
微囊藻水华暴发的水动力机理与模拟研究进展
2023-01-31 06:53吴一红杨方宇
中国水利水电科学研究院学报 2022年5期
杨 宇,曾 利,吴一红,杨方宇,刘 丰,韩 瑞
(中国水利水电科学研究院 流域水循环模拟与调控国家重点实验室,北京 100038)
1 研究背景
微囊藻水华是全球普遍面临的水域生态问题[1],我国也是微囊藻水华频发的国家。例如2007年太湖蓝藻水污染事件,影响了无锡市的用水安全,引起国内外的高度关注[2];三峡水库蓄水以来,香溪河、小江等支流速度变缓,库湾多次暴发不同程度的微囊藻水华[3-4],如图1所示。微囊藻水华暴发会产生诸多危害:①降低水体溶解氧浓度:大量微囊藻聚集在水体表层,抑制大气中氧气溶于水体,并通过降低透光性,减少水体中植物光合作用产生的氧气,同时,死亡藻类分解又会大量消耗氧气,进而导致鱼类等水生物缺氧,甚至死亡[1];②产生毒素:微囊藻死亡分解会产生有毒物质,人类、家畜以及其他动物接触后可能会产生不良反应[5-6];③生态系统失衡:大量微囊藻聚集在水体表面,会造成食物链断裂,原有的群落关系遭到破坏,生态功能逐渐退化[7];④影响供水:水体中微囊藻浓度过高,会严重降低水厂水处理效率,大幅增加制水成本[8];⑤影响旅游业:微囊藻大量聚集,会产生刺鼻性异味,不利于旅游、娱乐、观赏等[9]。微囊藻水华的暴发过程十分复杂,涉及温度、营养盐、光照、水动力等多种因素[10-13]。温度、营养盐、光照主要影响微囊藻生长,水动力直接影响微囊藻的迁移聚集[14]。在富营养化水体中,藻类生物量大多充足,水动力在水华暴发中具有重要作用。因此,微囊藻水华暴发的水动力机理与模拟对水华防治具有重要意义[15],已成为环境与生态水力学研究的前沿热点与难点。本文旨在对微囊藻水华暴发的水动力机理与模拟研究工作进行总结与分析,包括梳理静水环境下微囊藻个体与群体的垂向迁移机制、典型流态下微囊藻聚集的水动力机理以及微囊藻迁移分布数值模拟研究进展,凝炼该领域需要解决的关键问题,以为水华防治研究提供参考。

图1 蓝藻水华
2 微囊藻迁移分布的水动力机理
2.1 微囊藻自主迁移的生物学机制微囊藻属于蓝藻门,是一种最原始、最古老的藻类植物,属于光能自养型原核生物[16]。环境水体中,微囊藻通常以个体和群体两种形式存在。微囊藻细胞单体内存在由多个圆柱形伪空胞(Gas Vesicle)叠加而成的气囊(Gas Vacuole)。伪空胞通常以可变六边形方式紧密排列,可大大减少伪空胞腔壁体积,增大了腔气体积[17]。伪空胞主要成分为蛋白质,其疏水性能保证空气进入的同时又能截止外界液体,其较强的刚性能使伪空胞不易变形,压强每增加0.1 MPa其体积仅缩小千分之一[18]。微囊藻细胞被多层结构所包被,主要成分是胞外多糖(extracellular polymetric substances)。胞外多糖被认为是形成微囊藻群体的重要物质基础,通常以两种形式存在,其一是荚膜(或称胶鞘),紧贴细胞壁,其二为黏液层,在荚膜外。根据微囊藻胞外多糖与细胞结合的紧密程度,可将其分为:①紧密结合型,紧密附着在细胞壁外;②松散结合型,可向周围扩展、无明显边缘的黏液层;③溶解型,分布在群体最外层,多以胶体状或溶解性分子形式存在,极易分散到水中[19]。微囊藻形成群体有两种方式:细胞分裂和细胞黏结[20]。细胞分裂是指分裂的细胞留在同一胶鞘内形成群体,细胞排列很整齐,倍增时间一般是2 d,多数为5~7 d,速度较慢;细胞粘结是指单细胞通过胞外多糖的黏性形成群体,速度较快。与单细胞相比,微囊藻群体在野外更具生存优势,主要体现在:群体比单细胞对光强的耐受力更强;在水体波动较大的环境中,群体具有独特的营养盐利用策略;群体抵御被浮游动物捕食的能力更强;群体内部形成的细胞间隙能提供较大的浮力,有利于克服水体扰动带来的裹挟力。
2.2 静水环境中微囊藻的垂向迁移机理研究表明,静水环境中微囊藻的自主迁移主要受个体或群体浮力影响[20-21]。关于微囊藻浮力调节机制,主要有四种解释:细胞膨胀压、伪空胞的合成与破裂、镇重物含量、群体细胞间隙。Reynolds等认为活细胞膨胀压为0.3~0.4 MPa,随着水深的增加,静水压力也逐渐增加(0.011 MPa/m),在较低外部压力下伪空胞就容易塌陷,导致细胞浮力减弱,细胞下沉[21]。然而,Walsby发现气囊周围的蛋白质株具有较高的膨胀强度,能使细胞承受湖泊中的压力,膨胀压不是导致微囊藻垂向迁移的主要原因[17]。Hayes和Walsby认为伪空胞能为微囊藻垂向迁移提供较大的浮力,但伪空胞的合成需要24 h以上[22],这种机制虽能说明伪空胞是微囊藻细胞上浮或悬浮的主要因素,但解释不了水华在数小时内生消的现象。Van Rijn和Shilo[23]通过设置光照范围与营养物质区域,发现水表面的藻细胞碳水化合物比例较低,10 m水深处藻细胞碳水化合物比例高,因此认为细胞内碳水化合物的波动是造成细胞迁移的主要原因;Ibelings等[24]认为细胞浮力的日变化是通过碳水化合物的含量改变的,而不是伪空胞含量的变化。Walsby等[25]在研究夏季分层湖泊中束丝藻的垂直迁移发现:细胞内镇重物(主要包括蛋白质、糖类、核酸、糖脂)含量白天增加夜晚减少。镇重物的密度大于水。白天藻细胞进行光合作用,增加了镇重物的含量,伪空胞提供的浮力不足以抵消镇重物的积累,细胞会下沉;当光强降低时,呼吸作用大于光合作用,消耗胞内糖类物质,细胞密度降低,从而引起细胞上浮[26],因此有 “昼沉夜浮”的说法。对于微囊藻单细胞而言,伪空胞破裂会使浮力迅速丧失,且伪空胞重新合成速度较慢,细胞增殖对伪空胞的稀释作用也需要以细胞的世代时间来度量,因此除伪空胞以外还应存在其它浮力影响因子。张永生等[27]发现微囊藻群体内具有大量的细胞间隙,这些细胞间隙能为微囊藻群体提供足够的浮力,使其上浮至表面,但目前无法准确检测细胞间隙的气体含量及密度,只能定性分析细胞间隙对上浮的影响。近来的实验观测表明,微囊藻群体在上浮过程中还具有自主扩散特性[28]。
2.3 动水环境中微囊藻迁移聚集的水动力机理蓝藻水华暴发具有时空多变性,有时从出现到消失只有几个小时,此外发生在午后的情形也时有出现。这与 “昼沉夜浮”生长机制显著不符,需寻求更合理的科学解释。研究表明,富营养化水体中微囊藻水华暴发与微囊藻迁移聚集的水动力机理密切相关。
国内外学者对动水环境中微囊藻群体的迁移分布开展了较多研究[29-34]。动水条件下微囊藻水华暴发不仅涉及群体的自主上浮聚集过程,还伴随水流携带与湍动扩散、群体破裂与聚集,机理非常复杂。对于微囊藻来说,通常情况下,伪空胞提供的浮力不足以使其大量上浮至水体表面,只有形成较大的群体进一步增大浮力,才能克服水流垂向挟带,上浮形成水华。实际上,野外观测到的微囊藻多数以群体形式存在。群体的形成与胞外多糖的分泌密切相关,因此研究水流对胞外多糖的分泌、藻类生长以及群体尺寸的影响有助于深入理解微囊藻水华暴发的水动力机制。当前,相关研究主要以室内实验为主[35-37]。室内实验主要利用振荡式摇床、造波机、转筒、格栅来形成水动力扰动,研究其对胞外多糖分泌、藻类生长及群体尺寸变化的影响。相关研究发现:特定的扰动(50和100 r/min)对微囊藻细胞具有刺激作用,诱导其产生更多的胞外多糖,提高细胞黏性,促进微囊藻小群体快速形成大群体[36],这与野外观察到的台风过后微囊藻群体迅速增大的现象一致[34];同时扰动方式也会影响胞外多糖合成,间歇性扰动能促进胞外多糖的分泌,有利于大群体形成与维持,但长时间持续扰动会抑制微囊藻生长,使群体逐渐变小[37]。转筒模型实验发现[38-39]:流速影响细胞生长和群体大小,湍动耗散影响细胞新陈代谢及营养物质吸收,剪切力影响细胞形态;当流速为35 cm/s时,铜绿微囊藻群体尺寸最大,流动增加了细胞间的接触机会,促进了生长滞后期菌落的形成,但在指数生长期,流动对微囊藻群体具有冲刷作用;O’Brien等[40]采用格栅搅拌槽实验研究了湍流混合强度(10-9~10-4m2/s3)对微囊藻群体大小的影响,结果表明:随着湍动耗散的增大,群体尺寸整体变小,不利于水华的形成。此外,Yang等[41]采用解析的方法,分析了浮力控制型藻细胞在明渠流中的迁移特性。
风生流、异重流是湖泊、水库中广泛存在的流态。已有研究主要聚焦于这两种流态下的水华暴发规律[42-47]。水动力对藻类的垂向与水平分布均有重要影响[48]。一般来说,强水动力导致水体中生物量沿垂向充分混合,表层水华不易形成;同时,较强的水动力混合也增加形成更大和更多藻群体的机会,一旦水动力条件变弱,蓝藻群体便会向上迁移聚集,暴发水华[30]。例如秦伯强等[33]通过野外监测数据并结合室内实验发现:台风过程中藻类分布主要受水动力影响,水体表层和近底层叶绿素浓度几乎相等;而台风过后,水动力作用减弱,微囊藻群体开始向水面迁移,且群体上浮速度随直径的增大而增大。研究表明,对于流场主要受风驱动的浅水水域,风向和风速对蓝藻水华暴发有重要影响,只有在临界风速以下微囊藻才能克服水流的携带而向水面聚集形成水华,但不同的研究中提出的临界风速不尽相同[29,49-53]。表1给出了不同文献中根据野外观测资料提出的蓝藻水华暴发临界风速值。高永[54]基于太湖蓝藻垂向分布的实测资料,分析了风速对不同尺寸蓝藻群体垂向分布的影响,结果表明:同一风速下,尺寸较大的群体垂向分布更集中,而尺寸较小群体分布更均匀,因此群体尺寸可能是导致临界风速不一致的原因。垂向水动力影响能上浮至水面上微囊藻的数量和聚集程度,而水平向水动力决定了微囊藻的漂移、堆积和斑块形成过程[55-59]。秦伯强等[33]提出了蓝藻水华暴发的概念性解释:在适宜的温度、光照、营养盐条件下,蓝藻细胞分裂增殖并分泌胞外多糖形成小群体;在适宜的风浪条件下小群体碰撞形成大群体,大群体具备更大的上浮速度;在临界风速以下大群体能克服水流的携带迅速向水面聚集。

表1 蓝藻水华暴发的临界风速
刘德富等[4]基于 “临界层理论”[60]研究了三峡支流水华生消机理。研究表明[3-4,45-46]:由于干支流的水温差,支流库湾存在明显的分层异重流[45-46],水流沿深度方向出现反向流动,改变了水体原有的水温分层、光学特性及营养盐分布特征,靠近河口的深水区分层较弱,远离河口的浅水区分层较强,水体混合层深度沿库湾向上游逐渐变小;同时支流流速较缓,泥沙沉降导致水体透明度增大,临界层深度变深。临界层是藻类接受光照进行有效光合作用的最大深度,它直接影响藻类的生长繁殖过程;混合层是藻类可以迁移悬浮的水深范围,水体掺混越剧烈藻类越容易被携带至真光层以下,增大藻类的死亡率[61-62]。当支流水体混合层深度小于临界层深度,藻类就能大量接受光照而繁殖,易于暴发水华;当混合层深度大于临界层深度,藻类呈负增长,不易暴发水华。
综上所述,在微囊藻迁移分布的水动力机理研究领域,尚需深入研究的工作包括:(1)微囊藻群体形态对其自主迁移速度的影响机理;(2)微观尺度上微囊藻群体聚集与破碎的水动力机理;(3)风生流、异重流、风浪等典型水动力环境中微囊藻群体迁移的特征规律;(4)高浓度微囊藻群体与水流的互馈机理。
3 微囊藻迁移分布的数值模拟
3.1 粒子模型粒子模型将水体中微囊藻当作标量物质或者活性物质进行模拟。对于垂向自主迁移能力较弱的微囊藻细胞而言,可将其当作标量物质,模拟其随流迁移过程;对于自主迁移能力较强的微囊藻群体来说,通常将其当作活性物质,垂向迁移速度不可忽略。在鞭毛藻个体迁移模拟研究领域,也存在相关研究[63-67]。与微囊藻的自主迁移不同,鞭毛藻主要通过摆动鞭毛实现自主游动。当前,微囊藻群体的活性粒子模型主要基于两个假设:一是忽略藻群体垂向迁移对水体的影响;二是忽略藻群体之间的相互作用。
静水环境中,微囊藻的垂向迁移速度受微囊藻密度变化影响,而光照是影响微囊藻密度变化的重要因素[68]。部分个体模型忽略光照对微囊藻群体密度的影响,采用恒定上浮速度[69],这与野外观测结果不一致[70]。鉴于光照的影响,越来越多模型考虑了光对藻密度的影响。例如:Kromkamp等[71]考虑了蓝藻密度随光辐射的变化规律,即强光下沉、弱光上浮;Visser等[72]通过室内实验获得了微囊藻密度与光辐射的关系,尤其是强光对微囊藻生长的抑制作用;Wallace等[73]建立了考虑碳水化合物累积效应的藻密度变化模型;Guven和Howard[74]考虑了氮、磷、光协同作用下的藻密度变化。表2给出了藻密度变化模型。

表2 藻密度变化模型

续表2 藻密度变化模型
粒子模型中,单日光强随时间的变化可采用如下公式计算[72]:

式中:Is为水体表面接受的光强;Im为水体表面接受的最大光强;DL为光周期。
光强沿水深方向的衰减采用Lambert-Beer定律[72]:

式中:Iz为水深z处的光强;K为水体的消光系数;z为群体所在的水深位置。
在模拟连续多日微囊藻浓度变化时,俞茜等[77]采用如下光强函数:
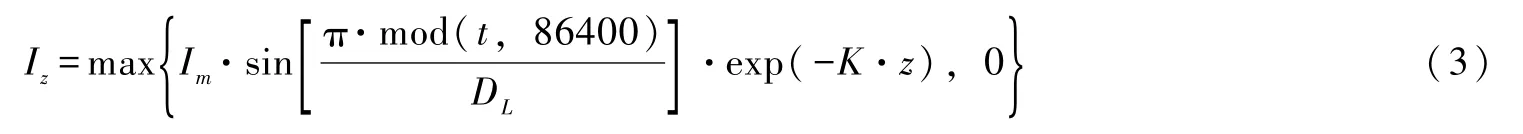
光强的变化会导致微囊藻细胞密度发生变化[72]:

式中各变量含义见表2。
考虑碳水化合物积累的延时效应,细胞密度随日内光照强度的变化而改变[73]:

式中:τr为响应时间,其余变量含义见表2。
由单细胞密度可以得到群体密度[78]:
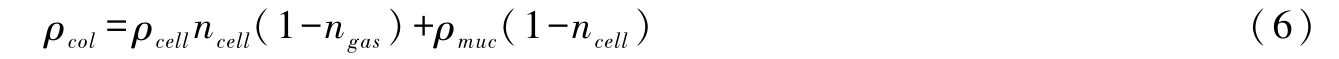
式中:ρcol为群体密度;ρcell为单细胞密度;ncell为微囊藻群体中细胞所占比例;ngas为单细胞中伪空胞所占比例;ρmuc微囊藻黏液密度。
在获取了微囊藻密度变化情况下,常用Stokes沉速公式计算微囊藻垂向迁移速度w和轨迹[79]:

式中:g为重力加速度;ρw为水体密度;D为微囊藻群体直径;μ为水体的黏滞系数;φ为群体形状系数;z(t+Δt)为 t+Δt时刻微囊藻垂向位置;z(t)为 t时刻微囊藻垂向位置;Δt为时间间隔。
式(1)—式(8)主要用于模拟静水中微囊藻群体随光强变化引起的垂向迁移。
动水条件下,模拟微囊藻的迁移还需考虑水流垂向速度u[80-81]的影响,如表3所示。

表3 粒子模型

动水环境中微囊藻迁移不仅受到自主迁移与对流影响,还受到水体的紊动扩散影响,常用一维拉格朗日模型来描述[82]:

式中:i为藻群体编号;Kz[z(i,t)]为垂向扩散系数;为垂向扩散系数的梯度;Rn为服从高斯分布的随机变量。有学者进一步考虑环境因素,如光照和水体紊动对藻群体垂向迁移的影响,提出了改进的拉格朗日模型[83]:

式中:ΔW为服从高斯分布的随机变量;K′为湍流扩散系数K′[z(i,t)]的导数。该模型仍然未考虑水动力对藻群体破裂或聚集的影响。
3.2 连续介质模型微囊藻迁移聚集的连续介质模型主要采用对流扩散型方程模拟藻类浓度分布,如表4所示。这些模型的差异主要体现在处理紊动扩散与垂向迁移项时有所不同。例如俞茜等[77]考虑微囊藻群体的浮力调节能力,建立了光强和紊动扩散协同作用下微囊藻浓度的垂向分布模型:

表4 藻浓度分布的连续介质模型

续表4 藻浓度分布的连续介质模型

续表4 藻浓度分布的连续介质模型

式中:C为微囊藻的垂向浓度;t为时间;V为微囊藻自身在水体中的迁移速度,受光照影响;w为水体垂向紊动流速;z为垂向坐标;D为水体的紊动扩散系数;S为源汇项。该模型中,紊动扩散系数被设为定值。大多数情况下,紊动扩散系数会沿水深方向变化[87]:

式中:DB为底部扩散系数;DS为水体表面扩散系数;wT代表水深;zT代表温跃层厚度。
微囊藻群体尺寸决定其自身垂向迁移能力。通常情况下,大群体易于上浮到水面,形成水面聚集;小群体垂向迁移能力弱,垂向分布相对均匀[77]。研究表明:野外环境中微囊藻群体形态、大小存在显著差异,群体大小可能服从某种分布函数。要更准确模拟微囊藻的垂向分布,需要考虑其群体大小分布对垂向迁移速度的影响。Chien等[82]模拟了非均匀大小微囊藻群体的密度变化和空间分布,结果表明:大群体呈昼夜迁移规律,而小群体需要更长时间才可能到达水面。
此外,还有学者研究了群体密度变化与湍动协同作用下微囊藻垂向浓度分布,所采用微囊藻浓度控制方程如下[78]:

式中:νt为涡黏系数;σT为湍流普朗特数。
微囊藻群体和水流的相互作用十分复杂,尤其是水动力作用下藻群体的聚集、分散[88],现有模型仍不能较好地反映该过程。此外,当前的浓度模型只考虑水动力对藻类的单向作用,忽略藻类对水动力特性的影响,仅适用于低浓度情形。水华暴发期,水体藻浓度较高,微囊藻群体对水体的影响较大,需要考虑水体与藻类的双向耦合效应。
综上所述,在微囊藻迁移分布的数值模拟研究领域,尚需深入开展的研究内容包括:(1)构建考虑外部水动力条件影响尤其是微囊藻群体大量破碎与聚集情形下的群体迁移速度模型;(2)考虑微囊藻群体与水流互馈机制的高浓度微囊藻群体分布动力学模型;(3)光照、温度、营养盐、水动力协同作用下微囊藻分布数学模型;(4)融合定点监测、卫星遥感、航空遥感、数学模型的实时预报方法。
4 结论与展望
在光照、温度、营养盐适宜的条件下,水动力是影响微囊藻水华暴发的关键因素。本文对微囊藻自主迁移的生物学机理、静水中微囊藻自主迁移特性、动水环境中微囊藻迁移聚集的水动力机理、微囊藻迁移分布数值模拟等方面的研究进展进行了梳理与分析,主要结论与展望如下:
(1)微囊藻尤其是微囊藻群体具有一定的垂向自主迁移能力。与微囊藻单细胞相比,微囊藻群体形态、结构更加复杂,不同群体的上浮速度存在显著差异。群体的形态特征与胞外多糖如何影响微囊藻的自主迁移速度还待深入研究。
(2)当垂向水流速度小于或者与微囊藻自身迁移能力相当时,微囊藻自身迁移效应不可忽略;当垂向水流速度远大于微囊藻自身迁移能力时,水动力携带起主导作用。水流不仅可以通过对流、紊动扩散影响微囊藻迁移,还可以通过改变微囊藻群体的形成与破碎,影响微囊藻的自主迁移能力,进而影响微囊藻的时空分布。微囊藻属于典型的柔性体,群体形态各异,其与水流的相互作用是当前研究的难点。
(3)微囊藻水华暴发的水动力机制十分复杂。浅水湖泊、深水水库的微囊藻水华暴发的水动力机理存在较大差别。前者主要受风生流影响,后者主要受异重流影响。当前主要从宏观尺度探讨这两种流态对水华暴发的影响,但微观尺度上水流如何影响了微囊藻的迁移聚集还有待深入研究。
(4)水流与微囊藻群体的相互作用研究主要集中在水流如何通过水动力影响微囊藻的迁移聚集。然而,微囊藻水华暴发时,水体中微囊藻浓度很高,其对水流的影响不可忽略,但相关研究还比较少,有待深入开展。
(5)微囊藻水华的迁移聚集的数值模拟,主要采用个体模型与连续介质模型。已有数学模型虽然考虑了微囊藻群体的自主迁移速度,但忽略了外部水动力条件对微囊藻群体自身迁移速度的影响,尤其是在微囊藻群体大量破碎与形成情形下的迁移速度,与真实情况存在较大差异,有待改善。此外,微囊藻水华暴发时,藻细胞浓度很高,环境水体已不同于纯水体,属于非牛顿流体,相关的数学模型研究也需要深入开展。
(6)微囊藻水华的暴发与光照、温度、营养盐、水动力条件密切相关,要准确模拟微囊藻水华的生消演变,需要结合水动力模型、水质模型、藻类生长模型构建多因素模拟预报模型。此外,结合定点监测、卫星遥感、航空遥感、数据同化、水动力模型的实时预报模型也有待深入研究[94]。
猜你喜欢
中南药学(2022年8期)2022-11-19
建材发展导向(2022年2期)2022-03-08
当代水产(2021年8期)2021-11-04
环境保护与循环经济(2021年7期)2021-11-02
建材发展导向(2021年14期)2021-08-23
环境科学导刊(2021年2期)2021-01-09
当代水产(2019年8期)2019-10-12
当代水产(2019年9期)2019-10-08
江苏农业学报(2019年1期)2019-09-10
中国环境监察(2017年3期)2017-05-14
