
棉铃虫围食膜中Bt抗性相关蛋白的分离与鉴定
2023-02-03 10:22李怡萍刘少凯袁向群
植物保护 2023年1期
李怡萍, 刘少凯, 袁向群,
(1.西北农林科技大学植物保护学院, 农业农村部西北黄土高原作物有害生物综合治理重点实验室, 杨凌 712100; 2.中国农业科学院植物保护研究所, 植物病虫害生物学国家重点实验室, 北京 100193)
棉铃虫Helicoverpaarmigera是鳞翅目夜蛾科的一种世界性重大农业害虫,其分布范围广泛,在我国西北内陆棉区、长江流域棉区和黄河流域棉区这三大主要棉区均有分布。棉铃虫寄主范围广,能取食包括棉花、玉米、大豆等多种农作物及经济作物,具有环境适应能力强、产卵量大、迁飞距离远等特点[1-2]。在20世纪60年代至70年代,对棉铃虫的防治均以化学防治为主,其中有机磷类、有机氯类等多种化学杀虫剂被先后使用。但是随着杀虫剂的大量使用,20世纪80年代末至90年代初期,棉铃虫快速对滴滴涕、硫丹、拟除虫菊酯类等常规杀虫剂产生了抗性,田间防治基本失效,加之适宜的气候条件等原因,棉铃虫在我国黄河流域和长江流域棉区连年暴发成灾,造成了极大的损失,直接经济损失超过百亿元[3-4]。
1997年,我国开始商业化种植转Bt(Bacillusthuringiensis,苏云金芽胞杆菌)基因的抗虫棉[5]。转Bt基因棉的种植,以及Bt生物杀虫剂的使用,不仅有效防控了棉铃虫等靶标害虫,还减少了很多化学杀虫剂的使用[4]。我国目前种植的转基因棉花均为表达cry1Ac基因或cry1Ac+cry1Ab融合基因的Bt棉,随着我国棉花种植空间不断向以新疆为主的西北棉区集中,长期使用单价cry1A家族的Bt棉必然会存在田间抗性进化的隐患[4]。
目前研究表明抗性产生的最主要原因是昆虫中肠的刷状缘膜囊泡(brush border membrane vesicles,BBMV)上与Bt杀虫蛋白结合的受体的改变或突变引起的[6-8]。昆虫取食Bt Cry1A类全长蛋白后,在中肠碱性环境中被胰蛋白酶或胰凝乳蛋白酶水解,N端去除40个左右的氨基酸,C端去除500个左右的氨基酸,原蛋白由130 kD左右被活化为65 kD左右有杀虫活性的核心片段。活化的蛋白片段穿过围食膜(peritrophic membrane,PM),进一步与中肠BBMV上受体结合,发挥作用。但是Bt杀虫蛋白只有穿过中肠围食膜才能达到中肠上皮细胞的作用位点与受体结合,且研究发现昆虫围食膜上也可能存在与Bt杀虫蛋白结合的受体蛋白,因此,昆虫围食膜蛋白的改变也可能引起Bt抗性的产生[9-11]。
1762年在鳞翅目幼虫中发现围食膜结构,1890年正式命名[12]。围食膜是中肠前端一直延伸至后肠的一层厚薄均匀的无色透明的非细胞结构薄膜,是由中肠细胞分泌形成的,主要由蛋白质、黏多糖和几丁质组成,是昆虫抵御病原微生物入侵及有害物质的第一道天然保护性屏障,具有保护中肠上皮细胞、阻止病原物入侵等防御功能[13-17]。有关围食膜蛋白的组成研究一直是学者们关注的热点,张严峻等[18]使用SDS-PAGE技术在棉铃虫围食膜中发现了16种蛋白,Campbell等[15]通过质谱技术鉴定出了41个棉铃虫幼虫围食膜蛋白质。随着生物技术的发展,梁振普等[17]通过LC-MS/MS技术鉴定了169个棉铃虫围食膜蛋白质。其中也包括了碱性磷酸酶2(alkaline phosphatase 2)、氨肽酶N1(aminopeptidase N1)、ATP合成酶β亚基(ATP synthase subunit beta)、V型质子ATP酶D2亚基(V-type proton ATPase subunit D2)等,这些蛋白是Bt杀虫蛋白结合的受体,与Bt抗性产生有一定的关系。
然而,对棉铃虫围食膜蛋白与Bt抗性的关系研究甚少,本研究以棉铃虫抗、感品系围食膜为对象,采用蛋白质分离和配体杂交(ligand blot)技术,研究棉铃虫围食膜上与Bt-Cry1Ac存在结合能力有差异的蛋白,通过质谱鉴定和生物信息学分析等技术,鉴定可能参与Bt杀虫蛋白抗性进化的差异蛋白。研究有助于理解棉铃虫围食膜蛋白与Bt杀虫蛋白之间的作用,为进一步明确Bt抗性机制、制定合理的Bt抗性治理策略提供理论依据。
1 材料与方法
1.1 供试昆虫及饲养条件
敏感品系棉铃虫(96S):于1996年采自河南省新乡棉田,室内用人工饲料饲养至今,从未接触任何杀虫剂。棉铃虫Bt抗性品系(BtR):在室内用Cry1Ac蛋白筛选了175代,相对抗性倍数为3 500倍,筛选方法参见史艳霞等[19]方法。饲养条件: 温度(27±2)℃,相对湿度(75±10)%,光周期为L∥D=14 h∥10 h。幼虫在指形管(直径2 cm,长度15 cm)中用人工饲料[20]饲养,成虫在产卵笼(直径29.5 cm,高35 cm)中用10%蜂蜜水饲养。上述所有棉铃虫品系均由中国农业科学院植物保护研究所棉花害虫组馈赠。
1.2 供试Bt菌株及抗体
供试BacillusthuringiensisHD-73菌株和Cry1Ac抗体均由中国农科院植物保护研究所生物技术组馈赠。二抗为山羊抗兔,购自北京中杉金桥公司。
1.3 棉铃虫围食膜蛋白的提取分离
1.3.1围食膜样品的获取
分别取抗、感品系健康的5龄棉铃虫幼虫,置于冰上10 min,剖取围食膜,用冷却的生理盐水(0.7% NaCl)冲洗去掉里面的食物,在滤纸上快速吸干多余液体,每1条围食膜放入1个EP管中,液氮中快速冷却,使用FD5508冷冻干燥机(美国SIM公司)冷冻干燥48 h后,置-80℃冰箱保存备用。
1.3.2棉铃虫围食膜蛋白的分离纯化
参考Campbell等[15]和李怡萍等[21]的方法,称取冷冻干燥的抗、感品系棉铃虫围食膜样品各1 mg,进行无水三氟利克酸(anhydrous trifluoromethanesulfonic acid,TFMS)处理,用GlycoPrfile TM IV,Chemical Deglycosylation Kit(美国Invitrogen公司)进行总蛋白的提取,再用2D Clean-Up Kit(美国GE Biosciences公司)纯化提取的总蛋白。
1.3.3棉铃虫围食膜蛋白质含量的测定
取1.3.2中纯化的抗、感品系围食膜蛋白沉淀(指初始取的冷冻干燥围食膜为1 mg,经TMFS处理与纯化后获得的总蛋白沉淀),分别用200 μL上样缓冲液IEF溶解。12 000 r/min、4℃,离心5 min(5145D台式离心机,德国Eppendorf Corporation),除去不溶物质和泡沫,取上清液,用2D Quant Kit(美国GE Biosciences公司)测定蛋白浓度。根据测定的蛋白浓度计算出1 mg干燥的围食膜样品中蛋白质总重和蛋白质含量。每样品重复3次,测定数据使用DPS 9.50中独立样本t测验进行差异显著性分析。
1.3.4棉铃虫围食膜蛋白的NuPAGE电泳
取1.3.2中纯化的抗、感品系围食膜蛋白沉淀,使用NuPAGE 4%~12% Bis-Tris Mini Gel预制梯度胶(美国Invitrogen公司)及NuPAGE电泳槽(美国Stragene公司)进行电泳,电泳电压为200 V,电泳时长为50 min。电泳完成后进行考染。
1.3.5棉铃虫围食膜蛋白与活化Cry1Ac的结合能力检测
采用1.3.4中的方法对抗性和敏感品系的围食膜蛋白进行电泳检测。上样之前要测其蛋白浓度,使二者保持相同的蛋白浓度。电泳结束后的胶块不染色,直接进行ligand blot,先加活化的Cry1Ac,再分别加一抗Cry1Ac抗体和二抗山羊抗兔,最后使用Eagle Eye System凝胶成像仪系统(美国Stragene公司)进行曝光检测。
1.4 抗、感品系棉铃虫围食膜差异蛋白的鉴定分析
1.4.1棉铃虫围食膜蛋白的蛋白胶内酶解及质谱鉴定
在考染胶上用挖胶笔挖出选定的有显著差异的蛋白条带,尽可能避免切到胶以外的部分,放入离心管中。蛋白胶内酶解和质谱鉴定由华大蛋白公司完成。
1.4.2棉铃虫围食膜蛋白的质谱数据生物学分析
将质谱鉴定过程中,由MALDI-TOF(基质辅助激光解吸电离离子源和飞行时间质量分析器联用)质谱仪(新加坡ABSciex公司)获得肽指纹图谱(peptide-mass fingerprint,PMF),用MASCO(http:∥mascot.proteomics.com.cn/)Aldente软件搜索NCBI nr和Swiss-Prot数据库,并参考了中国农业科学院植物保护研究所棉花害虫组的棉铃虫cDNA文库数据,进行BLAST,比对PMF的结果。比较数据库中相应多肽序列的质量数及其MS/MS图谱的相关性,由中国农业科学院植物保护研究所生物技术组束长龙老师帮助检索。
2 结果与分析
2.1 抗、感品系棉铃虫围食膜蛋白含量
由表1可以看出,敏感品系棉铃虫的围食膜蛋白含量为22.19%,抗性品系棉铃虫的围食膜蛋白含量为26.99%,抗性品系蛋白含量显著高于敏感品系(P<0.05)。

表1 棉铃虫围食膜蛋白质含量1)
2.2 棉铃虫围食膜蛋白的NuPAGE电泳分离
NuPAGE电泳分离结果(图1)显示,重复3次的结果可以看出,分离效果清晰,可以分离出大约30种围食膜蛋白,且所有蛋白包括大分子量220 kD和小分子量10 kD都能分离出来。
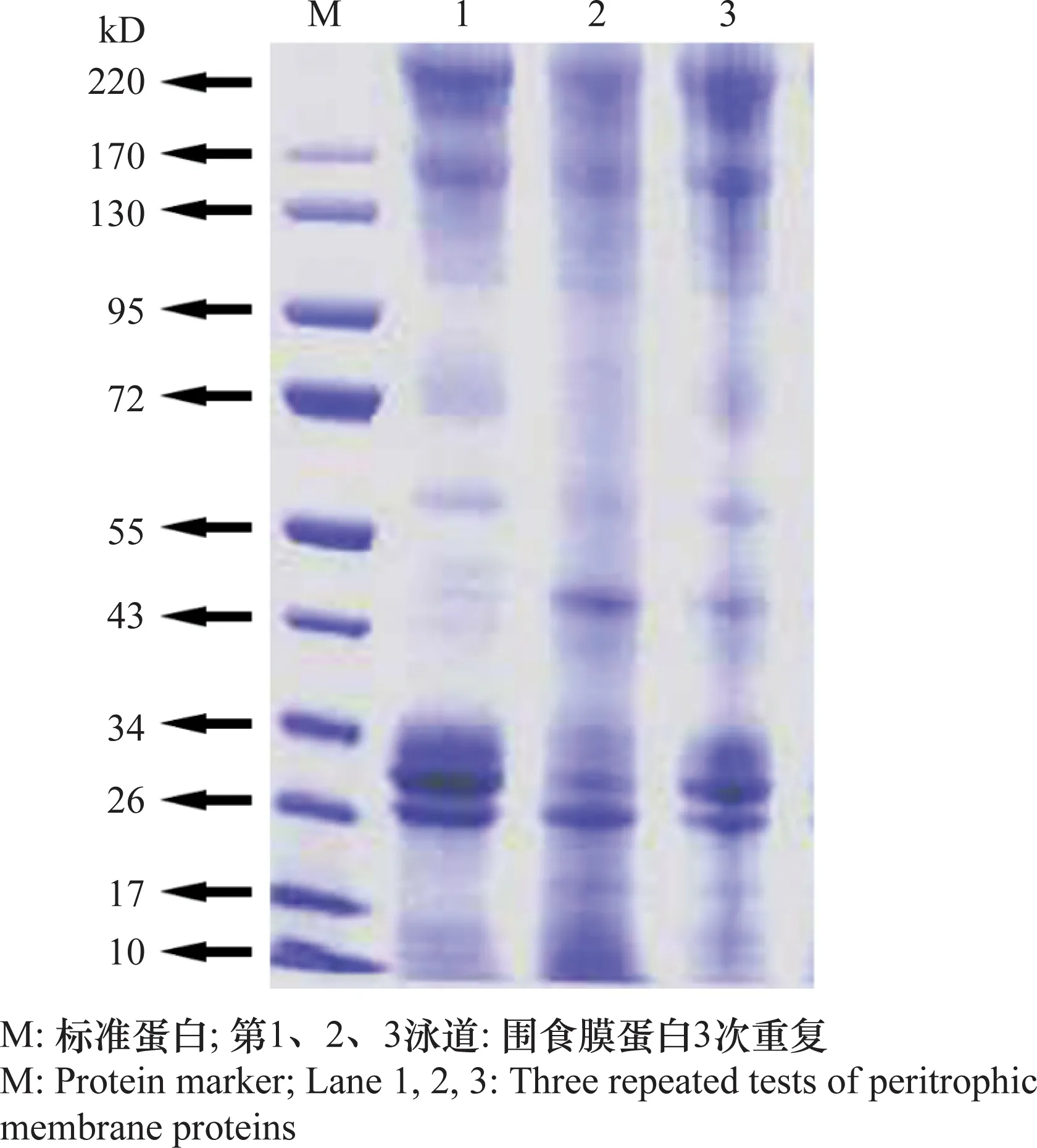
图1 棉铃虫围食膜蛋白质NuPAGE电泳结果
2.3 抗、感品系围食膜蛋白与Cry1Ac的结合能力比较
运用ligand blot检测分离纯化的抗、感品系棉铃虫围食膜蛋白与Cry1Ac的结合能力,结果发现蛋白大小分别为220、170、120、65、43、35 kD的6个围食膜蛋白可与Cry1Ac结合,而且不同品系中结合能力差异明显(图2)。由ligand blot结果可知,Cry1Ac与敏感品系的围食膜蛋白结合较强,与抗性品系结合弱。说明围食膜蛋白上可能存在Cry1Ac的受体。
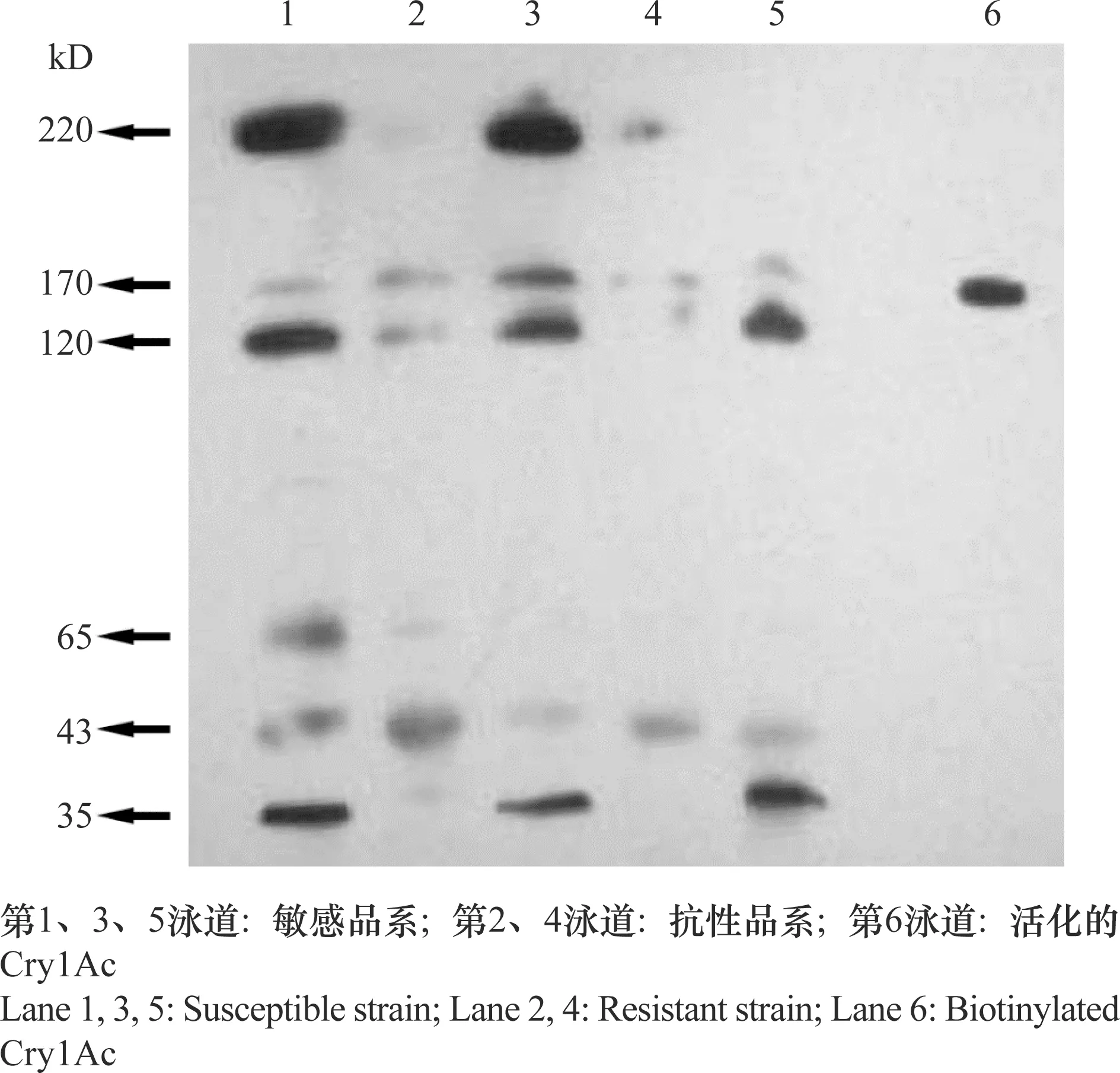
图2 Ligand blot检测CrylAc结合蛋白
2.4 棉铃虫抗、感品系围食膜差异蛋白分析及鉴定
运用NuPAGE Novex Bis-Tris Mini Gels电泳比较Bt抗、感品系棉铃虫围食膜蛋白,敏感品系(S)得到了220、170、120、60 kD左右的4个差异明显的蛋白条带,而在抗性品系(R)上很少或者几乎没有(图3)。

图3 棉铃虫抗感品系围食膜蛋白NuPAGE电泳分析图
2.5 棉铃虫抗、感品系围食膜差异蛋白的质谱鉴定
用MALDI-TOF质谱分析鉴定了2.4中筛选到的4个蛋白,获得肽指纹图谱PMF,通过比对成功鉴定得到了4个相关蛋白并获得相关生物学信息(表2)。
经NCBI blastn比对,蛋白1虽然有一定数量匹配结果,但是检索覆盖率都比较低,认为属于新蛋白。蛋白4与数据库中序列的一致性较低,也属于新蛋白。蛋白2与棉铃虫羧酸酯酶(carboxyl/choline esterase,61.7 kD)有较好的一致性。蛋白3与豌豆蚜Acyrthosiphonpisum的血影蛋白(spectrin beta chain-like,266.2 kD)有较好一致性(表2)。

表2 4种棉铃虫围食膜蛋白BLAST比对结果
3 结论与讨论
关于棉铃虫围食膜蛋白质分离与鉴定的研究比较多。张严峻等[18]采用SDS-PAGE不连续胶(3%分离胶,10%浓缩胶)发现很多不同分子量大小的蛋白质,分离出比较清晰的16个条带,蛋白质分子量一般在97.4 kD以下,很多蛋白质的分子量很接近。张小霞[22]研究发现使用12%分离胶和5%浓缩胶可以分离的围食膜蛋白质种类较多,至少有14种蛋白质,并且小分子量的蛋白质含量较高,分子量一般在97 kD以下,蛋白质的分子量也比较接近。Campbell等[15]以棉铃虫、美洲棉铃虫Helicoverpazea和烟芽夜蛾Helicoverpavirescens为试虫,发现利用无水三氟甲烷磺酸(TMFS)可以溶解包括几丁质在内的全部围食膜组分,可将蛋白从几丁质中解离,获得几乎所有的围食膜蛋白,并以此鉴定出了41种蛋白。梁振普等[17]参考Campbell等[15]的研究,利用TMFS与LC-MS/MS技术鉴定出了169个棉铃虫围食膜蛋白质。其中也包括了ALP、APN、ATP合成酶β亚基、ABC转运蛋白等可能为Bt杀虫蛋白受体的蛋白质,这也说明了棉铃虫围食膜蛋白确实与Bt抗性产生有一定的关系[23-26]。本研究运用Campbell等[15]和李怡萍等[21]的方法,将围食膜样品冷冻干燥后用TMFS法处理,然后进行NuPAGE梯度胶电泳,结果分离到约30种围食膜蛋白,分离效果较好。没有达到Campbell报道的41种蛋白,推测可能是因为选用的棉铃虫品系不同,存在地理差异,从而导致了分离出的蛋白数目不同。
有报道表明Bt Cry活化蛋白与中肠上皮细胞受体的结合,是Bt杀虫蛋白发挥作用的主要过程。类钙黏蛋白(cadherin-like protein,CAD)、氨肽酶N(aminopeptidase N,APN)、碱性磷酸酶(alkaline phosphatase,ALP)和ABC转运蛋白(ATP binding cassette sub family C)等都是重要的Bt杀虫蛋白受体[27-28]。其中APN为昆虫中肠BBMV中含量最高的蛋白酶(约占55%),在中肠的功能行使过程中发挥着重要作用[29]。
已有研究表明抗、感品系中肠上受体蛋白的差异表达可能是昆虫产生抗性的直接原因[6,11,14]。本研究使用2D Quant Kit测定总蛋白浓度,发现抗性品系蛋白含量显著高于敏感品系,其中敏感品系围食膜中蛋白含量为22.19%,抗性品系围食膜中蛋白含量为26.99%,推测围食膜中蛋白含量增加很可能和抗性的产生相关。其次,本研究测定的敏感品系棉铃虫围食膜蛋白含量和张严峻等[18]测定的34.47%的结果差距较大,推测可能是由于本研究使用的TMFS处理方法与张严峻等[18]使用的酸水解、碱水解以及Folin-酚处理方法不同,使得中间过程中损耗了蛋白质。
为了进一步验证围食膜上蛋白质的功能,本研究运用配体杂交技术比较了抗、感品系棉铃虫围食膜蛋白与Cry1Ac的结合特性。结果发现6个围食膜蛋白(大小为220、170、120、65、43、35 kD左右)可与Cry1Ac结合,且敏感品系上结合强度明显大于抗性品系。其中120 kD大小的蛋白推测可能是APN蛋白,65 kD大小的蛋白推测可能是ALP蛋白,大小与已有报道的棉铃虫中肠上的APN、ALP等Bt杀虫蛋白受体大小也基本一致[30-33]。Zhang等[34]发现APN基因突变导致棉铃虫对Cry1Ac抗性增加,Agrawal等[35]也发现减少斜纹夜蛾Spodopteralitura体内APN基因的表达,也会显著降低虫体对Cry1C的敏感性。也有报道表明,在部分鳞翅目抗性品系昆虫的中肠组织中,ALP的表达量有所下降[36-37]。
本研究利用NuPAGE电泳技术比较了抗、感品系围食膜蛋白的差异,获得了4种差异明显的蛋白,这4种蛋白只存在于敏感品系,在抗性品系中缺失;通过质谱鉴定和生物信息学分析方法,确定了其中2种为棉铃虫羧酸酯酶和血影蛋白,推测另外2种蛋白可能是新蛋白。羧酸酯酶是能水解含羧基官能团的酸酯、酰胺和硫酯类化合物的一类酯酶[38-39]。大部分羧酸酯酶与化学信息素代谢和杀虫剂抗性产生有关,如拟除虫菊酯类、氨基甲酸酯类、有机磷类等杀虫剂在昆虫体内的解毒过程中羧酸酯酶均发挥着重要作用[5]。也有文献报道棉铃虫中肠中有着较高活性的羧酸酯酶,在棉铃虫解除毒素毒性的过程中起着重要的作用[15]。血影蛋白(spectrin)是红细胞细胞骨架中最重要的蛋白质,它与锚蛋白(ankyrin)和跨膜阴离子交换蛋白(anion exchanger1, AE1)共同维持红细胞膜结构的稳定性和柔韧性[40-41]。而围食膜也是由中肠细胞分泌的一层非细胞组织,其中围食膜蛋白也有维持围食膜弹性和伸缩特性的功能,维持着围食膜的结构。综合以上研究结果,推测本研究中鉴定出的这两种蛋白很可能与Bt抗性有关。
本研究证明棉铃虫围食膜蛋白质中,存在能与Bt杀虫蛋白结合的蛋白,可能与Bt抗性相关,研究结果对于进一步明确Bt抗性机制、制定合理的Bt抗性治理策略提供了理论依据。
猜你喜欢
江苏农业科学(2016年11期)2017-03-21
哈尔滨医药(2016年3期)2016-12-01
新疆农垦科技(2016年2期)2016-08-21
西北农业学报(2016年6期)2016-07-02
现代农业(2016年5期)2016-02-28
现代农业(2016年4期)2016-02-28
现代农业(2015年5期)2015-02-28
癌变·畸变·突变(2015年4期)2015-02-27
现代检验医学杂志(2015年1期)2015-02-06
中国烟草学报(2012年5期)2012-04-12
