
光信号和光合产物调控豆科植物根瘤形成发育的研究进展*
2023-02-04 05:26李易玲陈平付智丹罗凯杜青高超任俊波杨雪丽刘姗姗杨立达袁晓婷彭新月雍太文杨文钰
中国生态农业学报(中英文) 2023年1期
李易玲,陈平,付智丹,罗凯,杜青,高超,任俊波,杨雪丽,刘姗姗,杨立达,袁晓婷,彭新月,雍太文,杨文钰
(四川农业大学农学院/农业农村部西南作物生理生态与耕作重点实验室/四川省作物带状复合种植工程技术研究中心 成都 611130)
工业固氮生产的氮肥极大地促进了作物增产,但其昂贵的工业成本和潜在的环境污染代价限制了农业生产的可持续性。因此,重视豆科(Fabaceae)植物的生物固氮,充分发挥其固氮潜力对农业的可持续发展具有重要意义。豆科作物是世界上最重要的油料和饲料来源之一,以大豆(Glycine max)为例,大豆产量约占全球豆类作物的68%左右,每年固定16 Tg 氮,占所有豆科植物固氮总量的77%[1]。豆科植物与根瘤菌共生,在根瘤菌固氮酶的作用下将分子态的N2固定为植物可利用的氨,固氮过程极其耗能,需要植株为根瘤菌持续提供能量。豆科植物结瘤过程起始于根系信号物质(类黄酮),被根瘤菌特异性识别后产生结瘤因子(nod factors,NFs),NFs 被植物感知后产生一系列的形态和生理变化,根瘤菌侵染后形成侵染线进入根系内皮层细胞,形成结瘤原基并发育成根瘤。根瘤细胞内,糖代谢产生的苹果酸等能源物质交换根瘤菌固定的氮,维持共生关系。根瘤固氮所需的能源来自植株地上部分光合产物,光环境调控了光合产物的合成及根瘤的形成发育[2]。在光合午休状态下,根瘤可与其他库器官竞争光合产物,诱导地上部分碳水化合物向根瘤分配[3]。因此,豆科植株为满足养分需求和协调内部能量供应,通过相应的控制网络调节根瘤形成发育。本文综述了光通过植株的光形态建成和光合作用控制根瘤形成发育的研究进展,对提高豆科植物结瘤固氮能力进行了展望,以期为豆科植物间套作种植等相关研究提供参考。
1 根瘤形成、发育过程及其影响因素
1.1 根瘤的形成发育过程
根瘤菌和宿主植物需要先交换一系列复杂的信号,从而决定是否建立共生关系[4]。当土壤缺乏氮素时,豆科植物根系会分泌类黄酮与根瘤菌中LysR 家族转录因子NodD 结合促进Nod基因转录形成结瘤因子(NFs),即脂质几丁寡糖(LCO)。植物的赖氨酸基序异源双链(LysM)受体(NFR)检测特异性的LCOs,来放大表皮细胞的NFs,启动共生信号途径,并启动EPR3 受体识别细菌的胞外多糖(EPS),以对该细菌做出容纳与否的反应[5-9]。共生信号途径中SYMRK 结合NFR 形成蛋白质复合物通过核膜上的关键组分如阳离子通道CASTOR、POLLUX/DMI1、环核苷酸门控通道CNGC15 以及核孔蛋白NENA、NUP85 和NUP133,诱发细胞核的Ca2+振荡。依赖Ca2+和钙调素的丝氨酸/苏氨酸蛋白激酶(CCaMK)结合并磷酸化CYCLOPS,促进结瘤起始启动子NIN和ERN1 的表达,最后启动早期结瘤基因ENOD的表达程序,以形成侵染线和根瘤原基[9-12]。侵染线起始于根毛表皮,后穿越外皮层、中皮层细胞的胞质桥向下延展至内皮层细胞增殖后的根瘤原基。侵染线在根瘤原基处释放细菌,植物细胞内吞细菌并形成一层包裹细菌的外周膜(PBM),膜内就是类杆菌的生存空间,植物与根瘤菌通过PBM 进行物质交换以建立共生关系[13-14]。
1.2 根瘤形成发育的影响因素
光、CO2浓度、温度、水分、氮的可用性均可影响根瘤的形成发育。光对结瘤有积极作用,光强、光质以光信号被植株的光敏色素感知,再把信号转移到可移动的蛋白运输到根瘤部分,通过整合根瘤菌诱导的结瘤信号通路调节根瘤形成,同时光合作用产生的碳水化合物为根瘤发育输送能量[15]。CO2浓度、温度、水分、盐分通过影响相关代谢酶的活性、光合产物输出,根系活力、O2渗透等影响根瘤生长发育。氮素对结瘤有负调节效果,通过类黄酮代谢、活性氧积累、植物激素、CEL 小肽等多种途径抑制结瘤[16]。
2 光信号调控根瘤形成
2.1 光信号介导根瘤菌附着
光影响根瘤菌的生长发育及其行为,根瘤菌存在感光蛋白感知光强和波长进而转化为细胞信号。如:细菌光敏色素(bacterio phytochrome)、光氧电压蛋白(light,oxygen,and voltage-domain proteins,LOV)、核黄素蓝光受体(BLUF)和光敏黄蛋白(PYP)[17]。光氧电压蛋白(LOV)是植物、细菌和真菌中光感受器的保守部分,可结合黄酮作为色团并感受蓝光。豌豆根瘤菌RL3841 (Rhizobium leguminosarumbv.viciae3841)的基因组编码一种N 末端LOV 结构域的传感器组氨酸激酶(R-LOV-HK)。R-LOV-HK 在蓝光和UV-A 区有典型的宽LOV 域吸收光谱,并显示缩短的光周期,调节着根瘤菌在光照下的生物膜、胞外多糖及其鞭毛的合成,影响根瘤菌的附着和运动能力,进而影响菌落在植物根际的定殖。试验证明R.leguminosarumbv.viciae3841 接种前的光照培养增加了单株豌豆根瘤和细菌侵染数量,光作为环境因子被R-LOV-HK 感应,间接控制R.leguminosarumbv.viciae3841 的适应性反应和共生效率[18]。光质对根瘤菌生长的影响更加明显,蓝光下百脉根根瘤菌(Mesorhizobium loti)大量减少,对其LOV-HK/PAS 和光解酶相关基因进行信号标签诱变,根瘤的蓝光生长抑制被部分恢复。此外,当用STM 菌株接种Ljcry1A和Ljcry2B(隐花色素表达缺陷)根沉默植株时,结瘤显著增强[19]。
2.2 光信号介导根瘤形成
2.2.1 蓝光诱导从叶片到根的转录因子启动根瘤形成
结瘤受光质影响,受光敏色素系统控制,根和叶片都存在光受体。大量研究表明蓝光激活光感受器协同植物激素参与下胚轴的伸长,并调节根的形成,根和根瘤的形成因存在多种基因重叠[20-26]。最近的研究认为,蓝光是促进根瘤形成的关键条件,敲除和过表达蓝光受体(CRY1b)可分别抑制和促进根瘤的形成[27]。ELONGATED HYPOCOTYL 5 (HY5)是光信号通路中起正向调控作用的转录因子,被光激活后表达,参与光形态建成、光合相关基因的表达,促进叶绿素合成和积累,促进叶片碳水化合物分配和根部硝酸盐吸收。维持不同光环境下植株的碳氮平衡,促进地上地下协同生长。HY5 作为一种从叶片起源的长距离运输信号,可激活根部HY5[28-33]。研究表明HY5 存在4 个仅在大豆叶片中表达的同源基因:GmFT2a、GmFT5a、GmSTF3和GmSTF4,当这些基因在叶片中表达时,根部也同时检测到了新表达的GmSTF3、GmFT2a 和GmFT5a 蛋白,对STF 或FT 转录因子敲除则抑制根瘤形成,过表达则促进根瘤形成。任意敲除其中1 个基因,都会抑制根瘤产生,表明FT和STF基因存在协同调控关系。进一步研究证明GmFT2a 和GmSTF3 从地上移动到地下后,GmSTF3 被GmCCaMK 磷酸化后与GmFT2a 协同促进根瘤基因的表达,表明GmCCaMK-STF-FT 模块共同调控根瘤的形成[27]。还有研究认为蓝光通过抑制根毛中的光敏色素(phytochrome)和隐花色素(cryptochrome)基因表达进而抑制百脉根(Lotus japonicus)根瘤的产生[19]。Ji 等[30]也有类似发现:GmCRY1s 介导蓝光信号,促进了根中HY5 同源基因STF1和STF2的表达,并与STF1和STF2相互作用以稳定STF1/2的蛋白质稳定性,STF1/2 直接抑制结瘤起始信号GmNINa。因此,不同时期和条件下,同一种蛋白可以产生相反的作用,在自养向异养过渡的时候,CRY1s更多作用于下胚轴伸长,而在光合作用启动后,CRY1s侧重于根和根瘤的形成(图1)。
2.2.2 红光调控光诱导的植物激素促进根瘤形成
众所周知,植株通过光敏色素B (PhyB) 感知红光与远红光比率(R/FR),豆科植物结瘤也有PhyB 参与,并整合下游植物激素调节侧根和根瘤形成[21,34]。Suzuki 等[35]认为远红光抑制Lotus japonicus结瘤。高R/FR 下,phyB 通过增加植物茉莉酸异亮氨酸(JAIle)浓度抑制庇荫反应并增强根瘤形成。相反,低R/FR 下,PhyB 失活,JA-Ile 浓度降低并抑制根瘤形成,启动避荫反应以节约能量。Nagata 等[36]也有类似发现:高R/FR 下可显著提高JA 响应基因的表达水平,豆科和非豆科植物根系分泌更多的茉莉酸和独角金内酯,促进了丛枝菌根(AM)共生。Van Gelderen等[37]认为,依赖PhyB 的HY5 通过降低PIN-FORMED3和类AUX1-3 生长素转运蛋白的质膜丰度来促进生长素由上至下极性运输,最终促进侧根发育。最近的研究发现,GA 信号通路的负调节因子DELLA 蛋白,可促进CCaMK-IPD3/CYCLOPS 复合物的形成,促进IPD3 磷酸化,而且能与NSP2-NSP1 复合物相互作用,增强NFs 诱导基因的表达以促进结瘤。而低R/FR 无phyB 活性,GA 合成增加,DELLA 表达下调,最终降低了根瘤数量[38-39](图1)。
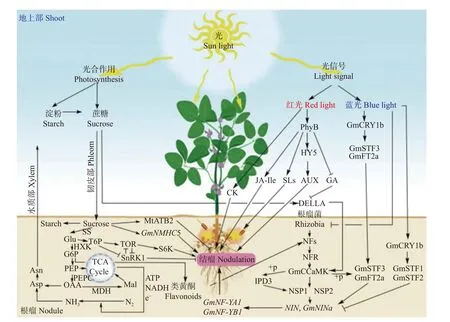
图1 光信号和光合产物调控根瘤形成发育机制Fig.1 Mechanism of root nodule formation and development regulated by light signal and photosynthetic products
3 光合产物调控根瘤生长发育
3.1 光合产物合成根系分泌物以招募根瘤菌
3.1.1 光合产物诱导根瘤菌侵染
光合作用产生的次生代谢物类黄酮可分泌到根际,引诱根瘤菌侵染[41]。研究发现,大豆营养生长期根系的类黄酮主要成分是大豆苷元和染料木素[42],大豆苷元特异性诱导Brady.japonicum nod基因的表达[43],缺氮和结瘤共生使大豆苷元和染料木素分泌量增加约10 倍,类黄酮的分泌量和根瘤干重显著正相关[44]。光合产物为类黄酮的生物合成提供了碳骨架、能量、还原剂。类黄酮的生物合成前后依次有糖酵解、磷酸戊糖、莽草酸、苯丙氨酸、类黄酮代谢途径的参与。糖酵解和磷酸戊糖途径分别提供前体磷酸烯醇式丙酮酸和赤藓糖-4-磷酸,为莽草酸途径合成莽草酸。莽草酸途径合成苯丙氨酸,苯丙氨酸又是类黄酮途径的前体。类黄酮的根系分泌过程有ABC 转运蛋白的参与,需要ATP 提供能量[45-46]。同位素示踪等研究发现,植物存在光合产物的“溢出效应”[47],类黄酮的分泌是营养胁迫的体现,特别在缺氮条件下,光合产物消耗大于同化,可溶性蛋白降低;植株可溶性糖、C/N、苯丙氨酸解氨酶(PAL)活性、苯丙氨酸、总黄酮、总酚含量增加,此时光合产物利用途径更多被导入黄酮等次级代谢物的合成上,为刺激植物生长素极性运输、消除活性氧、根系生长、根瘤菌招募、营养吸收等生存防御反应提供保障[48-50]。
光合作用增强促进植物生长和光合产物的合成,植株群落的多样性增加了地表光合面积和碳截获,根系分泌物随之增加。小麦(Triticum aestivum)蚕豆(Vicia faba)间作增加了蚕豆根系黄酮分泌量和根瘤数量,小麦-玉米(Zea mays)间作使系统类黄酮分泌量提高了2 倍,分离玉米根系分泌物单独处理蚕豆,根中查尔酮-黄烷酮异构酶(CHI)的基因表达量是原来的120%,并诱导结瘤关键基因(ENOD93)表达[51-52]。玉米根系分泌物中也存在类似染料木素、槲皮素等黄酮类物质,通过化感作用刺激蚕豆分泌更多的黄酮化合物以诱导根瘤菌侵染,在根瘤菌正反馈的作用下又会促进黄酮化合物的分泌(图1)。
3.1.2 光合产物为根瘤菌补充碳源
间套作种植模式下,光合产物合成的增加促进了根系营养物质的分泌,如氨基酸、有机酸、糖、蛋白黏液等,这些物质是土壤根瘤菌的重要碳源。玉米根系分泌的黏液中主要有葡萄糖、半乳糖、岩藻糖、甘露糖、阿拉伯糖等糖;半乳糖醛酸、葡萄糖醛酸等糖醛酸,尤其是苹果酸、富马酸、琥珀酸等有机酸是根瘤菌优先利用的碳源,促进了根瘤菌的聚集[53-55]。研究发现,谷类和豆类作物间作可显著提高谷物根系有机酸的分泌量,增加土壤磷、铁、锌等元素的可用性,丰富了细菌群落的多样性,提高了根际根瘤菌和niƒH基因的丰度[56-59]。
3.2 光合产物调控根瘤生长发育
3.2.1 光合产物在根瘤中的代谢过程
根瘤需要消耗大量碳水化合物维持固氮需要的能量和碳骨架,光合产物直接参与根瘤糖代谢。通过光合碳同位素标记和代谢组学分析证实:根和根瘤植物细胞质中主要进行的蔗糖合成/分解、淀粉合成/分解、糖酵解、TCA 循环过程,把光合产物蔗糖转化为根瘤菌直接利用的苹果酸、琥珀酸盐。根瘤菌的遗传学分析也证明,苹果酸、琥珀酸等在PBM和根瘤菌膜上的二羧酸转运体下进入根瘤菌体内,根瘤菌的固氮基因和二羧酸转运体的基因相整合,是共生固氮不可或缺的[60-63]。
光合作用产生的蔗糖(sucrose)先是经过韧皮部运输,通过糖外排转运蛋白(MtSWEET11)到达皮层细胞,在蔗糖合成酶(SS)或转化酶(INV)催化下水解成尿苷二磷酸葡萄糖(6-P-Glu)或葡萄糖(glucose),经过磷酸活化进入糖酵解生成磷酸烯醇式丙酮酸(PEP),再通过磷酸烯醇式丙酮酸羧化酶(PEPC)生成草酰乙酸(OAA),OAA 在苹果酸脱氢酶(MDH)催化下生成根瘤菌呼吸直接需要的苹果酸(Mal)。Mal 由二羧酸转运体(DctA)下进入细菌体内,在低氧条件下进入TCA 循环,产生大量ATP、NADH、e-,在固氮复合物作用下将N2还原为NH3。NH3与Glu (谷氨酸)合成Gln,在谷氨酸合成酶(GOGAT)、α1-抗胰蛋白酶(AAT)催化下形成天冬酰胺(Asn) (或形成酰脲)等经木质部运输到地上部供蛋白合成需要。为保证细菌的高呼吸活性,Mal 还需要通过苹果酸酶(ME)产生丙酮酸(Pyr)以继续生成OAA。菌外TCA 循环的中间产物琥珀酸也源源不断进入菌内刺激TCA 循环运转。在大豆根瘤中可观测到大量的丙二酸(破坏琥珀酸脱氢酶,抑制线粒体呼吸)和琥珀酸积累[2,14,40](图1)。
光合产物在根瘤中的分解代谢由SS、PEPC、MDH 3 个关键酶调节[2]。SS 在植株广泛分布,在大豆中额外补充糖、谷氨酰胺和尿囊素可上调SS 的表达,在根瘤中干扰其表达会导致蔗糖、果糖的浓度下降[2,64-65]。蔗糖磷酸合成酶(SPS)可以分解淀粉合成蔗糖,也可以把蔗糖转化为淀粉运送到根瘤活性中央区。SS 和SPS 都有利于维持根瘤共生后的蔗糖供应[66]。叶片中的PEPC 有助于叶肉细胞固定和存储CO2,给维管束细胞中的卡尔文循环提供原料,而根瘤中的PEPC 把糖酵解途径中的PEP 导向另一个分枝:氮代谢。其催化生成的OAA 可以给氨基酸合成4C 骨架,Asn、Asp (天冬胺酸)所需的C 有30%是根瘤PEPC固定的。停止PEPC 表达后,根瘤中乙炔还原活性,SS 和AAT 活性均显著降低,蔗糖、琥珀酸盐、天冬酰胺、天冬氨酸和谷氨酸的含量也有所下降。根瘤PEPC 被6-P-Glu 和磷酸三糖激活,被L-malate 和氮同化产物Asp 抑制,减少光合作用和光合产物输入,长暗处理和环剥降低了PEPC 的磷酸化水平,增加了PEPC 对L-malate 抑制的敏感性,恢复光照后PEPC 的去磷酸化水平明显降低。减少光合作用不会降低PEPC的最佳活性,PEPC 的磷酸化水平和活性调节依赖于就近的光合产物调节[67-68]。豆类根瘤中存在独特的根瘤增强型MDH (Ne-MDH),其转录水平量随着根瘤发育阶段增强。Ne-MDH 对Mal 的米氏常数(Km)远大于对OAA 的,且具有较高的催化常数(Kcat),表明Ne-MDH 催化的反应更有利于形成Mal[69-70]。
3.2.2 光合产物作为糖信号启动根瘤形成
光合产物中长距离运输的蔗糖及其分解产物葡萄糖可作为信号直接促进DELLA 蛋白表达基因上调,诱导结瘤共生信号,还可以通过促进侧根的发育以实现结瘤[38-39]。MADS-box基因被认为有调控侧根发育的功能,它的一种MIKC 型转录因子GmNMHC5,与拟南芥(Arabidopsis thaliana)中响应硝酸盐调控侧根生长的关键转录因子AGL17有高度相似的氨基酸序列,GmNMHC5和AGL17均在根系被高度表达。研究证明,转基因大豆中GmNMHC5的过表达显著促进了侧根发育和结瘤,外源蔗糖的添加可上调GmNMHC5[71]。在衰老期,蔗糖还可以加速根瘤衰老,蒺藜苜蓿(Medicago truncatula)参与根瘤衰老调节的转录因子MtATB2 和AtATB2 一样受到蔗糖和光的诱导,加入蔗糖可促使MtATB2 转录水平升高,根系和根瘤数量均减少[72]。蔗糖分解产生的葡萄糖也是一种广泛的糖信号,己糖激酶(HXK)是糖酵解第一步的限速酶,也是葡萄糖受体,参与葡萄糖信号转导,尤其对进入代谢的糖通量敏感[73-74]。蔗糖分解为葡萄糖后,经过HXK 催化后形成高活性的6-磷酸葡萄糖(G6P)可抑制蔗糖非发酵相关激酶1(SnRK1)活性。SnRK1 作为营养胁迫感应器并激活相应转录因子,通过磷酸化控制14-3-3 蛋白结合,同时抑制硝酸还原酶(NR)和蔗糖磷酸合成酶(SPS)的活性,进而延迟根瘤发育[75]。同时,G6P 在海藻糖6-磷酸合酶(TPS)和T6P 磷酸酶(TPP)的活性参与下通过糖代谢的另一个分支海藻糖途径,产生二糖海藻糖(Tre) (根瘤菌重要的渗透保护剂)。Tre 的前体T6P 是信号物质和生长调节剂,在低生理浓度下对SnRK1 进行抑制,进而促进根系发育和结瘤[76-77]。TOR 激酶是与SnRK1 拮抗作用的营养信号感受和生长发育调节器,在大豆根毛侵染线和成熟根瘤细胞内大量转录,有助于活性氧的减少,对植物与根瘤菌形成共生关系有重要贡献,TOR 可响应糖酵解强度或直接响应G6P 以激活根分生组织,为细胞壁合成提供中间代谢物和碳能量,还能感应自上而下极性运输的生长素信号,激活核糖体蛋白激酶S6K,磷酸化核糖体蛋白RPS6-1/2,促进侧根发育和根瘤菌诱导的皮层细胞分裂[78-80](图1)。
3.2.3 光合产物作为能源促进根瘤生长发育
根瘤生长发育需要的能源来自于植株光合作用产生并运输的蔗糖[2,81]。Fujikake 等[82]发现蔗糖处理可以显著增加根瘤直径,对植株进行照光也是如此,光照时间越长,根瘤直径增加越快;光照和蔗糖处理可以解除根瘤生长的硝酸盐抑制现象。还有研究表明,大豆生根培养基外源补充蔗糖可以显著提高根瘤菌在酸性、干旱、高温条件下的耐逆性,其细菌的渗透保护性物质:海藻糖、甘油都显著提升,并且大豆的结瘤数量增加[83]。把结瘤的苜蓿(Medicago sativa)移除地上部7 d 后,其根瘤的蔗糖、苹果酸和α-酮戊二酸含量降低,地上部碳同化能力的削弱,抑制了根瘤的生长。根瘤的C13含量在地上部移除7 d后显著增加,根部存储的碳水化合物就近分配给根瘤以维持根瘤的活性,随着时间的推移,根瘤中被标记的蔗糖含量降低,C13的氨基酸含量增加,证明了这部分C13从蔗糖移到了氨基酸[84]。高镁供应下,叶绿素含量增加,光合作用增强,蔗糖合成酶和磷酸合成酶活性增强,大豆叶片的蔗糖和淀粉积累减少,根瘤中的积累量增加,在不改变根瘤结构的情况下促进了根瘤数量和直径的增长。因此,镁通过改变光合产物的供应促进了根瘤的生长[85]。
促进光合产物向根瘤的分配将有利于根瘤的生长,但是光合产物对根瘤的促进作用存在限度。有研究表明增加环境CO2的浓度,光合产物增加,鹰嘴豆(Cicer arietinum)的根伸长,根系类黄酮分泌无显著性差异,根瘤形成个数先升高后降低。CO2的升高首先促进了根的伸长(伸长明显高于增粗的幅度),进而提高根瘤个数,继续增加CO2浓度,根瘤个数没有增加[86]。有人提出以下解释:在能量供应充足情况下,植株细胞代谢增强,自由基产生较多。高浓度CO2调控初级代谢进入衰老状态,且根瘤的固氮能力和蔗糖利用效率并不一定随着碳供应提高,多余的糖可在根瘤储存为淀粉[87]。高CO2浓度状态下植株的叶绿素含量和NBI (叶绿素和类黄酮的比值)不增反降,可知根瘤固氮能力和植株的氮素水平并未提升[88]。光合产物不能无限促进根瘤生长和固氮能力的根本原因是:光合产物经过蔗糖分解、糖酵解形成的PEP 有两条去路,一是直接(结合1 分子CO2)形成OAA 作为同化铵的骨架,二是继续脱羧(释放1 分子CO2)形成乙酰辅酶A (乙酰CoA),与OAA结合汇入TCA循环,根瘤从PEP 开始到TCA 需要丢失3 分子CO2,这意味着光合产物对根瘤菌呼吸的能量供应存在碳损失,地上部运输的糖不能完全满足根瘤对有机酸的旺盛需求,因为有机酸的消耗更快。为了保证铵同化的高效性,满足植物生长需要,减少对固氮酶的抑制,需要额外补充较高的CO2浓度,以提供源源不断的Mal 和OAA 去分别维持根瘤菌的能量需求和根瘤细胞的氮同化。直接增加根瘤细胞的CO2供应可显著改善根瘤活性,根瘤明显变红、变大,单颗根瘤干物质平均增重5 倍,根瘤数量也增加近1 倍,同时根瘤中的有机酸和氨基酸含量、木质部的Asn 含量均有显著提升[89]。
4 总结和展望
豆科植株和根瘤菌经过长期的共同进化形成共生互惠关系,植物给根瘤菌固氮需要的微氧环境和碳营养,而根瘤则成了专一输出铵的耗能细胞器。植株子叶出土后,光以光形态建成和光合作用的方式控制根瘤形成发育的各个方面。当光照条件较好时,植物的碳同化引发了氮需要,而在土壤NO3-/NH4+不足的情况下,豆科植物开始分泌类黄酮吸引根瘤菌。但是具体需要形成多少根瘤,还需要经过地上部叶片的调控,叶片通过光敏色素感知光质条件以调控根瘤的形成,根据体内糖代谢状态提供能量供应。根和根瘤的形成整合了大量相似的转录途径,属于此消彼长的关系,无论是直接吸收土壤的氮还是通过根瘤菌固定空气中的氮,都是植物高效利用资源,减少资源浪费的一种智慧反应。
全球气候变暖,作物生产的风险性增加,作物多样性种植对稳定产量,抵御逆境做出了巨大贡献,但归根结底是单位土地面积光截获量的提升。多样性种植[如豆科和禾本科(Poaceae)植物的间套作]增产稳产的同时会加大氮素的投入,势必带来更大的经济成本和环境压力。光比氮对植株的初级生产力影响更大,利用豆科植物间套作其他作物可以实现农田氮素和光能的双增益。关于豆科植物如何响应硝酸盐调控结瘤的分子机制研究已有很多,但光及光合产物如何调控根瘤形成发育的转录、代谢途径还不太明确。例如光合产物如何影响氮的转运和同化以调节植物氮水平进而决定是否形成根瘤? 根瘤形成以及固氮过程中的碳成本和氮收益? 光信号和光合产物是否也参与到结瘤自动调节途径(AON)调控中? 探究这些问题,有助于系统阐释豆科植物植株和群体光合产物利用和固氮机理,丰富豆科植物结瘤固氮理论,为间套作减肥增效技术研发提供理论依据。
猜你喜欢
山东冶金(2022年1期)2022-04-19
中国南方果树(2022年1期)2022-01-28
生态科学(2021年5期)2021-11-13
湿法冶金(2021年5期)2021-10-14
农技服务(2021年7期)2021-09-24
上海金属(2021年4期)2021-07-28
上海金属(2020年6期)2021-01-04
河北果树(2020年1期)2020-02-09
植物营养与肥料学报(2019年11期)2019-12-13
——固氮微生物
新疆农垦科技(2016年3期)2016-02-20
