
寒温带地区典型森林类型中外生菌根真菌多样性和群落组成及其影响因素
2023-02-03 04:12孟兆云杨勋爵吕桂芬牛艳芳张亚萍
生态学报 2023年1期
孟兆云,李 敏,*,杨勋爵,吕桂芬,牛艳芳,张亚萍
1 内蒙古师范大学生命科学与技术学院,呼和浩特 010022 2 内蒙古自治区高等学校生物多样性保护与可持续利用重点实验室,呼和浩特 010022
外生菌根(ECM)真菌常与林木互惠共生,促进宿主植物对氮、磷、钾等多种营养元素的吸收,并增强植物的抗病、抗寒、抗盐及抗重金属能力以及对极端环境的适应能力[1—4];同时宿主植物提供给ECM真菌光合产物等物质以供其生长[5]。由于ECM 真菌的这些关键功能,森林树木在自然条件下很大程度上依赖于ECM真菌类群。因此,ECM真菌被认为是温带森林生态系统的关键组成部分。在长期的生态系统演化过程中,植物与真菌之间相互选择与适应,导致了ECM真菌与宿主植物之间存在一定的专一性。但ECM真菌与植物的共生关系可以超越单一的树-真菌伙伴关系,形成共同的菌根网络[6—7]。同一宿主植物可与多种不同的ECM真菌共生,同一ECM真菌也可与多种植物共生。由于菌根网络同时连接了多个不同个体的宿主植物和ECM真菌,因此对提高生态系统的生产力和增加资源利用互补性有重要作用[8—9]。ECM真菌与宿主植物的相互作用在整个陆生植物进化过程中扮演了重要角色,强烈影响着森林生态系统的功能[3,9—11]。ECM真菌的物种及功能多样性是维持生态系统稳定性的重要因素[12—13]。
由于ECM真菌需从宿主植物获得赖以生存的有机碳源,因此宿主植物被认为是影响ECM真菌群落组成的最重要的决定因素[14—17]。而对欧洲山毛榉(Fagussylvatica)的研究发现,非宿主植物的灌木层和草本层对ECM真菌的分布有显著影响[18]。也有一些研究表明,宿主植物对ECM真菌群落没有影响或影响很小[19—20]。同样,也有研究者报道,包括土壤和气候变量在内的非生物环境因素是ECM真菌群落的强大驱动力。例如,气候差异和土壤养分解释了日本两座山区[20]和美国明尼苏达州东南部的七种柳树[21]中ECM真菌群落的差异。Ge等认为美国山核桃的ECM真菌群落受土壤中的K、Ca、Mg、Mn、P、Zn等矿物质的影响较大[22]。Koizumi等认为温度是构建ECM真菌群落的重要决定因素[23]。在气候变化的背景下,ECM真菌群落多样性会发生相应的变化,譬如全球变暖和长期氮沉降会导致ECM真菌群落多样性的降低[24—25]。Gong等认为,ECM真菌群落在不同海拔区域和季节间存在强烈的物种替代和显著的组成差异,ECM真菌多样性在森林区域随海拔升高而降低[26]。上述研究表明,在不同的地理尺度上,ECM真菌多样性变化的关键驱动因子有所不同。因此,明确ECM真菌的物种多样性及其群落构建的影响因素对指导森林生态系统的健康可持续发展以及预测环境变化对生态系统的潜在影响具有重要意义。
白桦(Betulaplatyphylla)、樟子松(Pinussylvestrisvar.mongolica)、兴安落叶松(Larixgmelinii)和偃松(Pinuspumila)均耐寒,是我国寒温带地区的常见树种,对森林和人类的生物多样性做出了重大的生态和经济贡献。它们都是典型的ECM依赖型树种,而对该地区的ECM真菌多样性及群落组成的认识非常有限。因此,本研究采用高通量测序技术,分析了寒温带地区4种典型森林类型的ECM真菌多样性,同时通过分析土壤理化性质对ECM真菌群落组成的影响,确定了导致ECM真菌群落变化的主要驱动因素。本研究的目标是:(1)明确寒温带地区4种典型森林类型的ECM真菌多样性和群落组成;(2)明确ECM真菌多样性和群落组成的主要驱动因素;(3)明确植物-ECM真菌相互作用中宿主和ECM真菌的偏好。本研究假设:(1)土壤因子在构建ECM真菌群落中发挥了重要作用;(2)ECM真菌和宿主植物间存在显著的偏好性。
1 材料与方法
1.1 研究区概况
研究区位于大兴安岭北麓根河市,年封冻期210天以上,年平均气温-5.3℃。白桦土样采自内蒙古大兴安岭森林生态系统国家野外观测研究站(50°54′12.81″E,121°29′46″N,海拔848 m);偃松土样采自静岭(51°2′35″E,121°17′43″N,海拔1034 m);樟子松土样采自好里堡东山(50°45′3.40″—50°45′3.84″E,121°29′01″—121°29′14.68″N,海拔685—795 m)阳坡;兴安落叶松土样采自好里堡东山阴坡。林下土壤均为淋溶土(棕壤土)。针叶林下层几乎没有维管植物,白桦林的下层植被包括稀疏的灌木、草本植物和蕨类植物。
1.2 采样方法
土样采集于2018年8月5—7日,采样方法同前期研究[27]。在4种不同森林类型内,各随机设置3个50 m×50 m的样地。每个样地内收集10棵健康树木的土样。采集土样后置于冰盒保存,尽快带回实验室用于土壤理化性质的检测及高通量测序。
1.3 土壤理化性质的测定
土壤pH值测定采用电位法(水土比2.5∶1);土壤含水量(WC)测定采用烘干法;土壤全氮(TN)和土壤有机质(SOM)含量测定分别采用重铬酸钾氧化法和半微量凯氏定氮法测定;土壤有效磷(AP)和速效钾(AK)含量测定分别采用钼锑抗比色法和乙酸铵浸提-火焰光度法;土壤铵态氮和硝态氮含量测定分别采用靛酚蓝比色法(2 mol/L 氯化钾(KCl) 浸提)和紫外分光光度法[28]。
1.4 土壤总DNA提取和高通量测序
使用PowerSoil® DNA Isolation Kit提取土壤微生物DNA,选用真菌ITS5F(5′-GGAAGTAAAAGTCG TAACAAGG-3′)和ITS1R(5′-GCTGCGTTCTTC-ATCGATGC-3′)为特异性引物,进行真菌 rDNA ITS 区段 PCR 扩增。扩增完成后,对PCR产物用2%琼脂糖凝胶电泳检测,并采用Axygen凝胶回收试剂盒回收PCR产物。2%琼脂糖凝胶电泳检测合格后,采用 Illumina Miseq平台上进行2×300 bp的双端测序。文库构建、检测、上机测序等委托上海派森诺生物科技股份有限公司完成。测序完成后,筛选有效序列,去除Reads接头和barcode序列,并用FLASH软件对原始数据进行过滤,采用Usearch(version 7.0 http://drive5.com/uparse/)去除嵌合体序列,并对有效序列进行数据统计。
1.5 数据分析
使用QIIME软件,调用UCLUST序列比对工具,对获得的序列按97%的序列相似度进行归并和可操作分类单元(operational taxonomic unit,OTU)的划分,并选取每个OTU中丰度最高的序列作为该OTU的代表序列,采用BLAST工具在 UNITE 数据库(Release 5.0,https://unite.ut.ee/)中对ECM真菌OTUs进行物种注释,分别在不同分类水平上统计各样本的群落组成和丰度。所有的ECM真菌OTUs需经均一化处理。通过Galaxy在线分析平台(http://huttenhower.sph.harvard.edu/galaxy/),对属水平的相对丰度矩阵进行LEfSe分析。使用R软件进行ECM真菌属水平的群落组成的主成分分析(PCA);ECM真菌群落Alpha多样性指数的分析使用QIIME软件完成;采用SPSS 22.0软件完成ECM真菌丰度及土壤理化性质的Pearson相关性分析及显著性检验。利用Canoco(version 5.0)软件对土壤理化性质与ECM真菌群落组成进行冗余分析(db-RDA)。
2 结果与分析
2.1 不同森林类型中ECM真菌多样性和群落组成
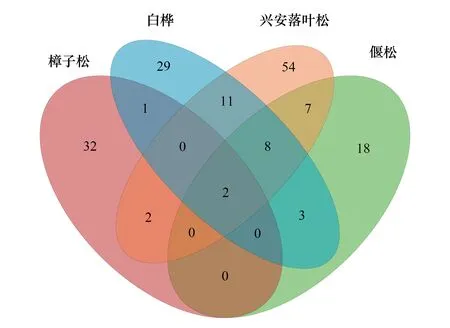
图1 不同森林类型ECM真菌OTUs数量Fig.1 The OTUs number in different forest types白桦:Betula platyphylla,Bp;樟子松:Pinus sylvestris,Ps;兴安落叶松:Larix gmelinii,Lg;偃松:Pinus pumila,Pp
4种典型森林类型根围土壤中共获得167个ECM真菌OTUs(图1)。兴安落叶松的ECM真菌OTUs数最多,为84个;其次是白桦,为54个;偃松和樟子松最低,分别为 38个、37个。4个树种的特有ECM真菌OTUs数分别为54个、29个、18个和32个。兴安落叶松的特有ECM真菌OTUs数占所有ECM真菌OTUs数的比例最高,为32.34%;其次为樟子松和白桦,分别占19.16%和17.37%;偃松最低,为10.78%。4个树种的共有ECM真菌OTUs仅有2个,占所有ECM真菌OTUs的1.20%,隶属于丝膜菌属(Cortinarius)和红菇属(Russula)。以上结果说明,不同森林类型的ECM真菌多样性有显著差异,树种对ECM真菌群落组成有显著影响。
所有ECM真菌隶属于2个门、4个纲、10个目、24个科、36个属。在门水平上,担子菌门(Basidiomycota)真菌的相对丰度占96.4%,子囊菌门(Ascomycota)真菌的相对丰度占3.6%。在纲水平上,蘑菇纲(Agaricomycetes)真菌的相对丰度最高,为96.4%,其次分别为座囊菌纲(Dothideomycetes,1.4%)、锤舌菌纲(Leotiomycetes,1.2%)和盘菌纲(Pezizomycetes,1.0%)真菌。在目水平上,红菇目(Russulales,30.9%)、蘑菇目(Agaricales,28.8%)、蜡壳耳目(Sebacinales,12.6%)、阿太菌目(Atheliales,9.9%)和革菌目(Thelephorales,8.6%)真菌的相对丰度均>5.0%。相对丰度>5.0%的科分别为红菇科(Russulaceae,30.9%)、丝膜菌科(Cortinariaceae,17.1%)、阿太菌科(Atheliaceae,9.9%)、革菌科(Thelephoraceae,8.6%)、蜡壳耳科(Sebacinaceae,8.0%)和丝盖伞科( Inocybaceae,7.2%)。红菇属(30.2%)、丝膜菌属(17.1%)、Piloderma(9.6%)、棉革菌属(Tomentella,7.2%)、丝盖伞属(Inocybe,7.2%)、蜡壳耳属(Sebacina,6.2%)的相对丰度均>5.0%,是该地区的优势ECM真菌类群(图2)。
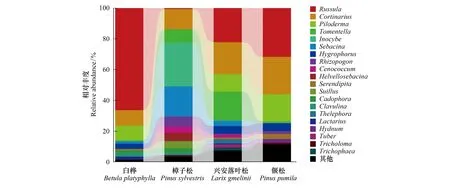
图2 不同森林类型ECM真菌属水平的群落组成Fig.2 The ectomycorrhizal fungal community composition on generic level in different forest types
2.2 不同森林类型ECM真菌群落组成差异
对4个不同森林类型ECM真菌优势属的分析表明(表1,图2),红菇属的平均相对丰度最高(30.2%),但在不同森林类型间的差异显著,白桦根围土壤中该属的相对丰度最高(66.4%),其次是偃松(31.7%)和兴安落叶松(22.2%);而樟子松根围土壤中该属真菌的相对丰度最低(0.6%)。丝膜菌属真菌的相对丰度在不同森林类型间无显著差异,分布较均匀。丝盖伞属在樟子松根围(28.4%)的相对丰度显著高于其它森林类型扩号中的数字表示其占所有ECM真菌的百分比;优势属后所标字母代表对样地内所有优势属的相对丰度的差异显著性检验,而各样地优势属后所标字母代表对该属在不同样地间的相对丰度的差异显著性检验;Bp:白桦,Betulaplatyphylla;Ps:樟子松,Pinussylvestris;Lg:兴安落叶松,Larixgmelinii;Pp:偃松,Pinuspumila(0.0%—0.3%)。白桦(9.5%)、兴安落叶松(11.3%)和偃松(17.6%)根围的Piloderma的相对丰度均较高,而樟子松根围没有发现该属真菌的OTUs。蜡壳耳属真菌在樟子松根围的相对丰度(19.6%)显著高于白桦(1.5%)和兴安落叶松(3.4%),而偃松根围并未发现该属真菌的OTUs。兴安落叶松(18.8%)根围土壤中棉革菌属的相对丰度显著高于樟子松(8.6%),白桦(0.6%)和偃松(0.9%)根围土壤中该属真菌的相对丰度显著低于上述二者。
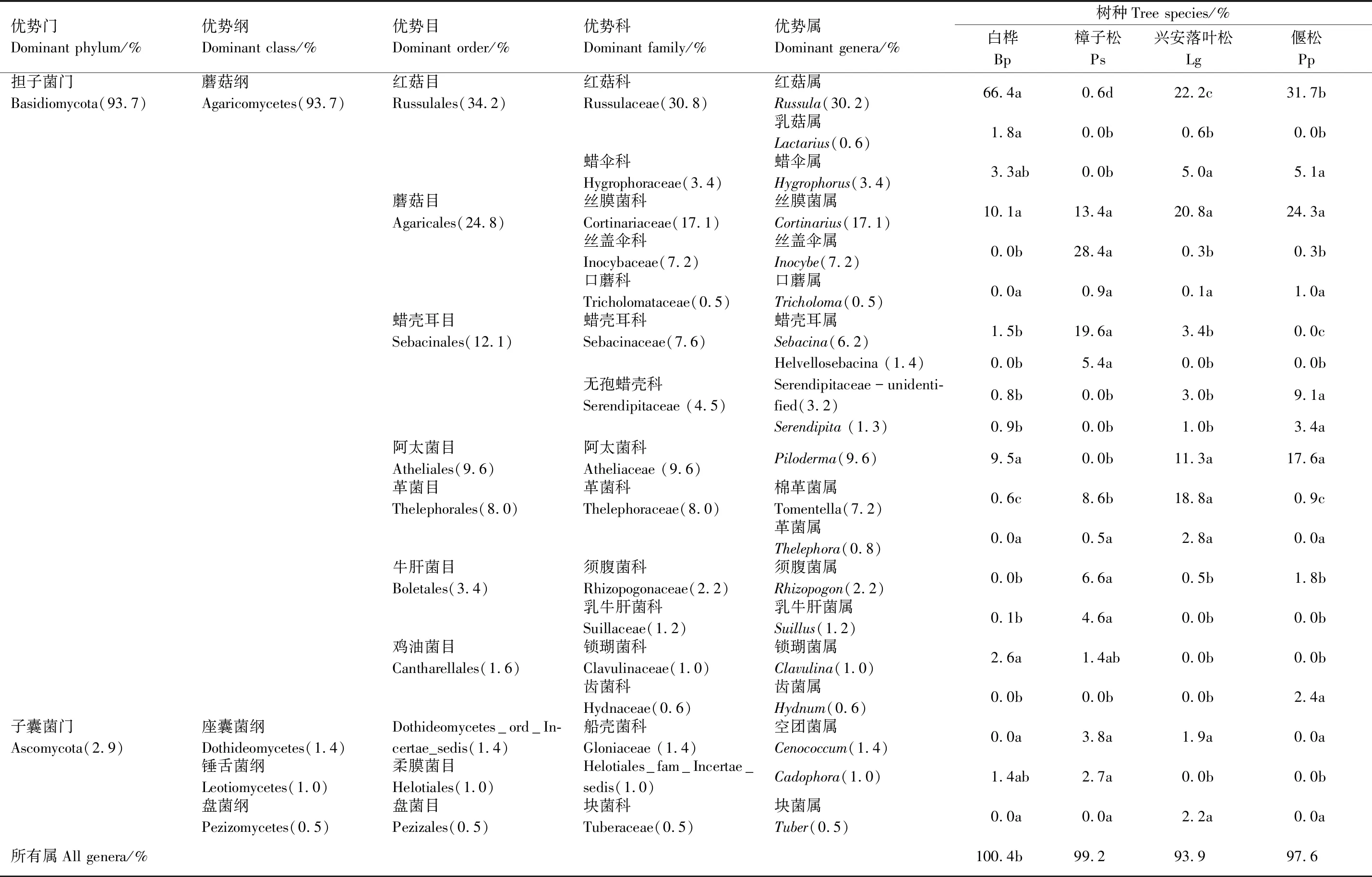
表1 不同森林类型外生菌根(ECM)真菌群落(相对丰度前20个属)组成Table 1 Composition of Ectomycorrhizal fungal communities (top 20 genera in relative abundance) in different forest types
还有部分ECM真菌属是某种森林类型的特有属,如Helvellosebacina属真菌的OTUs只出现在樟子松根围,齿菌属(Hydnum)真菌的OTUs只出现在偃松根围,块菌属(Tuber)真菌的OTUs只出现在兴安落叶松根围。对4个不同森林类型ECM真菌的LEfSe物种差异分析(图3)表明,白桦根围的红菇属、兴安落叶松的棉革菌属、偃松的Serendipita和樟子松的丝盖伞属、蜡壳耳属和须腹菌属(Rhizopogon)均是差异指示物种。以上结果表明,不同树种的优势ECM真菌在属水平上的组成存在显著差异。
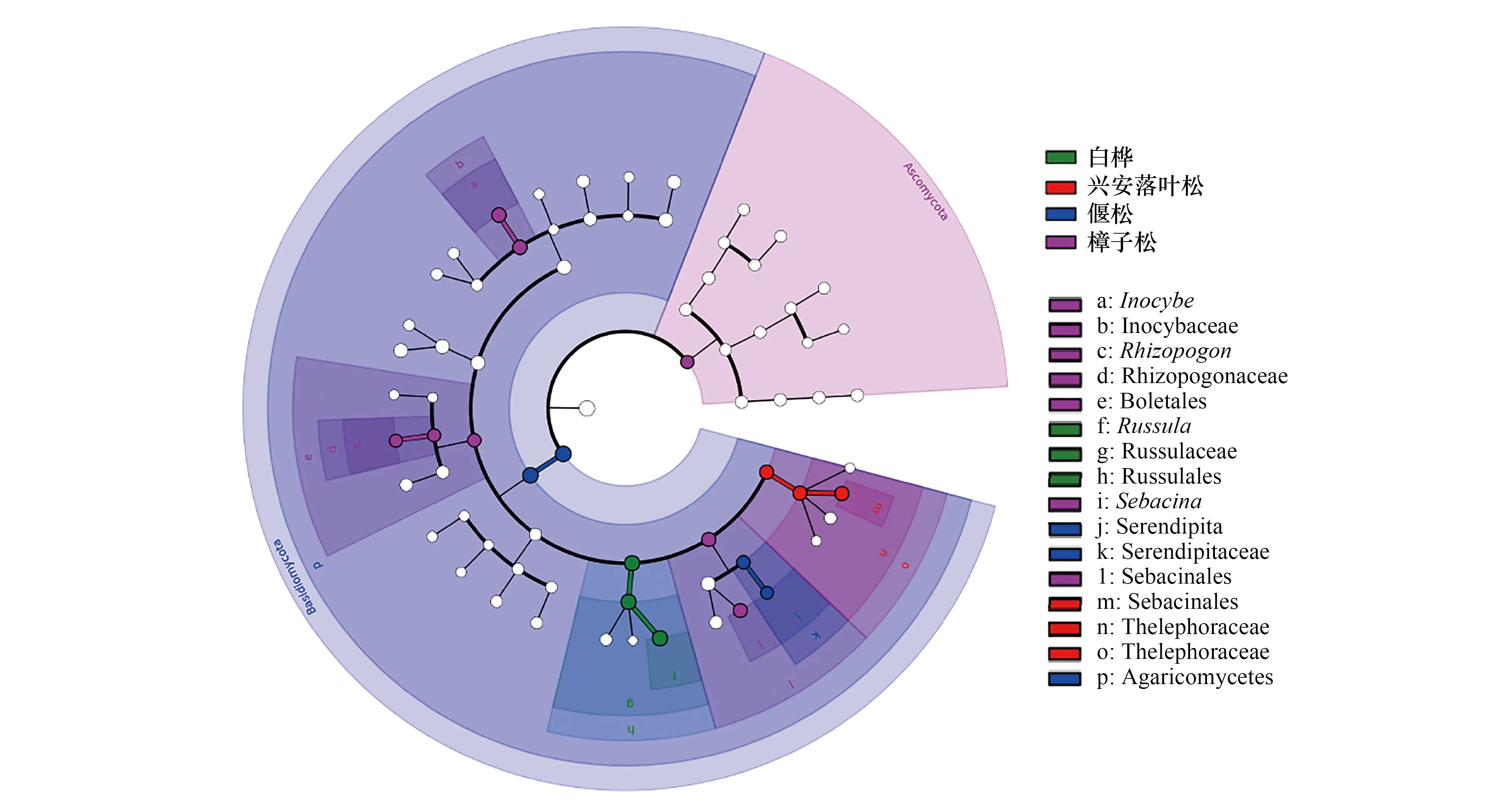
图3 不同森林类型ECM真菌LEfSe物种差异分析 Fig.3 LEfSe species difference analysis in different forest types
2.3 土壤理化因子对ECM真菌群落组成的可能影响机制
土壤理化因子对ECM真菌群落有显著影响(表2,图4)。所有因子共解释了ECM真菌群落变异的92.4%,其中硝态氮的解释度最高(60.4%),其次分别为土壤总氮(10.6%)和pH(9.5%),这3个因子对ECM真菌群落组成均有极显著影响(表2)。
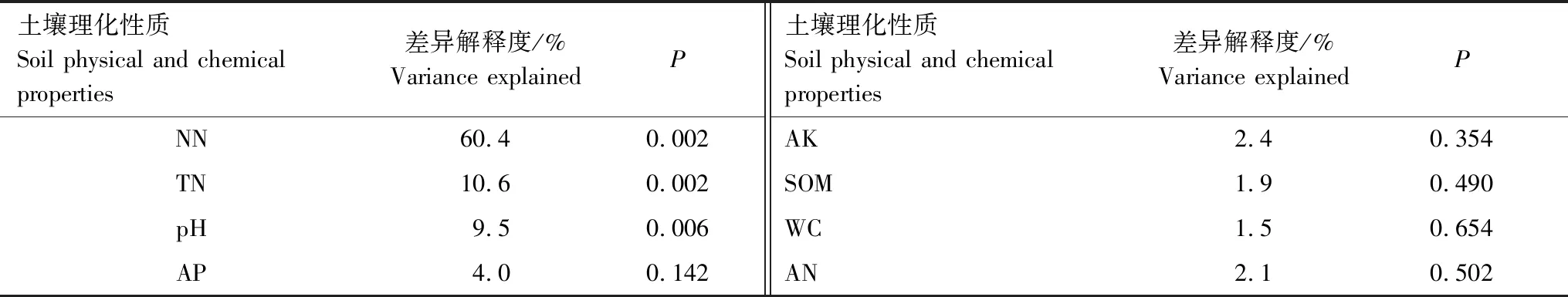
表2 土壤理化因子对ECM真菌群落组成的冗余分析Table 2 Results for db-redundancy analysis testing effects of soil physical and chemical factors on the ECM fungal community composition
冗余分析表明(图4),ECM真菌受土壤理化性质的显著影响,如口蘑属(Tricholoma)、空团菌属(Cenococcum)、须腹菌属、蜡壳耳属、丝盖伞属、乳牛肝菌属(Suillus)等更适宜栖息在干旱、高pH的森林土壤中;Serendipita、齿菌属、蜡伞属(Hygrophorus)、Piloderma、革菌属和棉革菌属在相对肥沃(高有机质、高氮)的森林土壤中的相对丰度更高;而丝膜菌属、块菌属和锁瑚菌属(Clavulina)对环境的依赖性并不明显。
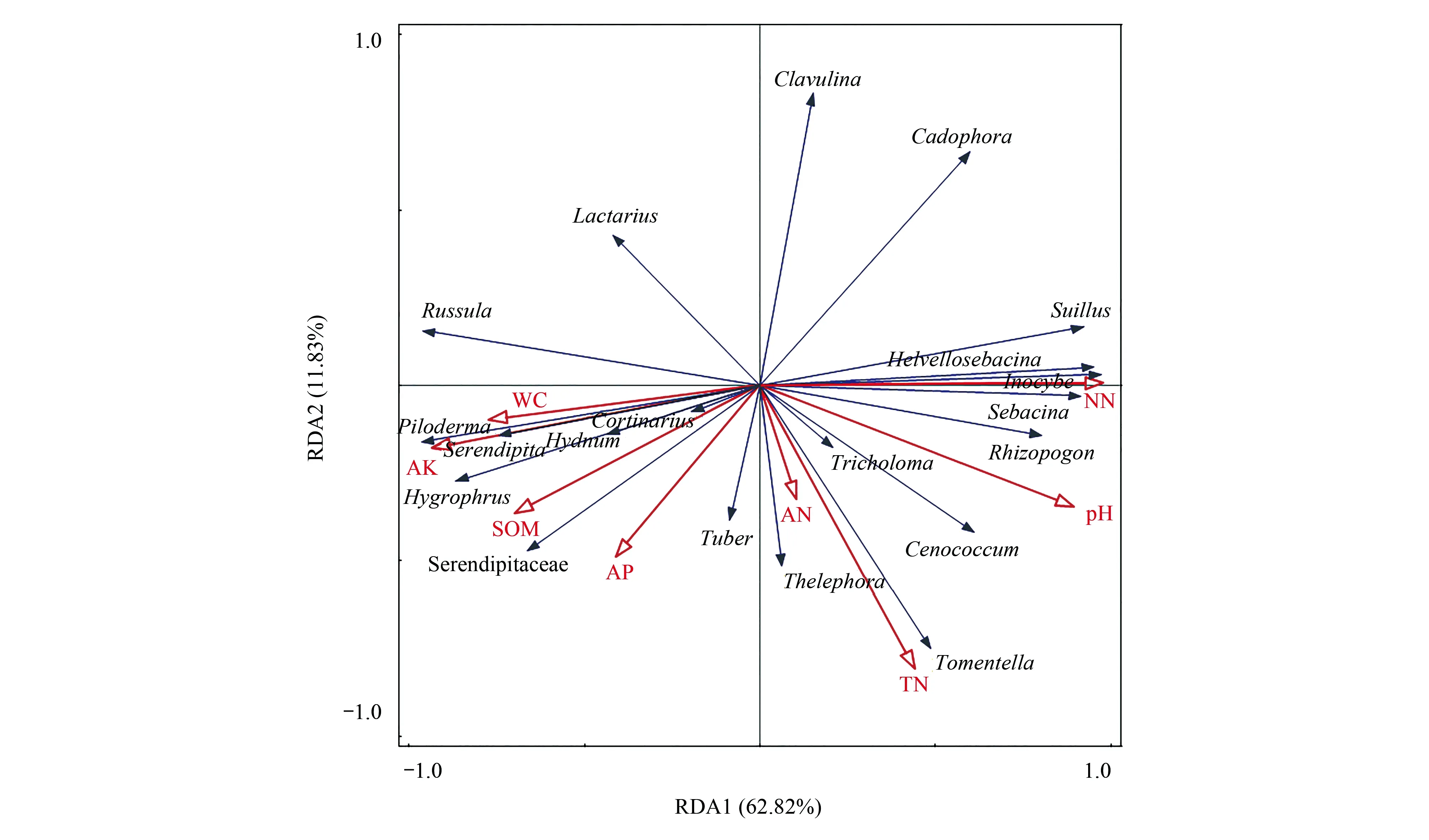
图4 ECM真菌群落组成(属水平)与土壤理化因子的冗余分析(RDA)Fig.4 Redundancy analysis (RDA) of ECM fungi community composition (generic level) and soil factorsNN:硝态氮Nitrate nitrogen;TN:土壤全氮total nitrogen;AP:土壤有效磷Available phosphorus;AK:速效钾Available potassium;SOM:土壤有机质soil organic matter;WC:含水量Water content;AN:铵态氮Ammonium nitrogen
3 讨论
3.1 不同森林类型ECM真菌多样性及群落组成
菌根真菌是植物共生微生物的主要组成部分,白桦、樟子松、兴安落叶松和偃松是大兴安岭寒温带地区的常见树种,是典型的ECM依赖型树种。ECM真菌与植物的共生对提高生态系统的生产力和增加资源利用互补性有重要作用[7—8,10,12]。本研究结果表明,不同森林类型土壤中均有较高的ECM真菌多样性,其中担子菌门真菌的相对丰度占96.4%,子囊菌门真菌占3.6%。此结果与大多数在森林生态系统中的研究结果一致,形成ECM的真菌多数为担子菌门真菌,少数为子囊菌门真菌[29]。由于森林土壤中存在较厚的凋落物层,担子菌门真菌在土壤中的优势地位可能与它们降解复杂木质纤维素组分的能力有关[30]。
寒温带地区4种典型森林类型的特有ECM真菌OTUs数占所有ECM真菌OTUs数的10.78%—32.34%;而共有ECM真菌OTUs仅有2个,占所有ECM真菌OTUs的1.20%;另外不同森林类型的优势ECM真菌在属水平上的组成存在显著差异;说明ECM真菌与树种间存在选择偏好性,ECM真菌群落组成受宿主植物的显著影响,与本研究提出的假设一致。
研究表明,菌根真菌更直接地依赖于树木凋落物,由于不同森林类型凋落物的化学成分不同[31],引起凋落物的分解速率不同,由此导致不同林分土壤有机碳等含量不同[32—33]。凋落物性质的不同为ECM真菌提供了不同的生态位和可利用资源[34],因此ECM真菌群落组成受森林植物群落的显著影响[35]。另外,由于针叶林下层几乎没有维管植物凋落物,因此针叶林凋落物分解速率相对阔叶林显著降低[32];同时针叶林凋落物层一般具有较高的碳氮比,可能会造成某些ECM真菌的不利生长条件[36]。相比之下,较开阔的白桦林的下层植被包括稀疏的矮灌木、草本植物和蕨类植物。因此在白桦林下土壤中观察到的ECM真菌群落相对于樟子松林和偃松林更丰富。
本研究中,除丝膜菌属真菌的相对丰度在不同森林类型的土壤中分布较均匀外,其余相对丰度>5.0%的优势菌如红菇属、Piloderma、棉革菌属、丝盖伞属和蜡壳耳属真菌在不同森林类型土壤中的丰度差异显著,可能是由于不同真菌的定植策略和资源需求各不相同,另外可能与植物在不同生长阶段对不同功能性真菌存在不同程度的偏好性有关[37]。丝膜菌属中的几种物种已被证明可以产生锰-过氧化物酶结合物,从而能够从复杂的有机物中分解获得氮[38]。本研究中,具有凋落物分解活性的丝膜菌属的丰富度较高,且在樟子松、兴安落叶松和偃松的相对丰度比白桦高,这与丝膜菌属真菌通常在较老的针叶林中占主导地位的研究结果一致[39]。本研究表明,红菇属真菌在寒温带针、阔叶林中的丰度都很高,与Danielsen等[40]和Mundra等[37]的研究结果一致。红菇属是北温带森林的成熟林中最为常见的ECM真菌类群之一,可能由于红菇属真菌具有更好的养分获取策略及高效的能分解获得氮和磷的酶系统而具有竞争优势[39]。综上,不同森林类型对ECM真菌群落具有较强的选择性,而这些ECM真菌群落形成的菌根共生网络又为宿主植物提供所需的营养和水分,双方互惠共生,共同维持森林生态系统的健康发展。
3.2 土壤因子在构建ECM真菌群落中的贡献
ECM真菌群落组成除受树种的影响外,还受到土壤理化性质的显著影响。本研究中不同森林类型土壤理化性质均存在显著差异,是由于植物凋落物和根系中含有大量的有机物,经微生物的分解后,改变了土壤理化性质,从而显著影响了ECM真菌群落组成。硝态氮、土壤总氮和pH对ECM真菌群落组成均有极显著影响,反映出ECM真菌群落组成受土壤因子的显著影响,与本研究提出的假设一致。硝态氮对ECM真菌群落变异的解释度最高,为60.4%;其次分别为土壤总氮(10.6%)和pH(9.5%)。土壤硝态氮含量与红菇属、Piloderma等ECM真菌的丰富度均呈显著负相关,是由于ECM真菌群落的生长吸收了土壤中的氮素,显著降低了根围土壤中氮的含量[41]。土壤硝态氮含量显著影响了土壤pH,也可能是造成ECM真菌群落组成差异的原因[42]。土壤pH与红菇属、Piloderma等ECM真菌的丰富度均呈显著负相关,这是由于ECM真菌群落在生长繁殖中产生了大量的有机酸,从而降低了土壤pH值[43];同时,土壤pH通过影响土壤基质的组成、化学性质从而显著改变了土壤理化性质,进而对ECM真菌群落组成产生显著影响。
本研究中,丝膜菌属真菌的相对丰度随土壤有机质含量的增加而增加,与Koizumi等的研究结果一致[23],与其分泌大量的过氧化物酶、从而能够有效分解有机物有关[38]。乳牛肝菌属真菌是典型的营养贫乏地区的优势菌[44],支持了本研究中乳牛肝菌属更广泛地分布在土壤有机质含量低的樟子松林下的结果。本研究中,Piloderma真菌在较低的土壤 pH值、低硝态氮环境中更丰富,与Peltoniemi等[45]、Högberg等[46]以及本课题组[47]前期的研究结果一致。研究表明,Piloderma真菌具有产生酸性磷酸酶的巨大潜力,能够释放有机磷并提供给宿主植物[48—49],暗示了Piloderma真菌可能在营养贫乏地区的森林生态系统的关键营养循环中发挥了重要作用。
温度是构建 ECM真菌群落的重要决定因素[24,44],表明ECM真菌物种的分布与其固有的温度生态位相对应。研究表明,ECM真菌群落可能是生态系统对温度变化恢复能力的关键,可能减轻全球变暖的影响[44,50]。本研究分析了寒温带森林生态系统中的ECM真菌多样性及群落组成,能够为建立关于ECM真菌群落如何响应全球生态系统环境变化的假设提供基础。
4 结论
本研究分析了寒温带4种典型森林类型ECM真菌多样性和群落组成,并分析了土壤因子在ECM真菌群落构建中的贡献。4种典型森林类型均具有较丰富的ECM真菌多样性,担子菌门真菌占绝对优势,红菇属、丝膜菌属、Piloderma、棉革菌属、丝盖伞属和蜡壳耳属真菌是该地区的优势ECM真菌类群。不同森林类型的优势ECM真菌组成存在显著差异。硝态氮、土壤总氮和土壤pH值对ECM真菌群落组成均有极显著影响,说明树种及土壤因子对ECM真菌群落组成均有显著影响。
猜你喜欢
当代水产(2022年8期)2022-09-20
昆明医科大学学报(2022年2期)2022-03-29
食品安全导刊(2021年20期)2021-08-30
科学(2020年2期)2020-08-24
河南科学(2020年3期)2020-06-02
农民致富之友(2019年8期)2019-05-22
小学科学(学生版)(2018年8期)2018-08-21
现代园艺(2018年2期)2018-03-15
现代园艺(2018年2期)2018-03-15
北极光(2018年12期)2018-03-07
