
多异瓢虫在3种豆科杂草上的种群动态及对其趋性行为反应
2023-02-03 10:22宋冰梅程宛楠潘洪生
植物保护 2023年1期
姜 岩, 陈 鑫,2, 宋冰梅,3, 张 宇, 程宛楠, 潘洪生*
(1.新疆农业科学院植物保护研究所,国家植物保护库尔勒观测实验站, 乌鲁木齐 830091; 2.新疆农业大学农学院, 棉花教育部工程研究中心, 乌鲁木齐 830052; 3.新疆大学生命科学与技术学院, 新疆生物资源和基因工程重点实验室, 乌鲁木齐 830046; 4.中国农业科学院植物保护研究所, 植物病虫害生物学国家重点实验室, 北京 100193)
棉花是全球纺织行业的主要材料[1],我国则是全球棉花最大生产国和加工国之一[2]。2021年,新疆棉花播种面积为250.6万hm2、产量为512.9万t,分别占全国棉花种植面积的82.76%和棉花产量的89.50%[3],已经连续27年居全国首位[1]。因此,棉花在新疆经济发展过程中有着举足轻重的作用,是关系到民生的战略性物资[4]。蚜虫是新疆棉田三大害虫之一[5],其通过刺吸式口器取食棉花植株汁液,造成叶片卷曲变形、光合作用降低[6],同时能够传播包括黄瓜花叶病毒(cucumber mosaic virus)[7]、黄病毒(cucurbit aphid-borne yellows virus)[8]和玉米矮花叶病毒(maize dwarf mosaic virus)[9]等多种植物病毒[10]。蚜虫种群密度高、个体小,繁殖周期短且繁殖能力强[11],一旦在棉田暴发会造成非常严重的经济损失[12]。目前,对于蚜虫的防治很大程度上依赖化学药剂,然而其抗药性不断增强[13],亟须开发具有持续性生态效应的绿色防控手段,其中保护和利用自然天敌成为实现棉田蚜虫绿色防控的重要途径[14-15]。多异瓢虫Hippodamiavariegata在新疆棉区发生较早[16]、种群数量大,能够捕食多种害虫,且捕食量大[17-18],是新疆棉区蚜虫的优势捕食性天敌[19]。因此,保护和利用多异瓢虫是实现新疆棉区蚜虫生物防治的一种重要措施。
农业生态系统指以种植农作物为主的生态系统,主要包括作物生境和周围的非作物生境[20]。大量研究表明,农业生态系统中的非作物生境能够为天敌提供替代猎物和避难所[21-22],其中的边缘效应和溢出效应也能够促进天敌的迁移活动,调控天敌的控害功能,减少虫害的发生和再猖獗,对保育天敌和防控害虫均有着重要的意义[23]。利用非作物生境保育天敌进行生物防治的前提是系统地调查非作物生境中天敌与害虫的发生及种群动态,详尽地研究天敌定位非作物生境中植物的过程及线索,综合评价其对天敌及害虫的作用[24]。甘草Glycyrrhizauralensis、骆驼刺Alhagicamelorum和苦豆子Sophoraalopecuroides是新疆棉区非作物生境中常见的3种豆科杂草[25-26]。本研究首先对棉田田埂这3种豆科杂草上多异瓢虫与棉黑蚜Aphisatrata种群动态进行了系统调查,同时利用Y形嗅觉仪测试多异瓢虫对3种豆科杂草不同处理的趋性行为反应,为合理利用非作物生境保育多异瓢虫,实现新疆地区棉田蚜虫的绿色防控提供重要的科学依据。
1 材料与方法
1.1 供试昆虫与植物
试验所用多异瓢虫成虫及棉黑蚜均采自国家植物保护库尔勒观测实验站(新疆巴州库尔勒市和什力克乡库勒村,41.75°N, 85.81°E)棉田周边田埂的甘草、骆驼刺和苦豆子上。其中多异瓢虫采用实验室连续饲养多代的桃蚜Myzuspersicae饲养于20 cm×10 cm×6 cm 的塑料养虫盒中,室内饲养环境条件为温度(25±2)℃,相对湿度(60±5)%,光周期L∥D=14 h∥10 h。取第二代5~8日龄的多异瓢虫雌雄成虫,饥饿24 h后用Y 形嗅觉仪测试。
3种供试豆科杂草甘草、骆驼刺和苦豆子选自国家植物保护库尔勒观测实验站周边不使用任何农药的棉田田埂上。供试植物发芽后,每种杂草用2个3 m×1 m×2 m的80目防虫网罩住,防止植食性昆虫和螨类为害。其中一个为健康植株处理,另外一个为棉黑蚜为害植株处理,即每种杂草进入苗期后、株高约50 cm时,用剪刀剪取田间自然发生的棉黑蚜,接在棉黑蚜为害植株处理罩笼中的每种杂草上,每株接棉黑蚜400头左右,为害48 h后从顶端截取长20 cm含有200头左右棉黑蚜的植株用Y 形嗅觉仪测试。
1.2 3种豆科杂草上多异瓢虫和棉黑蚜的种群动态
试验在国家植物保护库尔勒观测实验站开展,分别选取棉田田埂上宽约1 m,长度20 m的甘草、骆驼刺和苦豆子带各3个小区,每个小区调查5点,每点调查面积为1 m2(1 m×1 m)。5月中旬至8月中旬,采用目测法分别调查并记录其上多异瓢虫成虫、幼虫及棉黑蚜的种群数量,每7 d调查1次,同时记录3种豆科杂草的生育期。试验期间3种豆科杂草带均不施用任何化学农药。
1.3 多异瓢虫对3种豆科杂草的趋性行为反应
参考 Pan 等[27]的方法,利用Y形嗅觉仪测试多异瓢虫对3种豆科杂草不同处理的趋性行为反应。试验共设置4个处理:空白、棉黑蚜(200头)、健康植株和棉黑蚜为害48 h的植株。试验用Y形嗅觉仪由内径为3.0 cm的无色透明玻璃制成,其基部(C臂)及两臂(A、B臂)长均为15 cm,两臂夹角60°。Y形嗅觉仪放置在100 cm×100 cm×60 cm的暗室中,嗅觉仪上方40 cm处挂有1个40 W的日光灯(光强700 lx)。基部与瓢虫成虫的释放管相连接,两臂用Teflon管分别连接放置不同试验处理的味源瓶。用大气采样仪作为气流动力系统,气流进入味源瓶前经过活性炭过滤,以及蒸馏水加湿,气体流速设定为0.4 L/min。
将单头多异瓢虫成虫释放入Y形管C臂观察,当瓢虫处于C管长1/2处开始计时。根据前期测试结果,按照下述标准进行评判:当瓢虫越过A或B臂3 cm处且在此区域内停留5 s以上时,记为做出选择;当放入瓢虫3 min后无明显选择趋向,则记为无反应。每头瓢虫只测试1次,每测试2头交换A、B两臂位置及气流方向,每测试4头更换Y形管,每测试30头更换不同处理中的植物材料。每处理雌雄瓢虫成虫各测试60头。每天测试完成后用丙酮清洗Y形管、味源瓶及连接胶管。

1.4 数据统计与分析
采用Microsoft Excel 2003和SPSS 20对所有数据进行统计分析。3种豆科杂草上棉黑蚜和多异瓢虫种群数量均换算为每平方米虫量,用每平方米多异瓢虫成虫和幼虫总数与棉黑蚜进行相关性分析,通过皮尔逊相关系数(r)来描述相关程度:0≤|r|≤0.3为微弱相关;0.3<|r|≤0.5为低度相关;0.5<|r|≤0.8为显著相关;0.8<|r|<1为高度相关;|r|=1为完全线性相关[28]。Y形嗅觉仪测试中,成虫在两处理间是否呈假设H0为50∶50的理论分布用χ2检验,计算χ2值和相应的显著性水平P值,未做出选择的多异瓢虫不列入统计分析内。
2 结果与分析
2.1 3种豆科杂草上多异瓢虫和棉黑蚜的种群动态
甘草上,多异瓢虫成虫和幼虫均在5月下旬至6月下旬进入盛发期,并于6月7日种群数量达到最大值(成虫:2.80头/m2;幼虫:1.80头/m2),然后逐渐下降,7月12日种群数量趋近于零;棉黑蚜种群发生动态与多异瓢虫种群趋势相似,即多异瓢虫种群数量较多时,棉黑蚜种群数量也较多(图1a)。同时,在整个调查期间甘草上多异瓢虫与棉黑蚜种群数密度高度相关,相关系数为r=0.923(P<0.001)。
骆驼刺上,多异瓢虫成虫和幼虫也于5月下旬至6月下旬进入盛发期,并分别于6月14日和6月21日种群数量达到最大值(成虫:2.19头/m2;幼虫:1.45头/m2),然后逐渐下降,7月19日种群数量趋近于零;棉黑蚜种群发生动态与多异瓢虫种群趋势相似,且棉黑蚜的盛发期(5月17日-6月7日)、达到种群数量最大值日期(6月7日)和种群趋近于零日期(7月12日)均较多异瓢虫提前7~14 d(图1b)。并且,在整个调查期间骆驼刺上多异瓢虫发生量与棉黑蚜种群数量显著相关,相关系数为r=0.774(P=0.001)。
苦豆子上,多异瓢虫成虫和幼虫均在5月下旬至6月下旬进入盛发期,并于6月21日种群数量达到最大值(成虫:1.35头/m2;幼虫:0.93头/m2),然后逐渐下降,7月12日种群数量趋近于零;棉黑蚜种群发生动态同样与多异瓢虫种群趋势相似,且棉黑蚜的盛发期(5月17日至6月14日)和达到种群数量最大值日期(6月14日)较多异瓢虫提前7 d左右(图1c)。并且,在整个调查期间苦豆子上多异瓢虫发生量与棉黑蚜种群数量高度相关,相关系数为r=0.839(P<0.001)。

图1 3种豆科杂草上多异瓢虫和棉黑蚜的种群动态
2.2 多异瓢虫对3种豆科杂草的趋性行为反应
多异瓢虫雌雄成虫对甘草不同处理的趋性行为反应结果基本一致。具体表现在:多异瓢虫雌雄成虫对棉黑蚜的选择率均极显著大于空白对照(雌:χ2=9.60,P=0.002;雄:χ2=6.90,P=0.009);对棉黑蚜的选择率均显著大于健康甘草植株(雌:χ2=3.95,P=0.047;雄:χ2=4.90,P=0.027);对棉黑蚜为害的甘草植株的选择率均显著大于棉黑蚜(雌:χ2=4.57,P=0.033;雄:χ2=4.27,P=0.039);对棉黑蚜为害的甘草植株的选择率显著大于健康甘草植株(雌:χ2=6.67,P=0.010;雄:χ2=5.59,P=0.018)。综上所述,多异瓢虫雌雄成虫对甘草不同处理的趋性行为反应排序为:棉黑蚜为害的甘草植株>棉黑蚜>健康甘草植株>空白(图2a)。
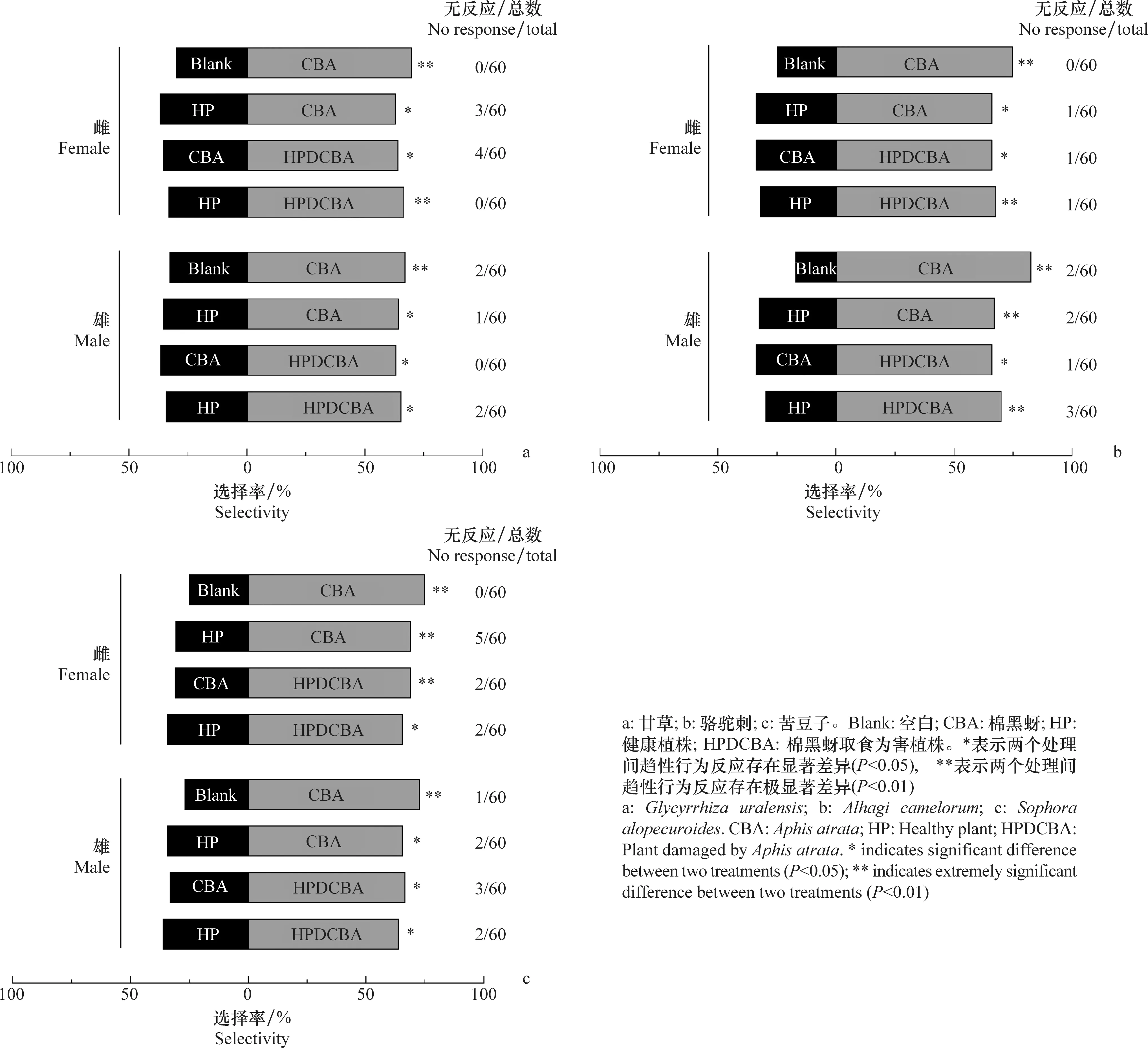
图2 多异瓢虫雌雄成虫对3种豆科杂草不同处理间的趋性行为反应
多异瓢虫雌雄成虫对骆驼刺不同处理的趋性行为反应结果也基本一致。多异瓢虫雌雄成虫对棉黑蚜的选择率均极显著大于空白对照(雌:χ2=15.00,P<0.001;雄:χ2=24.90,P<0.001);对棉黑蚜的选择率均显著大于健康骆驼刺植株(雌:χ2=6.12,P=0.013;雄:χ2=6.90,P=0.009);对棉黑蚜为害的骆驼刺植株的选择率均显著大于棉黑蚜(雌:χ2=6.12,P=0.013;雄:χ2=6.12,P=0.013);对棉黑蚜为害的骆驼刺植株的选择率极显著大于健康骆驼刺植株(雌:χ2=7.48,P=0.006;雄:χ2=9.28,P=0.002)。因此,多异瓢虫雌雄成虫对骆驼刺不同处理的趋性行为反应排序为:棉黑蚜为害的骆驼刺植株>棉黑蚜>健康骆驼刺植株>空白(图2b)。
多异瓢虫雌雄成虫对苦豆子不同处理的趋性行为反应结果基本一致。多异瓢虫雌雄成虫对棉黑蚜的选择率均极显著大于空白对照(雌:χ2=15.00,P<0.001;雄:χ2=12.36,P<0.001);对棉黑蚜的选择率均显著大于健康苦豆子植株(雌:χ2=8.02,P=0.005;雄:χ2=5.59,P=0.018);对棉黑蚜为害的苦豆子植株的选择率均显著大于棉黑蚜(雌:χ2=8.35,P=0.004;雄:χ2=6.33,P=0.012);对棉黑蚜为害的苦豆子植株的选择率显著大于健康苦豆子植株(雌:χ2=5.59,P=0.018;雄:χ2=4.41,P=0.036)。总的来说,多异瓢虫雌雄成虫对苦豆子不同处理的趋性行为反应排序为:棉黑蚜为害的苦豆子植株>棉黑蚜>健康苦豆子植株>空白(图2c)。
3 结论与讨论
本文通过田间系统调查发现,早春田埂3种豆科杂草上多异瓢虫种群密度较高,同一时期其上棉黑蚜种群密度也较高,且两者之间具有显著的相关性;Y形嗅觉仪测试表明,多异瓢虫雌雄成虫均显著偏向选择棉黑蚜为害后的3种豆科杂草。这与其他对非作物生境[22,29-30]和瓢虫行为反应[31-34]的研究结果相似,即非作物生境通过提供替代猎物和栖息地来有效保育天敌,保证其后期向作物生境迁移,行使生物控害的功能[23,35],同时被害虫为害的植株释放的挥发性物质对瓢虫的趋性选择行为有十分重要的作用[36-37]。
本研究涉及的3种豆科杂草均为多年生草本植物[38-40],出苗较早[41-43],其上棉黑蚜的发生也较早,本课题组前期调查研究表明,多异瓢虫每日捕食棉黑蚜最高可达157.30头[17]。因此,早春这3种豆科杂草上的棉黑蚜为多异瓢虫提供了丰富的食物资源,对其具有重要的保育作用。其次,大量研究表明,花粉、花蜜可以维持捕食性天敌的生长发育[44-46],甚至在促进其性成熟、延长寿命、提高生殖力及寄主植物的适合度等方面起到重要的作用[47-49]。而且,蚜虫及植物分泌的蜜露中同样含有捕食性天敌所需的糖类、必需氨基酸及矿物质[50-51],也可以为捕食性天敌提供相应的营养成分[52-54]。6月上旬3种豆科杂草均进入花期,其上的花粉和花蜜是否也能够成为多异瓢虫的替代食物资源,以及3种豆科杂草上棉黑蚜分泌的蜜露对多异瓢虫生长发育和繁殖能力的影响均有待进一步研究。
植物挥发物在捕食性瓢虫定位猎物所在寄主植物的过程中起着特别重要的作用[55-58]。因此,我们猜测植物挥发物在多异瓢虫对3种豆科杂草的趋向选择行为中同样具有重要的作用。为了验证这一猜想,我们通过Y形嗅觉仪测试发现多异瓢虫雌雄成虫对3种豆科杂草不同处理间的趋向选择行为反应相似,均为:棉黑蚜为害植株>棉黑蚜>健康植株>空白(图2)。这与多异瓢虫对棉蚜Aphisgossypii为害的黄瓜Cucumissativus植株[59]以及异色瓢虫Harmoniaaxyridis对蚜虫为害的多种作物[37]的行为反应结果均类似。因此,我们可以在后续的研究中继续探究棉黑蚜为害豆科杂草释放挥发性物质的种类和功能,了解多异瓢虫趋向选择行为利用的化学信息线索,同时进一步通过挥发物合理配比,研发多异瓢虫引诱剂,调控作物系统中天敌的发生时间及种群数量,为有效利用多异瓢虫进行蚜虫绿色防控奠定理论基础和提供技术支持。
综上所述,新疆棉区田埂3种豆科杂草上的棉黑蚜对多异瓢虫早春种群具有重要的保育作用,棉黑蚜取食为害后的3种豆科植物挥发物可能在多异瓢虫成虫对其趋向选择过程中发挥关键作用。本研究为利用非作物生境保育多异瓢虫及促进其生物控害作用奠定重要的理论基础。
猜你喜欢
上海农业科技(2022年2期)2022-05-13
科学中国人·上半月(2021年4期)2021-07-20
阅读与作文(小学高年级版)(2020年2期)2020-05-21
领导文萃(2018年3期)2018-03-12
北方人(2018年3期)2018-03-08
农业环境科学学报(2017年2期)2017-03-20
农业知识(2016年46期)2017-01-13
红蜻蜓·低年级(2016年4期)2016-11-19
草食家畜(2012年2期)2012-03-20
右江民族医学院学报(2012年4期)2012-01-26
