效率[83]。此外, 在水稻、高粱中利用和的共转化也实现转化效率提高[83]。在双子叶植物甜椒中, Heidmann等[84]发现瞬时表达基因可高效诱导体细胞再生, 并分化出大量可发育成完整植株的体细胞胚胎; 将和或和共转化烟草、番茄、葡萄等植物中可实现了芽的原位诱导[85]。因此解析体细胞胚胎发生过程中关键基因的功能及调控机制, 开发更高效的遗传转化方法, 有望实现更多植物的遗传高效再生体系建立。
8 总结与展望
体细胞胚胎发生是体细胞在没有受精的情况下再生为胚状体的过程, 是展现植物细胞具有全能性的一个重要途径。自20世纪50年代以来, 体细胞胚胎发生的研究经历了从经验方法到系统性理论研究的发展, 并在多个物种成功建立起再生体系[86]。对于体细胞胚胎发生的研究, 从最开始通过对外植体的选择、培养基类型、激素含量的配比及组合等进行优化开展研究; 发展到对植物体细胞胚胎发生调控基因的研究, 并利用调控体细胞胚胎发生的关键基因去提高植物的转化及再生效率; 再到对植物体细胞胚胎发生过程基因的调控网络、代谢物质的变化、表观遗传调控机制的研究, 说明对体细胞胚胎发生的分子机制不断深入及完善。
随着分子生物学的进步和测序技术的发展。多组学技术的应用加速了对植物体细胞胚胎发生分子机制的解析。利用RNA-seq揭示体细胞胚胎发生过程中基因的动态变化, 借助代谢组技术解析体胚发育各个阶段的内源物质变化, 并通过对表观组学的研究揭示体细胞胚胎发生过程中表观遗传机制(DNA甲基化、组蛋白甲基化、组蛋白去乙酰化及miRNA)对基因表达调节。并且随着将来单细胞测序技术、空间转录组等新技术在植物体细胞胚胎发生上的应用, 体细胞胚胎发生起源单细胞还是多细胞的问题有可能得到解决。此外, 随着转基因技术及基因编辑技术利用, 对植物体细胞胚胎过程中基因型依赖严重、转化效率低等问题也在逐步得到改善。
尽管当前体细胞胚胎发生的研究进展迅速, 但仍然存在许多谜团。2005年,创刊125周年之际公布了125个最具挑战性的科学问题, 其中植物细胞全能性被列入最重要的25个科学问题。2021年9月,公布了植物细胞生物学15个最重要的问题, 其中有多个问题都涉及到植物细胞全能性和多能性的表达。理论上, 只要条件合适, 所有的植物细胞都可以表现全能性, 得到再生植株。然而, 植物细胞再生实际上只能在有限的植物中实现; 而且离体细胞胚胎发生和器官再生的细胞学和分子机理的研究都不够完善。体细胞胚胎发生的起源是单细胞还是多细胞?在不同的发生体系中究竟是哪一类或哪几类细胞如何被选中进行重编程, 从而再生?虽然已有研究表明这些再生大都来源于植物的干细胞, 但植物的干细胞究竟如何界定?表观遗传修饰如何参与并协调再生过程?这些问题都是未来研究中的重点。
[1] Yang X Y, Zhang X L, Yuan D J, Jin F Y, Zhang Y C, Xu J. Transcript profiling reveals complex auxin signalling pathway and transcription regulation involved in dedifferentiation and redifferentiation during somatic embryogenesis in cotton., 2012, 12: 110.
[2] Lee H W, Kim N Y, Lee D J, Kim J.regulates lateral root formation in combination withdownstream ofandin., 2009, 151: 1377–1389.
[3] Fan M Z, Xu C Y, Xu K, Hu Y X. LATERAL ORGAN BOUNDARIES DOMAIN transcription factors direct callus formation inregeneration., 2012, 22: 1169–1180.
[4] Lee K, Park O-S, Seo P J. JMJ30-mediated demethylation of H3K9me3 drives tissue identity changes to promote callus formation in., 2018, 95: 961–975.
[5] Horstman A, Li M, Heidmann I, Weemen M, Chen B, Muino J M, Angenent G C, Boutilier K. The BABY BOOM transcription factor activates the LEC1-ABI3-FUS3-LEC2 network to induce somatic embryogenesis., 2017, 175: 848–857.
[6] Mendez-Hernandez H A, Ledezma-Rodriguez M, Avilez- Montalvo R N, Juarez-Gomez Y L, Skeete A, Avilez-Montalvo J, De-la-Pena C, Loyola-Vargas V M. Signaling overview of plant somatic embryogenesis., 2019, 10: 77.
[7] Skoog F, Miller C O. Chemical regulation of growth and organ formation in plant tissues cultured in vitro., 1957, 11: 118–130.
[8] Sakai H, Honma T, Aoyama T, Sato S, Kato T, Tabata S, Oka A., a transcription factor for genes immediately responsive to cytokinins., 2001, 294: 1519–1521.
[9] Su Y H, Liu Y B, Bai B, Zhang X S. Establishment of embryonic shoot-root axis is involved in auxin and cytokinin response duringsomatic embryogenesis.,2015, 5: 792.
[10] Neves M, Correia S, Cavaleiro C, Canhoto J. Modulation of organogenesis and somatic embryogenesis by ethylene: an overview.(Basel), 2021, 10: 1208.
[11] Chatfield S P, Raizada M N. Ethylene and shoot regeneration: hookless1 modulates de novo shoot organogenesis in., 2008, 27: 655–666.
[12] Zheng Q L, Zheng Y M, Perry S E. AGAMOUS-like15 promotes somatic embryogenesis inand soybean in part by the control of ethylene biosynthesis and response., 2013, 161: 2113–2127.
[13] Wang L C, Liu N, Wang T Y, Li J Y, Wen T W, Yang X Y, Lindsey K, Zhang X L. Theregulatory module controls initial cellular dedifferentiation and callus proliferation in cotton by modulating ethylene-mediated flavonoid biosynthesis., 2018, 69: 1081–1093.
[14] Langhansova L, Konradova H, Vanek T. Polyethylene glycol and abscisic acid improve maturation and regeneration ofsomatic embryos., 2004, 22: 725–730.
[15] Stasolla C, Yeung E C. Recent advances in conifer somatic embryogenesis: improving somatic embryo quality., 2003, 74: 15–35.
[16] Marhava P, Hoermayer L, Yoshida S, Marhavy P, Benkova E, Friml J. Re-activation of stem cell pathways for pattern restoration in plant wound healing., 2019, 177: 957–969.
[17] Iwase A, Harashima H, Ikeuchi M, Rymen B, Ohnuma M, Komaki S, Morohashi K, Kurata T, Nakata M, Ohme-Takagi M, Grotewold E, Sugimoto K. WIND1 promotes shoot regeneration through transcriptional activation of enhancer of SHOOT REGENERATION1 in., 2017, 29: 54–69.
[18] Ikeuchi M, Favero D S, Sakamoto Y, Iwase A, Coleman D, Rymen B, Sugimoto K. Molecular mechanisms of plant regeneration., 2019, 70: 377–406.
[19] Ikeuchi M, Iwase A, Rymen B, Lambolez A, Kojima M, Takebayashi Y, Heyman J, Watanabe S, Seo M, de Veylder L, Sakakibara H, Sugimoto K. Wounding triggers callus formation via dynamic hormonal and transcriptional changes., 2017, 175: 1158–1174.
[20] Bucher P, Trifonov E N. CCAAT box revisited: bidirectionality, location and context., 1988, 5: 1231–1236.
[21] Pelletier J M, Kwong R W, Park S, Le B H, Baden R, Cagliari A, Hashimoto M, Munoz M D, Fischer R L, Goldberg R B, Harada J J. LEC1 sequentially regulates the transcription of genes involved in diverse developmental processes during seed development.,2017, 114: E6710–E6719.
[22] Kwong R W, Bui A Q, Lee H, Kwong L W, Fischer R L, Goldberg R B, Harada J J. LEAFY COTYLEDON1-LIKE defines a class of regulators essential for embryo development., 2003, 15: 5–18.
[23] Lotan T, Ohto M, Yee K M, West M A L, Lo R, Kwong R W, Yamagishi K, Fischer R L, Goldberg R B, Harada J J.LEAFY COTYLEDON1 is sufficient to induce embryo development in vegetative cells., 1998, 93: 1195–1205.
[24] Orlowska A, Igielski R, Lagowska K, Kepczynska E. Identification of,andgenes and their expression during the induction phase ofGaertn. somatic embryogenesis., 2017, 129: 119–132.
[25] Zhu S P, Wang J, Ye J L, Zhu A D, Guo W W, Deng X X. Isolation and characterization ofgene related to embryogenic competence in., 2014, 119: 1–13.
[26] Le B H, Cheng C, Bui A Q, Wagmaister J A, Henry K F, Pelletier J, Kwong L, Belmonte M, Kirkbride R, Horvath S, Drews G N, Fischer R L, Okamuro J K, Harada J J, Goldberg R B. Global analysis of gene activity duringseed development and identification of seed-specific transcription factors., 2010, 107: 8063–8070.
[27] Gazzarrini S, Tsuchiya Y, Lumba S, Okamoto M, McCourt P. The transcription factorcontrols developmental timing inthrough the hormones gibberellin and abscisic acid., 2004, 7: 373–385.
[28] Brand A, Quimbaya M, Tohme J, Chavarriaga-Aguirre P.andorthologous genes are key regulators of somatic embryogenesis in cassava., 2019, 10: 673.
[29] Stone S L, Braybrook S A, Paula S L, Kwong L W, Meuser J, Pelletier J, Hsieh T F, Fischer R L, Goldberg R B, Harada J J.LEAFY COTYLEDON2 induces maturation traits and auxin activity: implications for somatic embryogenesis., 2008, 105: 3151–3156.
[30] Zhang Z Y, Zhao H, Li W, Wu J M, Zhou Z H, Zhou F, Chen H, Lin Y J. Genome-wide association study of callus induction variation to explore the callus formation mechanism of rice., 2019, 61: 1134–1150.
[31] Horstman A, Li M, Heidmann I, Weemen M, Chen B, Muino J M, Angenent G C, Boutilier K. The BABY BOOM transcription factor activates the LEC1-ABI3-FUS3-LEC2 network to induce somatic embryogenesis., 2017, 175: 848–857.
[32] Mathew M M, Prasad K. Model systems for regeneration:., 2021, 148: dev195347.
[33] Tsuwamoto R, Yokoi S, Takahata Y.encoding an AP2 domain transcription factor plays a key role in developmental change from vegetative to embryonic phase., 2010, 73: 481–492.
[34] Schoof H, Lenhard M, Haecker A, Mayer K F X, Jurgens G, Laux T. The stem cell population ofshoot meristems is maintained by a regulatory loop between theandgenes., 2000, 100: 635–644.
[35] Zuo J R, Niu Q W, Frugis G, Chua N H. Thegene promotes vegetative-to-embryonic transition in., 2002, 30: 349–359.
[36] Zhang T Q, Lian H, Zhou C M, Xu L, Jiao Y, Wang J W. A two-step model for de novo activation of WUSCHEL during plant shoot regeneration., 2017, 29: 1073–1087.
[37] Gordon S P, Chickarmane V S, Ohno C, Meyerowitz E M. Multiple feedback loops through cytokinin signaling control stem cell number within theshoot meristem., 2009, 106: 16529–16534.
[38] Zhang Z, Tucker E, Hermann M, Laux T. A molecular framework for the embryonic initiation of shoot meristem stem cells., 2017, 40: 264–277.
[39] Hassani S B, Trontin J F, Raschke J, Zoglauer K, Rupps A. Constitutive overexpression of a conifer WOX2 homolog affects somatic embryo development in pinus pinaster and promotes somatic embryogenesis and organogenesis inseedlings., 2022, 13: 838421.
[40] Su Y H, Zhou C, Li Y J, Yu Y, Tang L P, Zhang W J, Yao W J, Huang R, Laux T, Zhang X S. Integration of pluripotency pathways regulates stem cell maintenance in theshoot meristem., 2020, 117: 22561–22571.
[41] Elhiti M, Tahir M, Gulden R H, Khamiss K, Stasolla C. Modulation of embryo-forming capacity in culture through the expression ofgenes involved in the regulation of the shoot apical meristem., 2010, 61: 4069–4085.
[42] Schmidt E D L, Guzzo F, Toonen M A J, de Vries S C. A leucine-rich repeat containing receptor-like kinase marks somatic plant cells competent to form embryos., 1997, 124: 2049–2062.
[43] Li H Q, Cai Z P, Wang X J, Li M Z, Cui Y W, Cui N, Yang F, Zhu M S, Zhao J X, Du W B, He K, Yi J, Tax F E, Hou S W, Li J, Gou X P. SERK receptor-like kinases control division patterns of vascular precursors and ground tissue stem cells during embryo development in., 2019, 12: 984–1002.
[44] Hecht V, Vielle-Calzada J P, Hartog M V, Schmidt E D L, Boutilier K, Grossniklaus U, de Vries S C. Thegene is expressed in developing ovules and embryos and enhances embryogenic competence in culture., 2001, 127: 803–816.
[45] Singh A, Khurana P. Ectopic expression ofgenes () control plant growth and development in., 2017, 7: 12368.
[46] Borisjuk N, Sitailo L, Adler K, Malysheva L, Tewes A, Borisjuk L, Manteuffel R. Calreticulin expression in plant cells: developmental regulation, tissue specificity and intracellular distribution., 1998, 206: 504–514.
[47] Yang X, Zhang X. Regulation of somatic embryogenesis in higher plants., 2010, 29: 36–57.
[48] Anil V S, Rao K S. Calcium-mediated signaling during sandalwood somatic embryogenesis. Role for exogenous calcium as second messenger., 2000, 123: 1301–1311.
[49] Pandey G K, Grant J J, Cheong Y H, Kim B G, Li L G, Luan S. Calcineurin-B-like protein CBL9 interacts with target kinase CIPK3 in the regulation of ABA response in seed germination., 2008, 1: 238–248.
[50] Letarte J, Simion E, Miner M, Kasha K J. Arabinogalactans and arabinogalactan-proteins induce embryogenesis in wheat (L.) microspore culture., 2006, 25: 877–877.
[51] Perez-Perez Y, Carneros E, Berenguer E, Solis M T, Barany I, Pintos B, Gomez-Garay A, Risueno M C, Testillano P S. Pectin de-methylesterification and AGP increase promote cell wall remodeling and are required during somatic embryogenesis of., 2019, 9: 1915.
[52] Kreuger M, Vanholst G J. Arabinogalactan-protein epitopes in somatic embryogenesis ofL., 1995, 197: 135–141.
[53] Makowska K, Kaluzniak M, Oleszczuk S, Zimny J, Czaplicki A, Konieczny R. Arabinogalactan proteins improve plant regeneration in barley (L.) anther culture., 2017, 131: 247–257.
[54] Serpe M D, Nothnagel E A. Effects of yariv phenylglycosides oncell suspensions: evidence for the involvement of arabinogalactan-proteins in cell proliferation., 1994, 193: 542–550.
[55] van Hengel A J, Tadesse Z, Immerzeel P, Schols H, van Kammen A, de Vries S C. N-acetylglucosamine and glucosamine- containing arabinogalactan proteins control somatic embryogenesis., 2001, 125: 1880–1890.
[56] Cheng C S, Chen M N, Lai Y T, Chen T, Lin K F, Liu Y J, Lyu P C. Mutagenesis study of rice nonspecific lipid transfer protein 2 reveals residues that contribute to structure and ligand binding., 2008, 70: 695–706.
[57] Sterk P, Booij H, Schellekens G A, Vankammen A, Devries S C. Cell-specific expression of the carrotlipid transfer protein gene., 1991, 3: 907–921.
[58] Dodeman V L, Ducreux G, Kreis M. Zygotic embryogenesis versus somatic embryogenesis., 1997, 48: 1493–1509.
[59] Zeng F C, Zhang X K, Zhu L F, Tu L L, Guo X P, Nie Y H. Isolation and characterization of genes associated to cotton somatic embryogenesis by suppression subtractive hybridization and macroarray., 2006, 60: 167–183.
[60] Francois J, Lallemand M, Fleurat-Lessard P, Laquitaine L, Delrot S, Coutos-Thevenot P, Gomes E. Overexpression of thegene interferes with somatic embryo development in grapevine., 2008, 35: 394–402.
[61] Wojcikowska B, Wojcik A M, Gaj M D. Epigenetic regulation of auxin-induced somatic embryogenesis in plants., 2020, 21: 7.
[62] Bravo S, Bertin A, Turner A, Sepulveda F, Jopia P, Jose Parra M, Castillo R, Hasbun R. Differences in DNA methylation, DNA structure and embryogenesis-related gene expression between embryogenic and non embryogenic lines ofD. don.,2017, 130: 521–529.
[63] Nic-Can G I, Lopez-Torres A, Barredo-Pool F, Wrobel K, Loyola-Vargas V M, Rojas-Herrera R, De-la-Pena C. New insights into somatic embryogenesis:andare epigenetically regulated in., 2013, 8: e72160.
[64] Grzybkowska D, Moronczyk J, Wojcikowska B, Gaj M D. Azacitidine (5-AzaC)-treatment and mutations in DNA methylase genes affect embryogenic response and expression of the genes that are involved in somatic embryogenesis in., 2018, 85: 243–256.
[65] Shibukawa T, Yazawa K, Kikuchi A, Kamada H. Possible involvement of DNA methylation on expression regulation of carrotgene in its 5'-upstream region.,2009, 437: 22–31.
[66] Nakamura M, Batista RA, Kohler C, Hennig L. Polycomb Repressive complex 2-mediated histone modification H3K27me3 is associated with embryogenic potential in Norway spruce., 2020, 71: 6366–6378.
[67] Chanvivattana Y, Bishopp A, Schubert D, Stock C, Moon Y H, Sung Z R, Goodrich J. Interaction of polycomb-group proteins controlling flowering in., 2004, 131: 5263–5276.
[68] Mozgova I, Munoz-Viana R, Hennig L. PRC2 represses hormone-induced somatic embryogenesis in vegetative tissue of., 2017, 13: e1006562.
[69] Liu J, Deng S, Wang H, Ye J, Wu H W, Sun H X, Chua N H.regulates gene sets coordinating seed size and lipid biosynthesis., 2016, 171: 424–436.
[70] Ishihara H, Sugimoto K, Tarr P T, Temman H, Kadokura S, Inui Y, Sakamoto T, Sasaki T, Aida M, Suzuki T. Primed histone demethylation regulates shoot regenerative competency., 2019, 10: 1786.
[71] Kumar V, Thakur J K, Prasad M. Histone acetylation dynamics regulating plant development and stress responses., 2021, 78: 4467–4486.
[72] Bie X M, Dong L, Li X H, Wang H, Gao X Q, Li X G. Trichostatin a and sodium butyrate promotes plant regeneration in common wheat., 2020, 15: 12.
[73] Tanaka M, Kikuchi A, Kamada H. Thehistone deacetylases HDA6 and HDA19 contribute to the repression of embryonic properties after germination., 2008, 146: 149–161.
[74] Wojcikowska B, Botor M, Moronczyk J, Wojcik A M, Nodzynski T, Karcz J, Gaj M D. Trichostatin a triggers an embryogenic transition inexplants via an auxin-related pathway., 2018, 9: 1353.
[75] Moronczyk J, Braszewska A, Wojcikowska B, Chwialkowska K, Nowak K, Wojcik A M, Kwasniewski M, Gaj M D. Insights into the histone acetylation-mediated regulation of the transcription factor genes that control the embryogenic transition in the somatic cells of., 2022, 11: 863.
[76] Zhou Y, Tan B, Luo M, Li Y, Liu C, Chen C, Yu C W, Yang S G, Dong S, Ruan J X, Yuan L B, Zhang Z, Zhao L M, Li C L, Chen H H, Cui Y H, Wu K Q, Huang S Z.interacts withand participates in the repression of seed maturation genes inseedlings., 2013, 25: 134–148.
[77] Furuta K, Kubo M, Sano K, Demura T, Fukuda H, Liu Y G, Shibata D, Kakimoto T. The CKH2/PKL chromatin remodeling factor negatively regulates cytokinin responses incalli., 2011, 52: 618–628.
[78] Yang X, Wang L, Yuan D, Lindsey K, Zhang X. Small RNA and degradome sequencing reveal complex miRNA regulation during cotton somatic embryogenesis., 2013, 64: 1521–1536.
[79] Luo Y C, Zhou H, Li Y, Chen J Y, Yang J H, Chen Y Q, Qu L H. Rice embryogenic calli express a unique set of microRNAs, suggesting regulatory roles of microRNAs in plant post-embryogenic development., 2006, 580: 5111–5116.
[80] Long J, Liu C, Feng M, Liu Y, Wu X, Guo W. miR156-SPL modules regulate induction of somatic embryogenesis in citrus callus., 2018, 69: 2979–2993.
[81] Liu Z, Ge X, Qiu W, Long J, Jia H, Yang W, Dutt M, Wu X, Guo W. Overexpression of thegene encoding a B3 transcription factor promotes somatic embryogenesis in Citrus., 2018, 277: 121–131.
[82] Gordon-Kamm B, Sardesai N, Arling M, Lowe K, Hoerster G, Betts S, Jones T. Using morphogenic genes to improve recovery and regeneration of transgenic plants.(Basel), 2019, 8: 38.
[83] Lowe K, Wu E, Wang N, Hoerster G, Hastings C, Cho M J, Scelonge C, Lenderts B, Chamberlin M, Cushatt J, Wang L, Ryan L, Khan T, Chow-Yiu J, Hua W, Yu M, Banh J, Bao Z, Brink K, Igo E, Rudrappa B, Shamseer P M, Bruce W, Newman L, Shen B, Zheng P, Bidney D, Falco C, Register J, Zhao Z Y, Xu D, Jones T, Gordon-Kamm W. Morphogenic regulators baby boom and wuschel improve monocot transformation., 2016, 28: 1998–2015.
[84] Heidmann I, de Lange B, Lambalk J, Angenent G C, Boutilier K. Efficient sweet pepper transformation mediated by the BABY BOOM transcription factor., 2011, 30: 1107–1115.
[85] Maher M F, Nasti R A, Vollbrecht M, Starker C G, Clark M D, Voytas D F. Plant gene editing through de novo induction of meristems., 2020, 38: 84–89.
[86] Loyola-Vargas V M. The history of somatic embryogenesis. In: Loyola-Vargas V M, Ochoa-Alejo N, eds. Somatic Embryogenesis: Fundamental Aspects and ApplicationsCham: Springer International Publishing, 2016. pp 11–22.
Molecular mechanisms of somatic embryogenesis in plants
HAN Bei, SUN Si-Min, SUN Wei-Nan, YANG Xi-Yan*, and ZHANG Xian-Long
National Key Laboratory of Crop Genetic Improvement, Huazhong Agricultural University, Wuhan 430070, Hubei, China
Plant cell totipotency refers to that each cell has all the genetic information of the plant, andtissues or cells has the potential to develop into a whole plant under appropriate culture conditions. Somatic embryogenesis is the most efficient way to reflect the totipotency of plant cells. It has broad application prospects in the fields of artificial seeds, haploid breeding, asexual reproduction, and germplasm preservation, and its mechanism is also a hotspot in basic research. In recent years, with the development of technology and in-depth research, the molecular regulation mechanism of plant somatic embryogenesis has made important progress. Plant somatic embryogenesis is the result of the expression and regulation of a series of genes in spatiotemporal order. In this review, we systematically reviewed the roles of hormones and stress signal transduction, embryonic development related transcription factors, extracellular proteins, and epigenetic regulation in somatic embryogenesis, and prospected future research priorities and directions in this field.
somatic embryogenesis; hormone and stress signaling transduction pathways; transcription factor; epigenetic regulation
10.3724/SP.J.1006.2023.24103
本研究由国家重点研发计划项目(2018YFD1000907)资助。
This study was supported by the National Key Research and Development Program of China (2018YFD1000907).
杨细燕, E-mail: yxy@mail.hzau.edu.cn
E-mail: bhan_z@163.com
2022-04-08;
2022-08-01;
2022-08-09.
URL: https://kns.cnki.net/kcms/detail/11.1809.S.20220808.1609.002.html
This is an open access article under the CC BY-NC-ND license (http://creativecommons.org/licenses/by-nc-nd/4.0/).
猜你喜欢
西部林业科学(2021年4期)2021-08-11 03:30:32
生物学通报(2021年4期)2021-03-16 05:41:34
今日农业(2020年24期)2020-12-15 16:16:00
科学(2020年4期)2020-11-26 08:27:10
中南林业科技大学学报(2020年10期)2020-11-03 07:55:20
天津农学院学报(2020年3期)2020-10-14 06:15:28
科学大众(2020年12期)2020-08-13 03:22:38
乡村科技(2019年9期)2019-06-26 03:52:04
数码世界(2018年7期)2018-12-24 15:06:05
中西医结合心脑血管病杂志(2016年20期)2016-03-01 04:20:45
