
上海环城绿带群落植物多样性20年动态及其对干扰响应*
2023-01-30 08:32贾熙璇张庆费戴兴安商侃侃
中国城市林业 2022年6期
贾熙璇 张庆费 戴兴安 陈 颖 商侃侃
1 中南林业科技大学风景园林学院 长沙 410004
2 上海辰山植物园 上海 201602
3 邵阳市双清区自然资源局 湖南邵阳 422001
城市绿地植物群落的健康与稳定是衡量绿地质量的前提,而群落结构的合理性是关键[1]。动态演变研究是基于自然属性及人工属性叠加的城市环境下的植物群落变化研究,对科学管理城市绿地和生物多样性保育具有理论与实践意义。城市绿地植物群落结构是多重因素综合作用的结果,是“人工形成—自然进程—人工调控”动态过程的体现[1]。欧美国家较早开展城市公园绿地物种组成动态变化研究,如德国柏林研究城市植物组成的变化,伦敦海德公园分析人为活动对植物区系的影响,时间跨度长达100年[2]。1947—1994年纽约佩勒姆湾公园由于人为活动和自然演替过程使25.5%的本地物种消失[3]。Van der Veken等[4]在研究比利时城区1880—1999年植物物种的变化时也同样发现,广适性物种变得越来越普遍,而专适性物种的分布范围减少。捷克皮尔森编制城市植物区系表对120年时间尺度上物种丰富度的变化进行描述[5]。印度班加罗尔研究城市公园树木多样性及历史变化[6]。目前,国内对城市绿地动态研究以植被景观格局动态居多,植物群落结构动态研究较少[7-9],而绿地物种组成动态变化研究更少[10],且对于城市同一绿地群落动态过程和植物组成变化的长时段研究更鲜见报道。
上海申纪港段环城绿带是1999年营造的城市生态防护绿地,采用块状混交方式进行植物配置,养护管理方式比较简单,以日常的除草、修剪和病虫害防治为主。从绿地建设初期开始调查,通过对2001年、2011年和2021年的3个时段植物群落动态变化调查分析,研究新建绿地人工植物群落的演变特征,探讨绿地自然发育过程中人为养护和干扰对植物多样性的影响,为城市绿地植物群落构建和养护管理提供借鉴。
1 研究区概况与研究方法
1.1 研究区概况
作为研究地的环城绿带位于上海市嘉定区申纪港段(31°14′N,121°20′E),面积2.94 hm2。该地块原为船厂和建材堆桟,1999年开始绿地建设,利用76 000 m3建筑垃圾堆填,形成两个小山堆,所用建筑垃圾颗粒较小,结构较好,基本不含有害化学物质及生活垃圾等成分[11]。作为城市生态防护林,追求快速成林,经过20多年的树木生长和群落发育,形成以树木为主体的郁闭绿地。
1.2 研究方法
1.2.1 植物群落调查
在绿地营造初期(2001年)进行植物群落和植物多样性调查,选取以香樟(Cinnamomum camphora)、女贞(Ligustrum lucidum)、无患子(Sapindus saponaria)、日本晚樱(Prunus serrulatavar.lannesiana)、垂柳(Salix babylonica)、雪松(Cedrus deodara)和臭椿(Ailanthus altissima)为优势种的8个典型群落,采用样地调查法,设置面积为200 m2的样方,对样方内所有植物进行分层次全部调查,分别在2011年与2021年进行两次复查,比较群落植物多样性变化。
1.2.2 数据处理
通过计算群落不同层次的物种重要值说明某个种在各层中的优势程度。


α多样性指数用于说明群落内的物种数目和各物种的个体数目分配的均匀度。其中,物种丰富度(S)、Simpson优势度指数(D)、Shannon-Wiener指数(H)、Pielou均匀度指数(E)计算公式如下:

式(4)至式(6)中:ni为种i的个体数目;N为所有种的个体数;Pi为种i的个体数占所有种个体数的比例[12-13]。
2 结果与分析
2.1 物种组成
2001年各植物群落共调查到植物38科63属72种,其中被子植物37科62属71种;2011年共调查到植物43科69属72种,其中被子植物41科67属70种;2021年共调查到植物43科70属78种,其中被子植物41科68属76种。3次调查仅记录到1种裸子植物(雪松),2011年与2021年仅记录到井栏边草(Pteris multifida)1种蕨类植物(表1)。

表1 申纪港段环城绿带维管束植物组成
2.2 生活型组成
将调查样地的植物分为乔木、灌木、草本、藤本类型。20年间样地乔木种类增加7种,但乔木层树种未增加,增加的均为灌草层的乔木更新苗,如朴 树(Celtis sinensis)、复 羽 叶 栾(Koelreuteria bipinnata)、石 楠(Photinia serratifolia)、棕 榈(Trachycarpus fortunei)等;2001年,红 枫(Acer palmatum‘Atropurpureum’)作为主要乔木树种出现,但在后续的调查中并未出现,这与红枫生长不良而调整淘汰有关。3次样方调查(图1)显示:灌木种类无明显变化,2001年灌木主要以更新苗的形式随机出现在样方草本层;2011年增加红瑞木(Cornus alba)、金丝桃(Hypericum monogynum)、结香(Edgeworthia chrysantha)等观赏灌木,相应去除了部分更新苗;2021年新种植了南天竹(Nandina domestica),同时清除了红瑞木、野蔷薇(Rosa multiflora)、火棘(Pyracantha fortuneana)等更新幼株。3次调查中,草本植物种类均占总种数的一半以上,2011年的草本植物种类有所降低。2001年和2021年的藤本植物均为7种,2011年藤本植物为11种,主要为盒子草(Actinostemma tenerum)、萝藦(Cynanchum rostellatum)、葎 草(Humulus scandens)等。
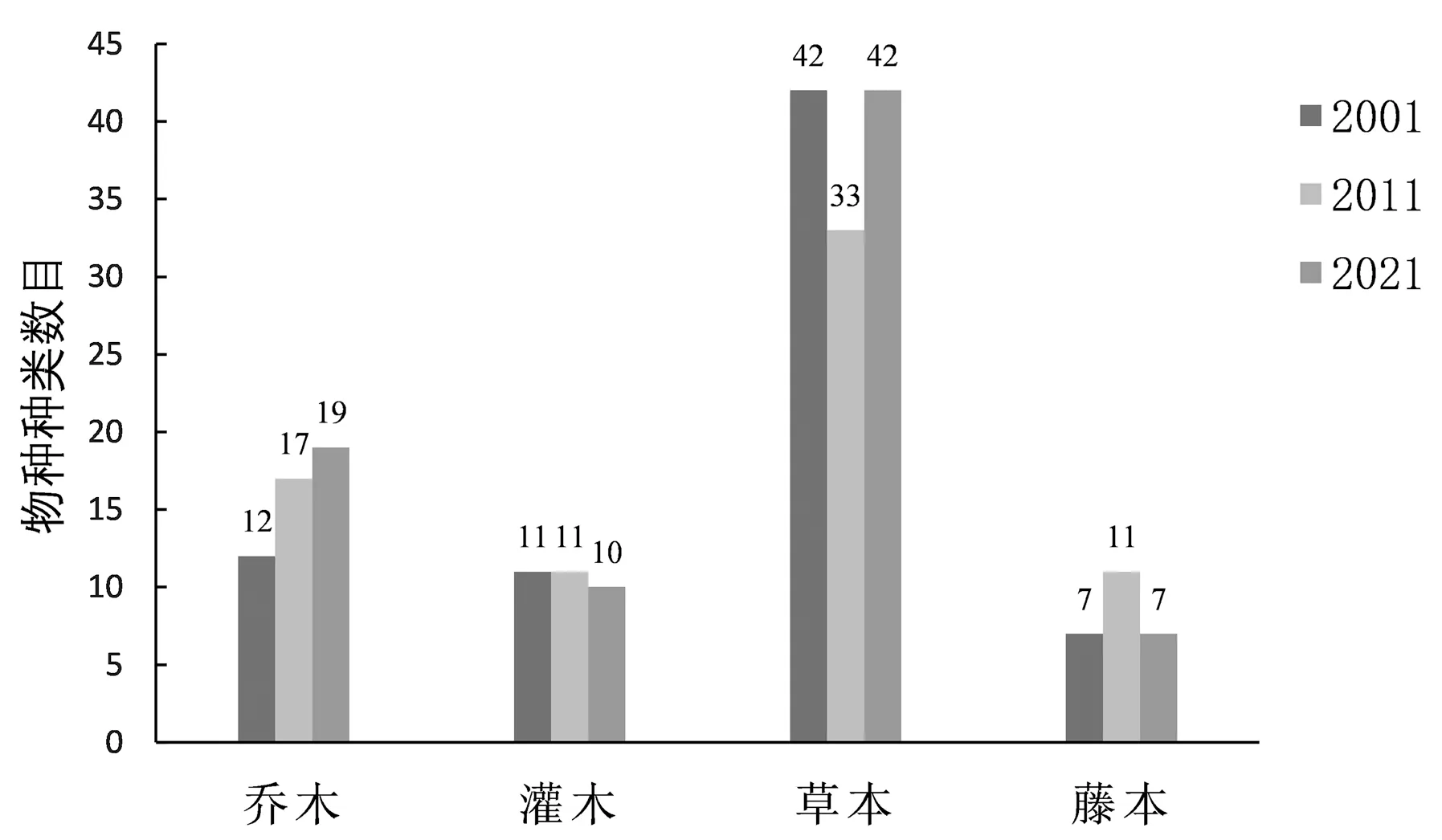
图1 申纪港段环城绿带维管束植物生活型组成
2.3 物种来源组成
由图2可知,各植物群落均以上海原生种为主,3次调查原生种分别占种数的61.11%、58.33%和56.41%,其中草本植物占优势,超过原生种的一半,如地锦草(Euphorbia humifusa)、黄鹌 菜(Youngia japonica)、龙 葵(Solanum nigrum)、马唐(Digitaria sanguinalis)、马蹄金(Dichondra micrantha)、铁苋菜(Acalypha australis)等。20年间栽培种的种类逐渐递增,由最初调查的16种增加至21种,其中新栽植灌木有珊瑚树(Viburnum odoratissimum)、结香、桂花(Osmanthus fragrans)等,草本有大花酢浆草(Oxalis bowiei)、发状薹草(Carex albula)。自然更新物种有复羽叶栾、棕榈、银木(Cinnamomum septentrionale)等乔木。外来归化种3次调查均为6种,3次调查均出现的有垂柳、八角金盘(Fatsia japonica)、鳢肠(Eclipta prostrata)、香附子(Cyperus rotundus);外来入侵种的种类变化不大,白车轴草(Trifolium repens)、红花酢浆草(Oxalis corymbosa)、喜旱莲子草(Alternanthera philoxeroides)、小蓬草(Erigeron canadensis)占据优势。2010年之后,该绿地采用林下密植或满铺山麦冬(Liriope spicata)、扶芳 藤(Euonymus fortunei)、常春藤(Hedera nepalensisvar.sinensis)等地被,外来归化种和外来入侵种的生存空间受到约束;此外,日常的绿地管护也清除了此类草本植物,因此外来种占据空间较小。
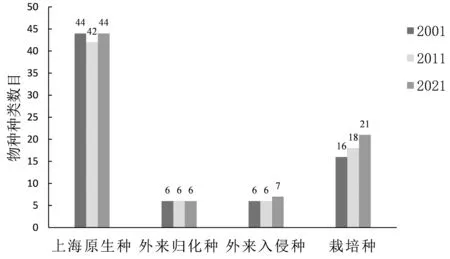
图2 申纪港段环城绿带维管束植物物种来源组成
2.4 各群落物种重要值变化
2.4.1 乔木层
由于调查绿地以块状混交为基本配置方式,乔木层多呈单优种或显著优势种。3次调查(表2)显示,香樟群落(样方2、样方3)、臭椿群落的优势种重要值均高于70%,无患子群落、垂柳群落和女贞群落始终保持单优种,其重要值均为100%。
雪松群落在2001年的调查中优势度并不高,重要值45.35%,其共优种为红枫。随着时间的推移,雪松逐渐占据群落优势地位,2011年后雪松成为样方内的单优种(表2)。
3次调查结果(表2)表明,日本晚樱的重要值逐年降低,而其伴生种香樟的重要值逐渐上升,重要值差值从原有的58.94%缩小至15.98%。这可能是因为日本晚樱长势比香樟缓慢,且对生境要求较高,不能较好适应调查地比较贫瘠的土壤。2001年调查的40株日本晚樱到2021年仅剩余8株。
2.4.2 灌木层
2001年的群落样方多为单一的乔草模式。臭椿群落因臭椿自然更新能力强,臭椿更新幼苗形成林下灌木层。香樟群落(样方3)和雪松群落的灌木层分别种植木槿(Hibiscus syriacus)和瓜子黄杨(Buxus sinica)球(表2)。

表2 各群落乔灌木层重要值
日本晚樱群落和女贞群落在2011年的调查数据显示,灌木层存在海桐(Pittosporum tobira)、桂花等种植灌木和香樟、女贞、桑树(Morus alba)等更新幼苗,但群落在2021年的调查中未发现灌木层。这是因为绿地养护中用吉祥草(Reineckea carnea)、发状薹草、山麦冬替代了原有的栽培植物和更新苗,同时草本层覆盖面积大于85%,限制了其他植物的生长空间。
雪松群落和臭椿群落的灌木层植物为人为设计种植,并保持其植物配置形式和植物种类。2001年在雪松群落调查到3株瓜子黄杨;2011年瓜子黄杨被红瑞木取代,重要值达74.94%;2021年瓜子黄杨又以球状灌木的形式出现,重要值为100%,这表明人为调整决定了灌木层的物种组成。臭椿群落更新苗被新栽植的结香取代,并保留女贞更新苗,2011年和2021年结香的重要值分别为62.96%和40.78%,女贞的重要值分别为20.12%和27.92%。2021年群落内出现新种植的南天竹,重要值为23.06%,这也是样方内结香重要值降低,女贞重要值增长幅度较小的原因。此外,由于香樟林下的木槿生长不良,香樟群落(样方3)中木槿逐渐被淘汰,2011年香樟群落(样方2)人工种植的木槿也在后来被淘汰。
无患子群落、香樟群落、垂柳群落的灌木层多以乔木更新幼苗为主,如香樟、女贞、桑树、构树(Broussonetia papyrifera)等。在人为干预过程中,更新苗被不断清除,又不断自然萌发,对乔木层的结构影响较小,对灌木层的影响也无明显规律,仅香樟群落(样方2)有一株自然发育的构树,呈现城市绿地较为典型的人为干预控制自然发育生长现象。由于灌木层更新幼苗往往受到干扰胁迫,重要值降低;除更新苗外,灌木层还有人为种植的灌木,如八角金盘、海桐、卫矛(Euonymus alatus)等,其长势缓慢,株数稀少。
2.5 各群落多样性指数变化
2.5.1 物种丰富度
由图3可知,各群落乔木层的物种丰富度变化不明显,波动幅度在1~3种左右。香樟群落(样方3)的物种丰富度显著上升,增加的新物种主要为自然发育的更新植株,如银木、桑树、海桐、女贞。2011—2021年各群落灌木层物种丰富度变化波动较大,物种数大幅增加后又略降低。

图3 各群落物种丰富度变化情况
香樟群落灌木层物种丰富度较高,主要有八角金盘、珊瑚树、海桐、火棘等人工种植灌木和香樟、女贞、构树等大量乔木更新幼苗。日本晚樱林下地被覆盖度高,更新幼苗难以生长发育,日本晚樱群落的灌木层物种丰富度在每次调查均为最低值。
草本层的物种丰富度最高,但在第一个10年内物种数逐渐减少,之后的10年变化波动较小。在人为管理下,草本植被受到一定控制,垂柳群落、雪松群落、女贞群落的草本层物种丰富度相对较高,3次调查均名列前茅。臭椿群落草本层的物种丰富度始终较低,这可能是因为臭椿群落林下种植大面积的灌木,灌木覆盖率高达90%,草本植物生长空间较少。
2.5.2 多样性指数
无患子群落、垂柳群落和女贞群落为纯林,乔木层为单一物种。由图4可知,乔木层Simpson优势度指数整体先减后增,而Shannon-Wiener指数呈现先增后减的趋势。2001年和2011年香樟群落(样方3)Simpson优势度指数均最低,Shannon-Wiener指数均最高;2021年日本晚樱群落的数值Simpson优势度指数最低,Shannon-Wiener指数最高。Pielou均匀度指数除日本晚樱群落外均呈下降趋势,2001年香樟群落(样方3)的数值最高(0.998),2011年和2021年日本晚樱群落的数值最高,分别为0.845和0.650。总体而言,申纪港段环城绿带植物群落乔木层的多样性下降,物种数目受到人为影响,物种比较单一。

图4 各群落乔木层多样性指数变化情况
经调查,2001年各群落无灌木层或灌木层仅存在单一物种,因此物种多样性指数缺失。如图5所示:2011年香樟群落(样方2)Simpson优势度指数最低(0.202),Shannon-Wiener指数最高(1.935);2021年则香樟群落(样方3)Simpson优势度指数最低(0.288),Shannon-Wiener指数最高(1.587)。10年内香樟群落(样方3)、垂柳群落Pielou均匀度指数上升,物种分布更加均匀化。2011年调查的日本晚樱群落的Pielou均匀度指数最高,为0.906,2021年垂柳群落的Pielou均匀度指数最高,为0.918。整体上看,香樟群落(样方3)和臭椿群落灌木层多样性指数略微上升,其余群落呈下降趋势。
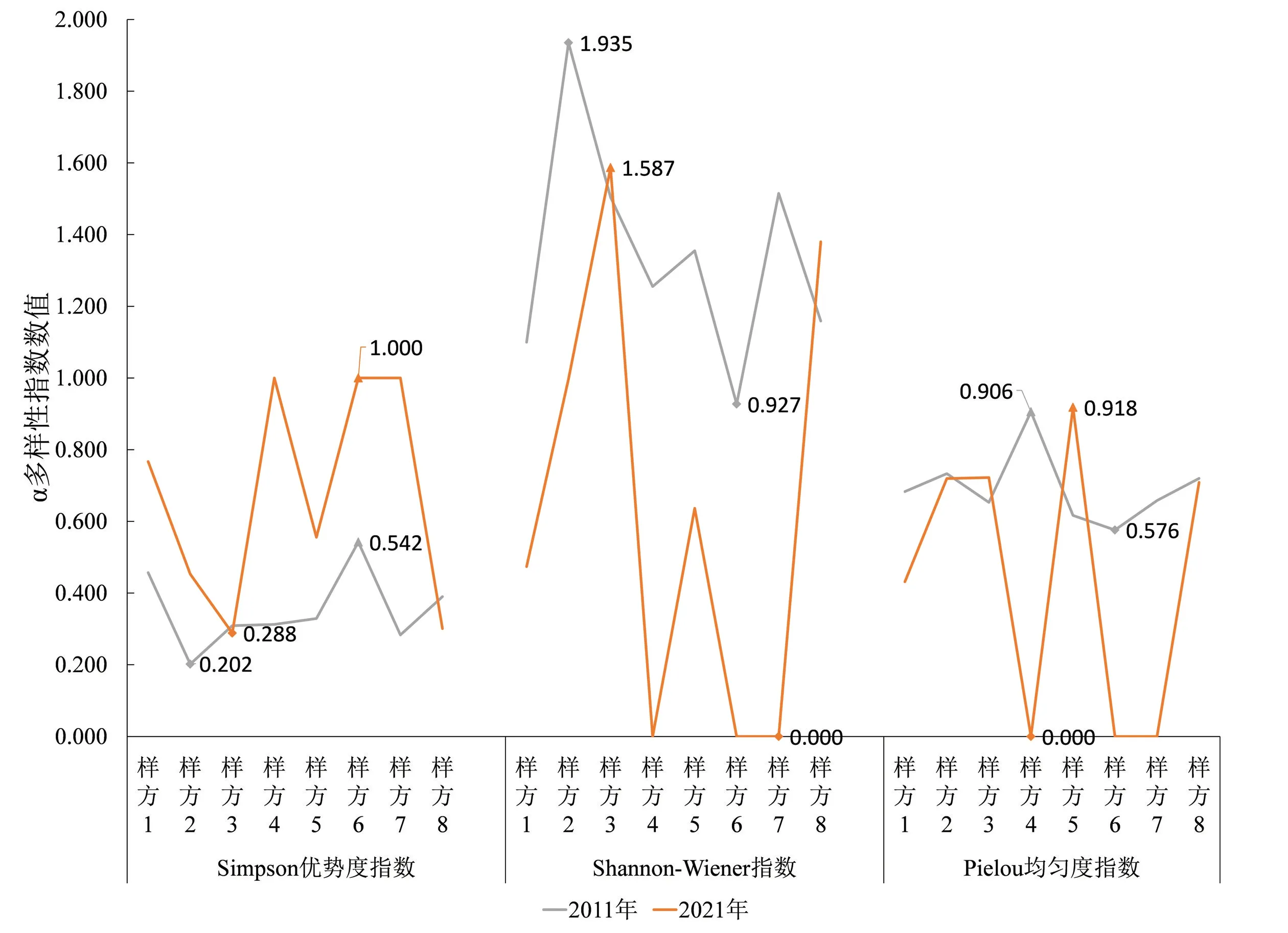
图5 各群落灌木层多样性指数变化情况
调查样地草本层多样性较高,种类组成丰富,群落结构趋于均匀,但仍存在人为干扰的不稳定性,大多数群落的各项指数都呈现先增加后减少的状态。如图6所示:2001年草本层中女贞群落的Simpson优势度指数最低,Shannon-Wiener指数最高,香樟群落(样方2)的多样性指数则相反;2011年女贞群落的Simpson优势度指数最低(0.071),Shannon-Wiener指数最高(2.969),无患子群落的Pielou均匀度指数最高,物种分布最均匀,臭椿群落因人为管护无草本层植物;2021年垂柳群落的草本层多样性指数最高,日本晚樱群落则最低,草本层被吉祥草完全覆盖,少见其他草本植物。
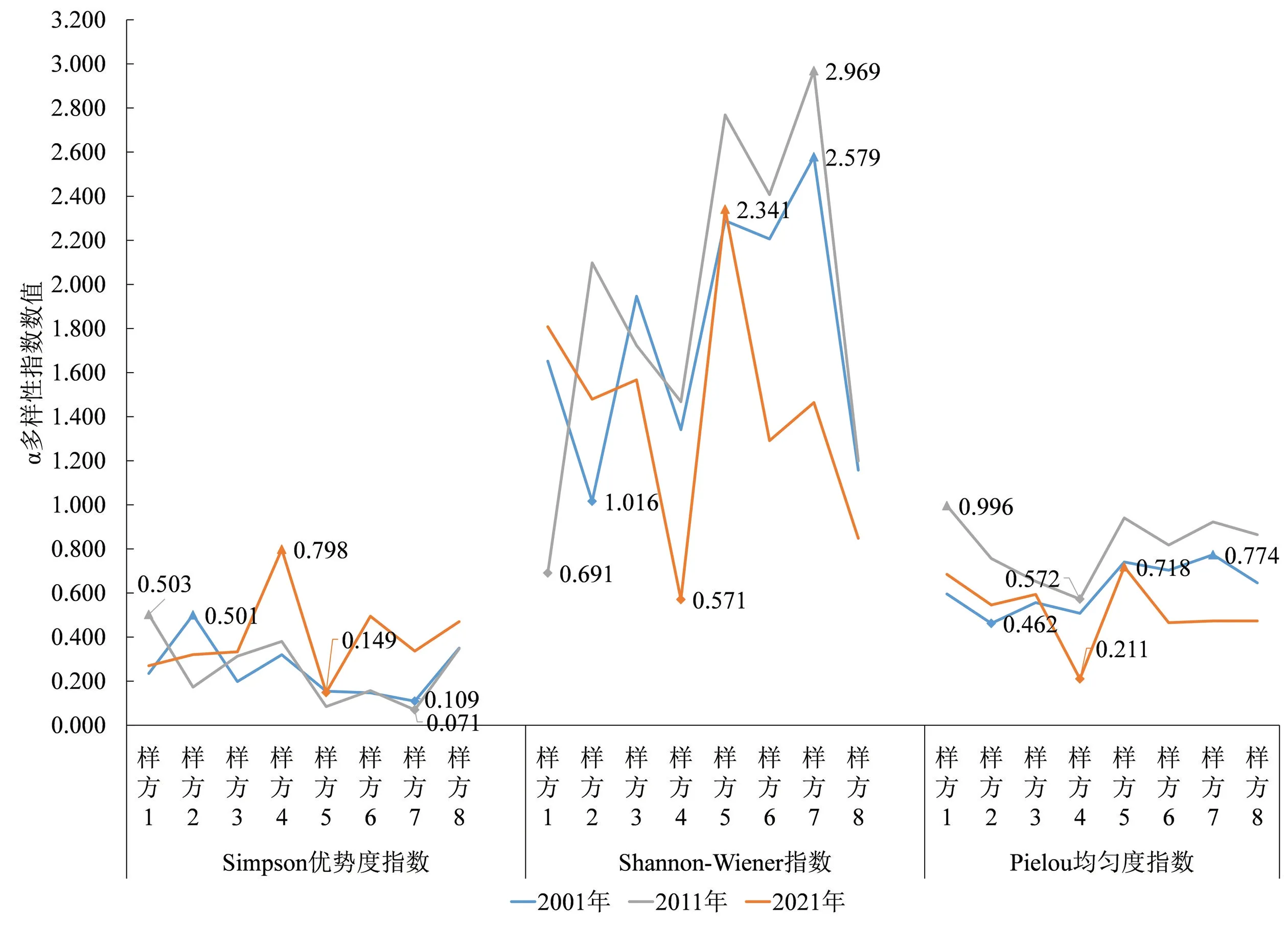
图6 各群落草本层多样性指数变化情况
2.6 人为干扰对植物群落多样性的影响
绿地养护使群落中乔木的数量受到不同程度控制,物种种类变化较小(图4)。由表3可知,除香樟群落(样方3)外,其他群落的乔木第3次调查数目都较前两次调查有所减少或不变。第1次调查初始乔木种植单位面积数量多、密度大,为了给乔木提供更好的生长空间,管理者往往会移除长势较差或死亡的植株。

表3 各群落乔木层优势种的数量变化 株
除乔木层外,林下更新层也受到养护控制,灌草层物种种类和数量受到很大干扰。大部分群落结构都呈现乔草或乔灌单一模式,抑制了种间的物种渗透,更新幼苗难以发育成幼树,幼树也难以长成大乔木,植物多样性受人为干扰波动较大。经调查分析,申纪港段环城绿带林下层人为管护主要体现在以下几个方面:
1)清除型养护。20年间,除垂柳群落和雪松群落外,其他群落下层植被都进行了选择性或全部清除(图4)。在这种简单“一刀切”的养护过程中,绿地能够保持井然有序、层次分明的景观风貌,却抑制自生植物和物种自然更新。由图5和图6可知,2011年大多数群落林下灌草层多样性指数均高于2001年和2021年,这是由于2001年为绿地建设初期,群落物种以人为种植为主,10年后物种的更新和自生植物的繁殖,使群落多样性逐渐增加,而后受到长期频繁的人为清除,群落多样性降低。多数群落灌木层物种数目呈现从零到有再到少的变化,3次调查的多样性指数先上升再下降(表2)。2021年调查结果表明,无患子群落和香樟群落(样方2)清除全部更新幼树幼苗,并对金丝桃、海桐、珊瑚树等树种进行部分清除和调整改造;日本晚樱群落和女贞群落灌木层物种被全部清除。20年间草本层的自生植物也一直被人为控制,在2011年臭椿群落甚至清除了全部的草本植物,同时林下密植结香,草本物种多样性大幅降低。
2)引种型管理。在绿地管护过程中,通过引种植物对绿地进行调整,一方面引进观赏性物种,增加金丝桃、红瑞木、结香、南天竹等灌木和吉祥草、大花酢浆草等草本植物以及扶芳藤等藤本植物;另一方面引进新物种替换长势较差的植株,如雪松群落用瓜子黄杨替换红瑞木,臭椿群落用南天竹代替自然淘汰的结香。此外,引种型管理还能形成提升绿地的某些功能,如香樟群落(样方2)种植珊瑚树取代原隔离护栏,垂柳群落种植水生植物净化水体等。人为引种作为清除型养护的补偿机制,对清除地块进行填补,种植新的林下植被,促进林下地被覆盖度,达到“黄土不见天”的目标。
3)林下密植地被。除垂柳群落外,其余群落林下都逐步采用密植地被的种植形式,即使是臭椿群落灌木层大面积种植结香和女贞,草本层山麦冬和发状薹草的覆盖度也仍然达80%。由表4可知,垂柳群落无密植地被,林下多样性指数均为最高;日本晚樱群落的多样性指数值均为最低,密植地被盖度高达98%。此外,相比双物种密植,单一物种密植对林下多样性的影响更明显,女贞群落和臭椿群落密植山麦冬和发状薹草,其盖度不低于80%,但物种丰富度和林下多样性指数都高于无患子群落。林下密植地被导致植物群落结构干净而整齐,但抑制植被的自生培育潜力,难以形成自然野趣、结构层次丰富的植物群落。
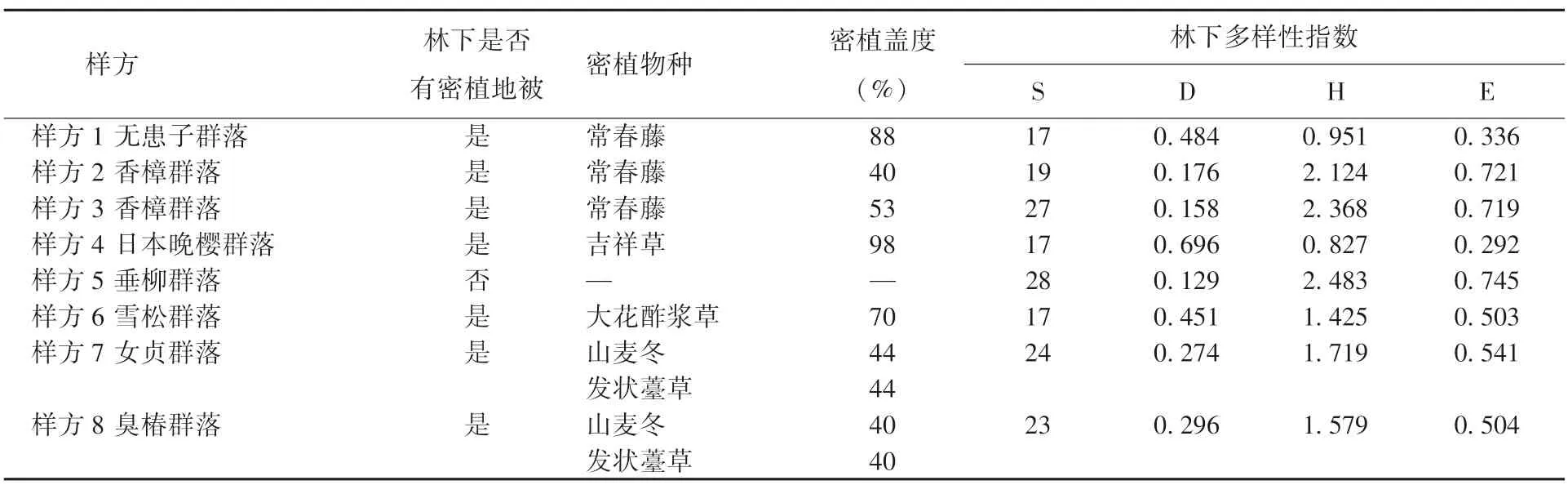
表4 2021年各群落林下物种多样性对比
3 讨论
城市绿地为典型的人为干扰或干预系统,其结构常常是一个未成熟系统的临时框架,不仅在历史中形成,在演替中发展,更在人为干扰中嬗变[14]。因此,养护管理伴随着城市绿地的生长发育,城市绿地总是在自然发育与人为干扰之中不断演化。从图3~6可以看出,没有出现群落各层次植物多样性均明显提高的情况,这些都是人为干预绿地群落发育的结果。Connell的“中度干扰假说”认为中度干扰有利于群落多样性的提高,并使群落达到平稳的状态[15]。2001—2011年,人为干扰使调查区域各群落物种丰富度上升,大多数群落的多样性指数达到20年来最高值,此后绿地在长期干扰下,各项指数又逐渐降低。此外,2011年日本晚樱群落和女贞群落的灌木层群落多样性明显高于无患子群落,但在2011—2021年的清除型管护中,无患子群落采用选择性清除更新苗使其灌木层群落多样性居于中等水平,草本层多样性指数升高。
20年间的各项管护措施都趋于抑制自生植物发育和植被自然更新,这是该绿地植物多样性变化较小,增长缓慢的主要原因。李仓拴等[16]认为,城市植物群落的设计与管理抑制场地自然力和群落演替过程,增加维护成本,削弱群落生态功能。雷金睿等[17]认为,生境差异性和人为干扰是产生城市植物多样性差异的重要因素,少有人为干扰的离海公园植物群落多样性高于滨海公园。其实绿地的人为干预与自然演替虽有冲突,但也能合理调控,协调发育。适度的干扰可以促进生物多样性和生物资源的保护[18]。而且,减少人为管护使绿地植物适度再野化,在自然适应和竞争的过程中,适应性场地环境和繁殖能力高的原生物种更容易发育和定居,借助自然发育提高植物多样性,丰富群落结构[19]。因此,人为干扰是把“双刃剑”,能抑制绿地的自然发育,不利于植物多样性培育,但适当调控群落空间结构,也能促进自然更新苗生长发育,控制入侵植物和恶性杂草,诱导地带性植被发育,有助于提高群落物种丰富度[20],使植被呈现多样的自然形态,丰富绿地植物景观。
4 结论
通过20年间上海申纪港段环城绿带的定期定点调查观测,共调查到维管植物58科113属130种,其中上海原生种有41科74属81种,占总种数的62.31%,与临近绿地的植物群落物种组成和种植方式较为相似。20年间申纪港环城绿带植物群落的种植方式和配置形式没有太大变化,在长期简单化养护管理模式下,维持原有的种植植物。由于少量引进树种,清除自生植被,林下密植地被(如常春藤、发状薹草、山麦冬等)等干扰,各层植物群落多样性指数整体变化幅度平稳。绿地建成至今,控制自生植物的养护方式抑制城市绿地植物群落多样性变化,人为干预导致植物群落结构变化较小,基本反映了上海普通城市绿地的演变特征。
猜你喜欢
林业勘查设计(2022年1期)2022-02-15
乡村科技(2021年17期)2021-10-20
安顺学院学报(2021年4期)2021-09-16
西安交通大学学报(社会科学版)(2021年2期)2021-04-06
北方音乐(2019年6期)2019-07-16
摄影之友(影像视觉)(2019年2期)2019-03-05
现代园艺(2017年23期)2018-01-18
小小说月刊·下半月(2015年12期)2015-05-14
诗歌月刊(2014年1期)2014-03-11
祝您健康(1989年2期)1989-12-30
