
CO2加富与LED光对黄瓜幼苗抗氧化特性的影响
2023-01-12 05:49姜玮莉康红旗孙启颖周璇孔静雯孙坡李清明刘彬彬
山东农业大学学报(自然科学版) 2022年5期
姜玮莉,康红旗,孙启颖,周璇,孔静雯,孙坡,李清明,刘彬彬
CO2加富与LED光对黄瓜幼苗抗氧化特性的影响
姜玮莉1,康红旗1,孙启颖1,周璇1,孔静雯1,孙坡1,李清明2*,刘彬彬3*
1. 山东农业大学园艺科学与工程学院, 山东 泰安 271018 2. 中国农业科学院都市农业研究所, 四川 成都 610299 3. 作物生物学国家重点实验室, 山东 泰安 271018
以‘津优35号’黄瓜品种为试材,采用裂区试验设计,主区因素为CO2浓度处理,分为大气CO2浓度(≈ 400 μmol·mol−1)和加富CO2浓度(≈ 800 μmol·mol−1)2个水平。裂区因素为光配方处理,分为白光(W)、红蓝光(RB=7:3)和红蓝光加UV-A(RBU=7:3+20 μmol·m-2·s-1)3个光配方,研究了CO2加富与LED光配方对黄瓜幼苗抗氧化性的影响。试验结果表明:CO2加富和RBU处理可以显著提高SOD、POD、CAT、APX、GPX、GR、DHAR等抗氧化酶的活性以及可溶性糖、游离氨基酸、可溶性蛋白和脯氨酸等渗透调节物质的含量,提高ASA、GSH含量以及ASA/DHA、GSH/GSSG的比值,促进ASA-GSH循环,从而降低RBU处理下MDA及ROS的含量,减缓添加UV-A后对黄瓜幼苗所造成的氧化损伤。综上所述,CO2加富和RBU处理可以通过提高抗氧化酶活性以及渗透调节物质的含量来减少活性氧的积累,提高植物的抗氧化性,从而有利于幼苗的健壮生长,培育壮苗。
黄瓜; CO2加富; LED光配方; 抗氧化性
随着工业化进程的加快,大气中CO2浓度不断升高[1,2]。预计到本世纪末,大气中二氧化碳的浓度将增加一倍[3]。CO2作为植物光合作用的重要原料,其浓度高低将直接影响植物的生长发育及生理代谢过程[4]。适宜的光环境是影响植物工厂化育苗的关键因素[5],包括光强、光质、光照时数和光的分布四个方面,其中光质是影响蔬菜细胞内活性物质生物合成的主要因素,不仅影响植物的生长发育,而且对植物的物质代谢也起着至关重要的作用,可以作为调控蔬菜的抗氧化活性的措施之一[6,7]。
有研究表明,CO2加富能提高植物的抗病性,降低农药的使用量[8]。在增施CO2后,植株生长健壮,叶片肥厚,SOD、POD等抗氧化酶活性明显增强,MDA含量和电解质渗漏率降低,显著提高了植株的抗氧化性[9,10]。其原因是CO2浓度升高能促进光合作用,提高光合产物,充足的光合产物加强了抗坏血酸等物质的合成,提高了抗氧化酶的活性,从而提高植物的抗氧化能力[11]。另外,CO2浓度升高增加了CO2/O2的比例,抑制了C3植物的光呼吸,减少了以O2作为底物形成的活性氧,进而降低了植物体内ROS的积累[12]。并且高浓度的CO2也促进了碳水化合物的合成,使叶片可溶性糖等渗透调节物质含量增高,从而提高作物的抗性[13]。ASA-GSH循环是植物体内重要的非酶促抗氧化系统,它在帮助植物抵御逆境胁迫时发挥着重要的作用。Muthusamy M等研究发现,CO2加富能够显著提高十字花科蔬菜幼苗的ASA含量和抗氧化性能[14]。王骞春等的研究结果表明,CO2加富除了可以显著提高ASA的含量,还可以显著提高GR、APX等ASA-GSH循环关键酶的活性,从而提高植物的抗氧化能力[15]。
植物的衰老过程受温度和光照等环境因子的影响,近年来,有许多国内外学者以光质作为环境调控因子,探讨其对植物抗氧化系统的影响[16]。蒲高斌等的研究结果表明,红光和蓝光处理与白光处理相比可以显著提高POD、SOD及APX等抗氧化酶的活性[17]。而王虹、高美芳等人的研究进一步表明,蓝光和UV-A能够提高作物抗氧化能力,诱导抗氧化酶相关基因的表达,提高SOD、POD、CAT、APX等抗氧化酶的活性,减少叶片中MDA以及ROS的积累[18,19]。而绿光、黄瓜则会抑制抗氧化酶的活性,加速植物的衰老,说明植物体内抗氧化酶的活性受光质影响较大。
黄瓜(L.)是我国北方地区设施栽培中的常见蔬菜作物,栽培面积已超过70万hm2且呈稳定增长趋势,随着黄瓜生产的不断扩大及人工成本的不断升高,优质种苗的需求量也在不断增加[20]。ROS是一类具有很强氧化能力的含氧物质,植物体内若产生并积累过多的ROS将会破坏植物组织结构与功能[21];而当浓度较低时,ROS就可以作为植物体内一类重要信号分子,参与调节植物的生长发育和逆境胁迫响应,提高植物的抗氧化性从而提高幼苗的健壮度[22]。本试验通过研究CO2加富和LED光配方对黄瓜幼苗ROS含量的调节,旨在探究CO2加富和LED光配方提高黄瓜幼苗抗氧化性的生理机制,以期找到适于黄瓜幼苗健壮生长的环境,为培育优质黄瓜壮苗提供理论依据。
1 试验设计与方法
1.1 试验材料
本试验于2021年8月到2021年11月在山东农业大学园艺试验站进行,供试材料是‘津优35号’黄瓜,购自天津市科润黄瓜研究所。供试的LED光源购买自青岛索广电器有限公司,根据试验要求设计成形状为长(120 cm)×宽(10 cm)的方形植物生长灯,包含白光、红光、蓝光、UV-A四种光质,根据试验要求按照不同比例搭配成LED组合光。
黄瓜种子经过浸种催芽后,挑选出芽一致的种子点播于50孔塑料穴盘中,一穴一粒,基质配制比例为草炭:蛭石:珍珠岩=3:1:1。待幼苗长到一叶一心之后挑选出长势一致的黄瓜幼苗移栽到直径为11 cm的营养钵中,在日光温室缓苗一至两天后移至不同光配方下进行处理。
1.2 试验设计
试验采用裂区设计,主区因素是CO2处理,分为大气CO2浓度(≈400 μmol·mol-1)和加富CO2浓度(≈800 μmol·mol-1)两个水平,裂区因素是不同光配方处理,分为白光(W)、红蓝光(RB=7:3)、红蓝光加UV-A(RBU=7:3+20 μmol·m-2·s-1)3个水平,光周期为12 h·d-1,光强为300 μmol·m-2·s-1,每个处理之间用遮光布隔开。试验在自行设计的顶通风式塑料拱棚(长6 m,宽6 m,脊高2.6 m)内进行,棚内设有环境控制系统。CO2由液态CO2钢瓶供给,浓度由CO2自动控制系统控制(Auto 2000,北京奥托)。当CO2浓度低于目标浓度时,电磁阀会自动开启进行补充CO2,使浓度达到800±50 μmol·mol-1。
1.3 试验方法
(1)叶片丙二醛、ROS含量的测量丙二醛(MDA)含量采用硫代巴比妥酸(TBA)显色法测定,超氧阴离子自由基(O2.-)产生速率采用羟胺氧化法测定[23];过氧化氢(H2O2)含量按照试剂盒说明测定(科铭生物科技有限公司,中国苏州)。
(2)叶片渗透调节物质含量的测量脯氨酸含量采用茚三酮-磺基水杨酸法测定[24];游离氨基酸含量采用茚三酮显色法测定[25];可溶性糖含量采用蒽酮比色法测定[25];可溶性蛋白含量采用考马斯亮蓝染色法测定[24]。
(3)叶片抗氧化酶活性的测量用NBT法测定超氧化物歧化酶(SOD)、用愈创木酚法测定过氧化物酶活性(POD)、用紫外吸收法测定过氧化氢酶(CAT)活性[26]。
(4)叶片ASA、DHA、GSH、GSSG含量及ASA-GSH循环关键酶活性的测量还原型抗坏血酸(ASA)含量采用2,2-二联吡啶的比色法测定[26];还原型谷胱甘肽(GSH)含量采用5,5-二硫代双(2-硝基苯甲酸)比色法测定[27];脱氢抗坏血酸(DHA)、氧化型谷胱甘肽(GSSG)含量按照试剂盒说明测定(科铭生物科技有限公司,中国苏州)。抗坏血酸过氧化物酶(APX)活性参照吴强盛的方法[26];谷胱甘肽过氧化物酶(GPX)、谷胱甘肽还原酶(GR)、脱氢抗坏血酸还原酶(DHAR)活性按照试剂盒说明测定(科铭生物科技有限公司,中国苏州)。
1.4 数据处理
利用Microsoft Excel 2019和DPS 15.10软件对数据进行处理和显著性分析,并采用SigmaPlot 10.0软件绘图。
2 结果与分析
2.1 CO2加富与LED光配方对黄瓜幼苗叶片丙二醛及ROS含量
植物在受到逆境胁迫时,体内往往会产生大量的丙二醛和活性氧(ROS)。ROS是植物在生化反应中生成的具有强氧化活性、能够加速植物衰老的物质。MDA是膜脂过氧化产物之一,可以反应细胞的受损程度[19]。MDA含量大幅增加会使细胞膜脂过氧化作用加剧,从而加重膜的损伤。从图1中可以看出,在RB处理下,超氧阴离子产生速率、丙二醛和过氧化氢的含量都显著低于W处理,而RBU处理提高了超氧阴离子产生速率、丙二醛以及过氧化氢的含量,说明添加UV-A之后黄瓜幼苗受到了一定程度的胁迫。而CO2加富处理均降低了RB和RBU处理下的超氧阴离子的产生速率、丙二醛和过氧化氢的含量,说明CO2加富处理可以减少各个光配方处理下的活性氧物质的积累,提高黄瓜幼苗的抗氧化能力。
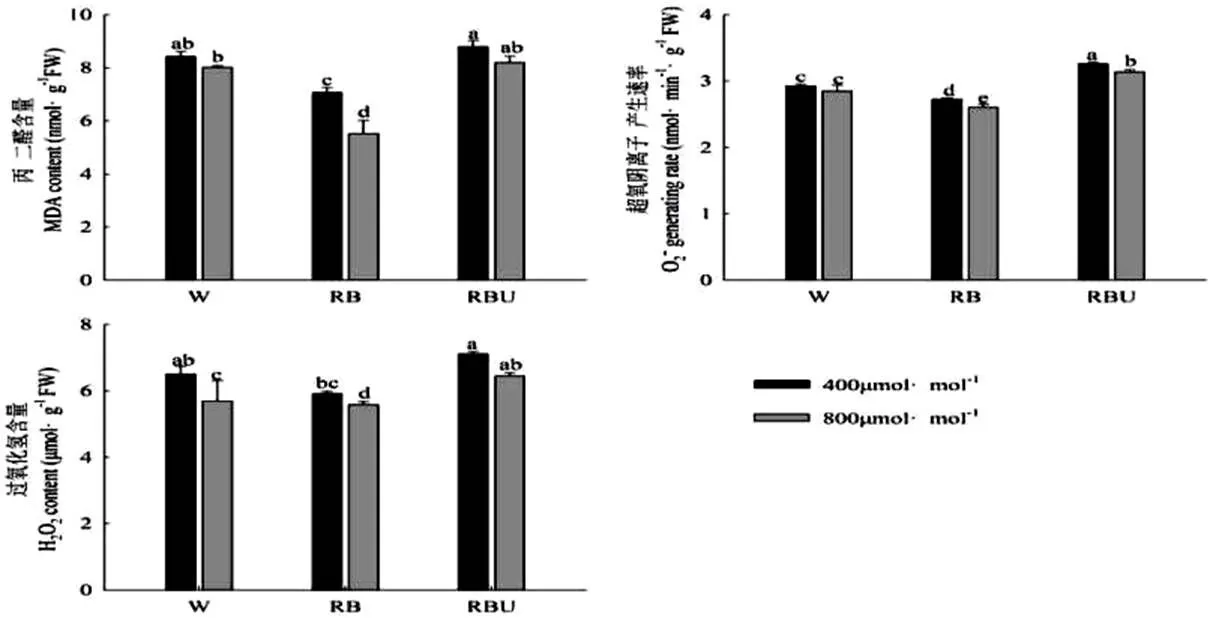
图1 CO2加富与LED光配方对黄瓜幼苗丙二醛及活性氧含量的影响
2.2 CO2加富与LED光配方对黄瓜幼苗叶片渗透调节物质含量
在植物中,渗透调节物质往往可以帮助植物抵御逆境。由图2可知,RB和RBU处理与W处理相比,显著提高了脯氨酸、可溶性蛋白和可溶性糖的含量,CO2加富处理进一步提高了RB和RBU处理下的脯氨酸和可溶性糖的含量,分别提高了22.76%、20.17%、72.28%、19.7%。但是对可溶性蛋白含量的影响不大。在不同光配方处理下,游离氨基酸的含量在RBU处理下的含量最高,在RB处理下的含量最低,并且CO2加富处理进一步降低了不同光配方处理下游离氨基酸的含量。
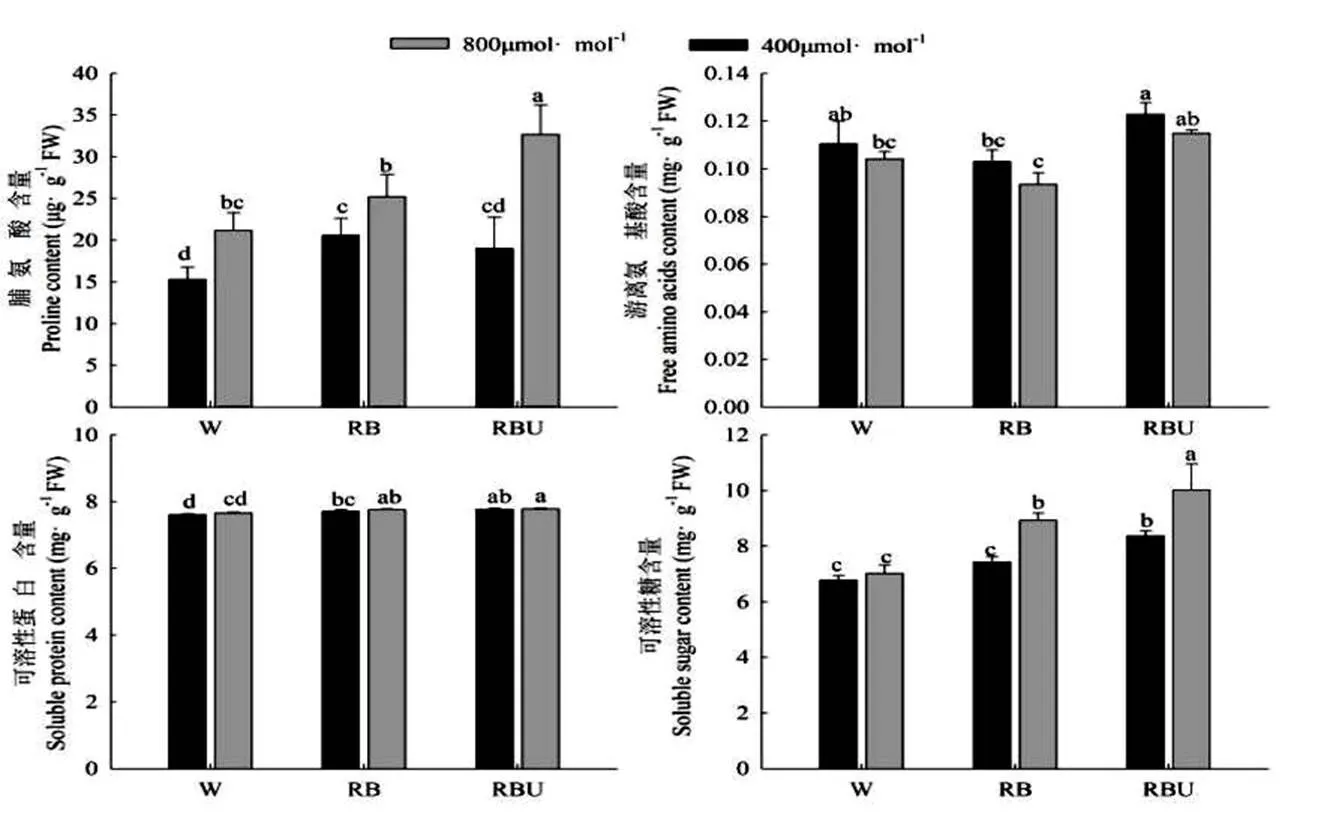
图2 CO2加富与LED光对黄瓜幼苗渗透调节物质含量的影响
2.3 CO2加富与LED光配方对黄瓜幼苗叶片抗氧化酶活性
由图3可以看出,在大气CO2浓度下,与W处理相比,RB和RBU处理显著提高了过氧化氢酶(CAT)、过氧化物酶(POD)和超氧化物歧化酶(SOD)的活性,说明RB和RBU处理在大气CO2浓度下可以通过提高抗氧化酶的活性来提高黄瓜幼苗的抗氧化能力。而CO2加富处理进一步提高了各个光配方处理下的抗氧化酶的活性,其中过氧化氢酶(CAT)和过氧化物酶(POD)的活性在RBU处理下达到最大值,与W处理相比提高了7.19%、51.3%,SOD的活性在RB处理下活性到达最大值,与W处理相比提高了98.05%,差异达到显著水平。
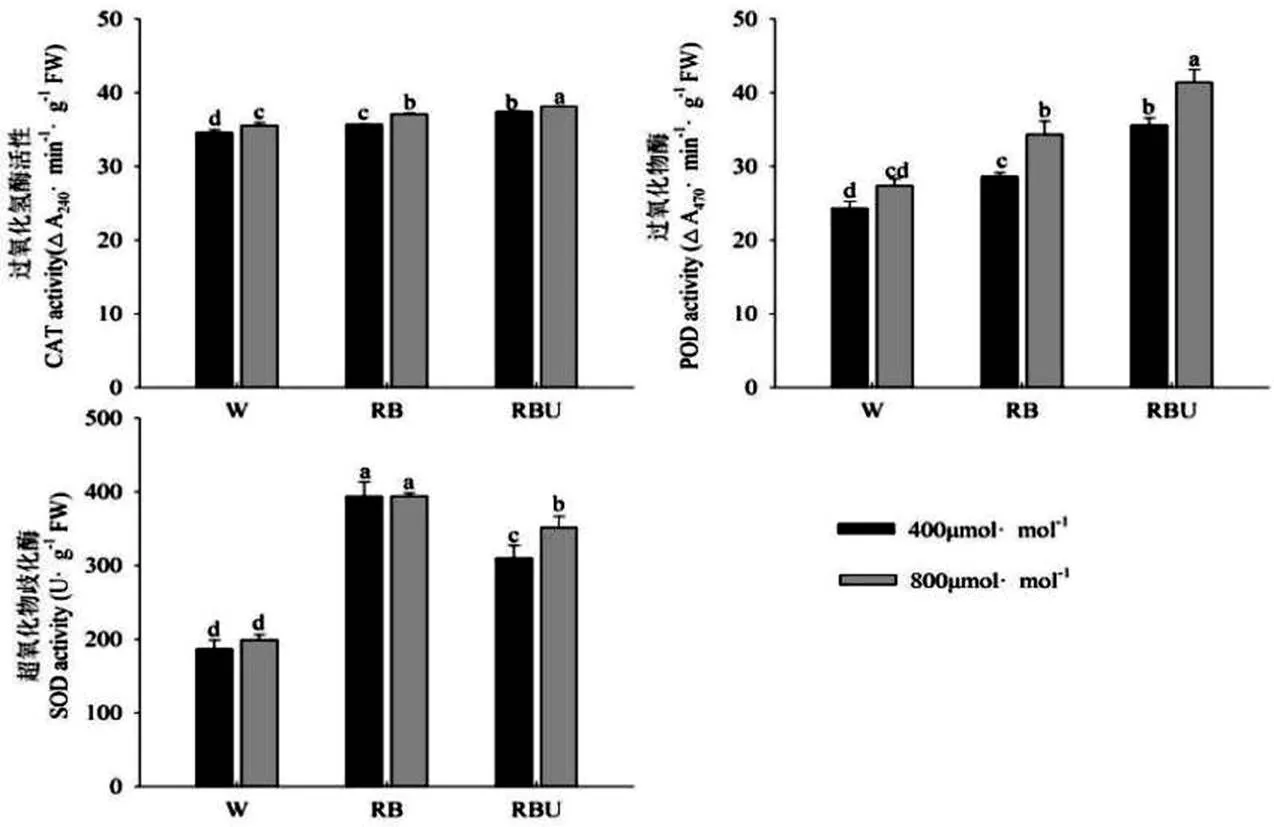
图3 CO2加富与LED光对黄瓜幼苗抗氧化酶活性的影响
2.4 CO2加富与LED光配方对黄瓜幼苗叶片ASA、DHA、GSH、GSSG含量
由图4可以看出,不管是在大气CO2还是加富CO2浓度下,RB和RBU处理下的ASA、DHA、GSH、GSSG的含量均显著高于W处理,而CO2加富处理进一步提高了RB和RBU处理下的ASA、DHA、GSH、GSSG含量,其中在RBU处理下的含量均达到最大值,与W处理相比提高了30.56%、23.38%、28.13%、23.74%,这说明在红蓝光的基础上添加20 μmol·m-2·s-1UV-A可以显著提高ASA、DHA、GSH、GSSG的含量。
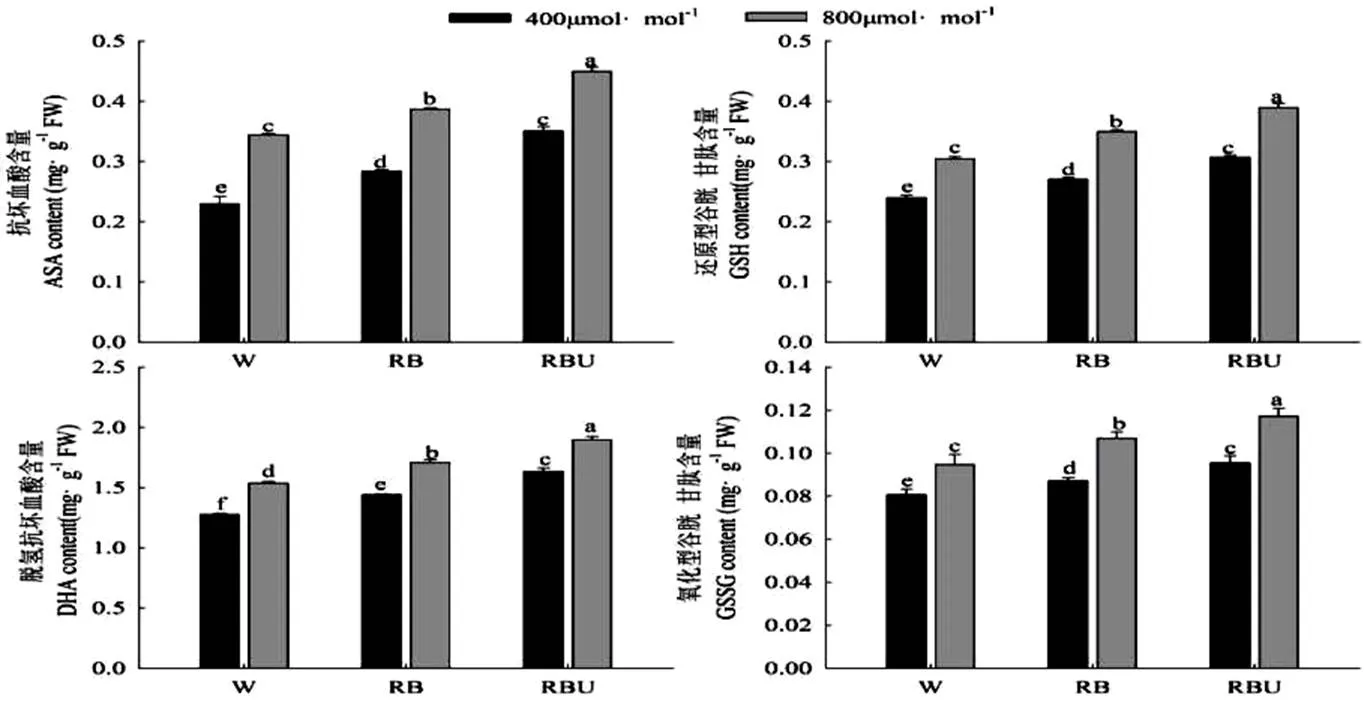
图4 CO2加富与LED光对黄瓜幼苗ASA-GSH循环抗氧化剂含量的影响
2.5 CO2加富与LED光配方对黄瓜幼苗叶片ASA/DHA、GSH/GSSG比值
ASA/DHA、GSH/GSSG的比值可以反应植株的受损程度,比值越低说明植株的防御系统越弱。从图5中可以看出,RB和RBU处理下的ASA/DHA、GSH/GSSG的比值显著高于W处理,并且CO2加富处理进一步提高了RB和RBU处理下的ASA/DHA、GSH/GSSG的比值,在RBU处理下达到最大值,说明CO2加富和RBU处理可以促进ASA-GSH循环的代谢速率,从而提高黄瓜幼苗叶片的抗氧化能力。
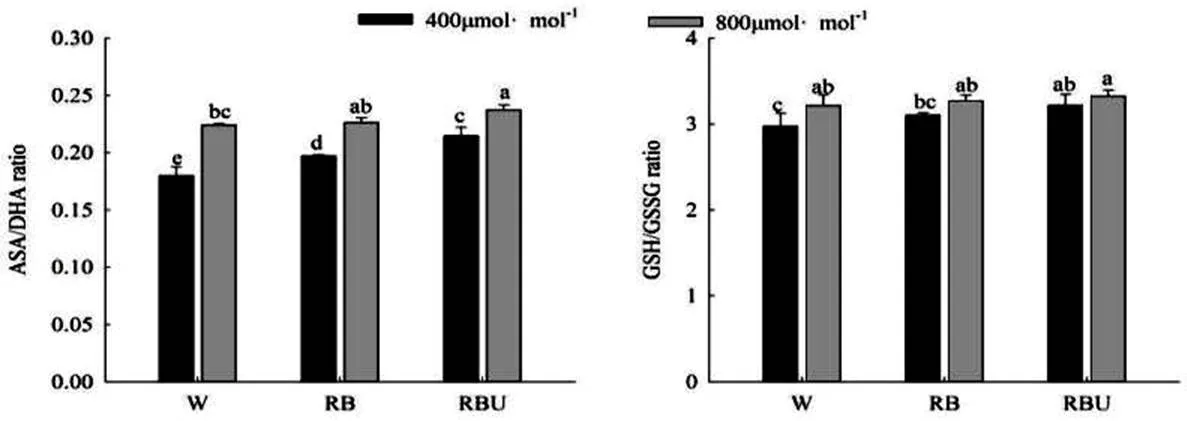
图5 CO2加富与LED光对黄瓜幼苗ASA/DHA、GSH/GSSG的影响
2.6 CO2加富与LED光配方对黄瓜幼苗叶片ASA-GSH循环关键酶活性
由图6可以看出,在大气CO2浓度下,RB和RBU处理均显著提高了抗坏血酸过氧化物酶(APX)、谷胱甘肽过氧化物酶(GPX)、脱氢抗坏血酸酶(DHAR)、谷胱甘肽还原酶(GR)的活性,其中GPX、DHAR、GR的活性均在RBU处理下达到最大值,与W处理相比提高了57.02%、59.82%、15.71%,APX在RB处理下达到最大值,与W处理相比提高了18.3%。而CO2加富处理进一步提高了各个光配方处理下的APX、GPX、DHAR、GR的活性,并且均在RBU处理下达到最大值,说明CO2加富配合RBU处理可以显著提高ASA-GSH循环的关键酶活性,从而提高黄瓜的抗氧化能力。
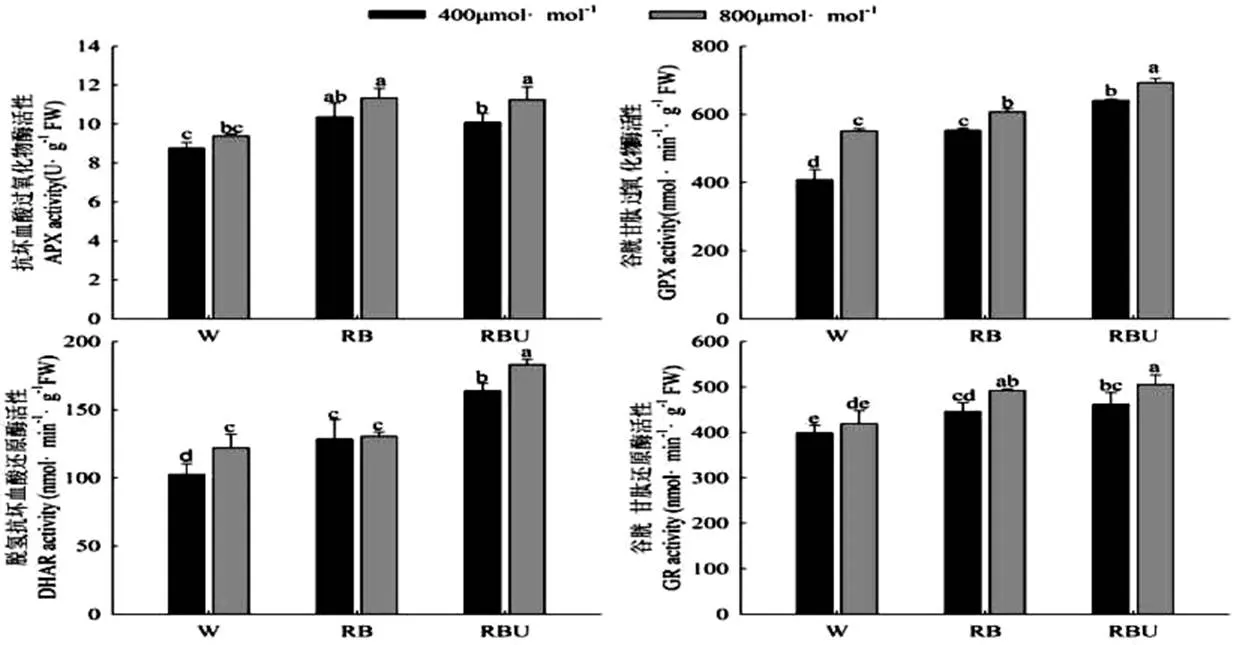
图6 CO2加富与LED光对黄瓜幼苗ASA-GSH循环关键酶活性的影响
3 讨论
植物在生长发育过程中受到非生物胁迫时会产生大量的ROS,而ROS积累过量会对细胞造成损伤,最终可能导致植物的坏死[18]。而遭受可承受或较为温和的胁迫时,ROS可作为一种信号分子,参与调节植物的生长发育和逆境胁迫响应,激活氧化防御机制来适应胁迫环境[22]。O2.-和H2O2是植物细胞代谢过程中的产物,具有很强的氧化性,会使细胞膜发生膜脂过氧化损伤,破坏细胞膜结构,造成膜损伤。而MDA是植物细胞膜脂过氧化作用的最终产物,对细胞膜具有毒害作用[28]。在本试验中,在大气CO2处理下,RB处理下的MDA、O2.-以及H2O2的含量都显著低于W处理,而RBU处理下MDA、O2.-、H2O2含量均显著高于W处理(图1),并且差异显著,说明添加UV-A后对黄瓜幼苗造成了一定程度上的胁迫,诱导黄瓜幼苗叶片产生了大量的ROS和丙二醛,而积累过多的ROS和丙二醛会造成严重的氧化应激和膜脂过氧化损伤,对植物生长发育造成不良影响[29]。当植物体内积累过多的ROS时,会激活植物体内的抗氧化系统,从而提高抗氧化酶的活性来修复由逆境造成的活性氧代谢失衡,从而清除植物体内的ROS。SOD、CAT、POD是植物抗氧化系统的关键酶,其活性的高低可以反应植物受逆境影响的程度。SOD是抗氧化系统清除活性氧的第一道防线,可催化O2.-转化为H2O2和O2,CAT和POD在清除ROS方面也发挥着重要的作用,可以将H2O2反应生成水[30]。在正常生理活性下,三种酶相互协同合作,可以维持ROS保持在一个稳态水平。在本试验中,大气CO2处理下的RB处理显著提高了CAT、POD、SOD等抗氧化酶的活性(图3),这与刘福霞等人研究结果一致[31]。在RB处理的基础上添加UV-A之后酶活性得到进一步提高,说明在添加UV-A黄瓜受到一定程度的胁迫之后,体内积累的ROS诱导黄瓜幼苗对UV-A胁迫做出了适应性反应,提高抗氧化系统关键酶的活性,减少了ROS的积累,降低了ROS对细胞膜的损伤,从而提高植物的抗氧化能力。而不同光配方处理在CO2加富之后显著降低了MDA、H2O2的含量以及O2.-的产生速率(图1),这与既往研究结果一致[28,32,33]。这是因为CO2加富可以提高叶绿体CO2/O2的比率,增加CO2的同化[15],减少因O2作为电子受体而形成的活性氧,减轻O2.-的毒害作用,而且CO2浓度的提高还可以降低光呼吸形成的H2O2[34,35]。同时,CO2加富处理显著提高了不同光质处理下的CAT、POD、SOD等抗氧化关键酶的活性,从而降低了丙二醛和ROS含量。
多种逆境都会对植物产生直接或间接的水分胁迫,而脯氨酸、可溶性糖、可溶性蛋白、游离氨基酸等渗透调节物质可以在植物受到逆境时对胁迫作出适应性反应。维持细胞正常膨压,保护酶活性,维持气孔开放,促进植物的光合作用,从而提高植物的抗氧化能力[36,37]。其中,脯氨酸是一种抗氧化剂,可清除细胞内部的ROS[38]。本试验结果表明,RB和RBU处理均可以显著提高大气CO2浓度下脯氨酸、可溶性蛋白和可溶性糖的含量,并且CO2加富处理可以进一步提高不同光质处理下三种渗透调节物质的含量(图2),说明在CO2加富和RB、CO2加富和RBU组合处理下,有利于渗透调节物质的积累,降低细胞水势,提高细胞吸水和保水的能力从而提高抗氧化能力,减少逆境的损伤。游离氨基酸含量与上述三种物质含量趋势相反,在RB处理下显著降低,而在RBU处理下含量显著高于W处理(图2)。这可能是因为RB处理提高了抗氧化酶活性,降低了ROS含量,从而对游离氨基酸的需求量减少。而CO2加富处理进一步降低了RB和RBU处理下游离氨基酸的含量(图2),这与Högy P等和Li SH等的研究结果一致[39,40],可能是游离氨基酸降低的这部分是非结构性碳水化合物增加的结果[41]。
ASA-GSH循环是植物体内清除细胞活性氧的重要途径之一,能够提高植物的抗逆性[42]。该循环系统在植物清除活性氧,抵抗氧化胁迫,促进细胞生长等方面具有重要功能。抗坏血酸(ASA)和谷胱甘肽(GSH)是普遍存在于植物体内的小分子水溶性非酶抗氧化物质,可以清除植物体内活性氧,抵抗氧化胁迫[43]。他们的存在状态也会影响其功能,当受到一定程度的氧化损伤时,植物可将氧化型抗坏血酸和氧化型谷胱甘肽都转化成还原型状态,以减轻损害[44]。魏国芹等研究认为较高的ASA/DHA和GSH/GSSG能保证ASA-GSH循环代谢正常运转,提高植株清除ROS能力[45]。在本试验中,RB和RBU处理显著提高了ASA、DHA、GSH、GSSG的含量以及ASA/DHA、GSH/GSSG的比值,并且在CO2加富处理下得到进一步提高(图4,5),说明CO2加富和RB以及CO2加富和RBU的组合处理可以显著提高植株体内抗氧化剂的含量,以减轻逆境胁迫。APX、GPX、DHAR、GR是ASA-GSH循环的关键酶,对维持ASA-GSH循环的有效运行具有重要的作用。ASA在APX的催化作用下与H2O2反应,将其分解生成H2O,从而达到清除H2O2的目的(曹碧珍,2021),而ASA则被氧化成MDHA,MDHA极不稳定,进一步再氧化脱氢生成稳定的DHA。而DHA又以GSH为底物在DHAR酶的催化作用下重新生成ASA,而此反应所产生的GSSG在NADPH参与下又被GR催化还原成GSH[46-48],因此,GR活性直接影响细胞GSH库的水平[49]。本试验结果表明,在RB和RBU处理下的APX、DHAR、GPX、GR活性均显著高于W处理,而CO2加富处理进一步提高了各个光配方处理下的酶活性,其中在RBU处理下APX、DHAR、GPX、GR活性最强(图6)。说明CO2加富和RBU的组合处理可以有效提高ASA-GSH关键酶活性,从而使ASA-GSH循环代谢正常有效地运行,提高清除活性氧的能力。
综合以上讨论说明,植物在受到轻微胁迫的时候,植物体内产生的ROS可以起到信号调节的作用,刺激植物启动体内的抗氧化系统,提高抗氧化酶的活性以及加速渗透调节物质在黄瓜体内的积累以维持细胞正常膨压。加速活性氧的清除,从而使植物体内ROS的产生和清除达到一个平衡状态,减轻膜损伤,维持植物的正常生长发育。
4 结论
综上所述,黄瓜幼苗在CO2加富和RBU处理下抗氧化性最高。CO2加富和RBU处理可以通过提高渗透调节物质以及ASA、DHA等抗氧化剂的含量,SOD、POD、CAT、APX等抗氧化酶的活性,来降低RBU处理下的ROS和丙二醛,从而提高幼苗的抗氧化能力,有利于培育壮苗。
[1] 刘晓聪,董家华,欧英娟.大气中CO2与O3浓度升高对植物光合作用影响研究[J]环境科学与管理,2016,41(3):152-155
[2] 肖石红,张卫强,黄芳芳,等.CO2加富对施盐处理下银叶树幼苗叶片光合特性的影响[J]中国水土保持科学,2020,18(5):127-135
[3] 杨连新,王云霞,朱建国,等.开放式空气中CO2浓度增高(FACE)对水稻生长和发育的影响[J].生态学报,2010,30(6):1573-1585
[4] 许小勇,武佩琪,张新岭,等.外源增施CO2对茄子生长及光合特性的影响[J]北方园艺,2021(14):65-73
[5] 张云,吕镇城,徐良雄,等,LED红蓝光对植物生长发育和光合特性影响的研究进展[J]惠州学院学报,2019,39(6):33-40
[6] Amitrano C, Arena C, Pascale SD,Light and Low Relative Humidity Increase Antioxidants Content in Mung Bean (L.) Sprouts [J]. Plants, 2020,9(9):1093
[7] 王鹏,曾固,熊姜玲,等.植物工厂中光质对高品质蔬菜生产的影响研究进展[J]安徽农学通报,2021,27(23):24-29,35
[8] Berkel VN. CO2enrichment in the netherlands [J]. Acta Horticulturae, 1984,21(162):197-206
[9] 王忠,蔡恒,高煜珠,等.CO2加富对黄瓜的增产效应及其原因分析[J]江苏农学院学报,1993,15(2):37-44
[10] 魏珉,邢禹贤,马红,等.果菜苗期CO2施肥壮苗效果研究[J]山东农业大学学报(自然科学版),2000,31(2):196-200
[11] 卢涛,何兴元,陈玮.O3和CO2浓度升高对油松针叶抗氧化系统的影响[J]生态学杂志,2009,28(7):1316-1323
[12] 任锦,郭双生,杨成佳,等.紫背天葵在不同二氧化碳浓度和LED光质条件下的抗氧化应激能力研究[J]航天医学与 医学工程,2014,27(2):122-128
[13] 尚春明,董晓菲,王玉静,等.增施CO2对温室番茄产量、品质及病害的影响[J]北方农业学报,2017,45(5):33-36
[14] Muthusamy M, Jung EH, Suk HKElevated carbon dioxide significantly improves ascorbic acid content, antioxidative properties and restricted biomass production in cruciferous vegetable seedlings [J]. Plant Biotechnology Reports, 2019,13(3):293-304
[15] 王骞春.CO2浓度升高对落叶松叶片抗氧化系统的影响[J]防护林科技,2017,3(3):11-13
[16] 胡瑜辉,杨振宇,宋诗娟,等.不同光质对黄芪抗氧化酶活性及产量和品质的影响[J]山西农业科学,2021,49(10):1173-1178
[17] 蒲高斌,刘世琦,张珍.光质对番茄幼苗生长及抗氧化酶活性的影响[J].中国蔬菜,2005(9):21-23
[18] 高美芳,刘厚诚.UV-A辐射对设施作物生长及生理生化的影响[J].照明工程学报,2020,31(5):9-16
[19] 王虹,姜玉萍,师恺,等.光质对黄瓜叶片衰老与抗氧化酶系统的影响[J]中国农业科学,2010,43(3):529-534
[20] 宋阳,杜宪,王鹏,等.CO2加富对黄瓜幼苗光合特性及生长发育的影响[J]内蒙古农业大学学报(自然科学版), 2020,41(2):13-19
[21] 徐松华.逆境条件下植物体内活性氧代谢研究进展[J]安徽农学通报,2021,27(21):29-32
[22] 刘聪,董腊嫒,林建中,等.逆境胁迫下植物体内活性氧代谢及调控机理研究进展[J]生命科学研究,2019,23(3):253-258
[23] 中国科学院上海植物生理研究所,上海市植物生理学会.现代植物生理学实验指南[M].北京:科学出版社,1999
[24] 赵世杰,苍晶.植物生理学实验指导[M].北京:中国农业出版社,2015
[25] 王学奎,黄见良.植物生理生化实验原理与技术[M].北京:高等教育出版社,2015
[26] 吴强盛.植物生理学实验指导[M].北京:中国农业出版社,2018
[27] Paoletti E, Gellini R. Stomatal density variation in beech and holm oak leaves collected over the last 200 years [J]. Acta Oecologica, 1993,14:173-178
[28] 赵天宏,孙加伟,付宇,等.CO2和O3浓度升高对春小麦活性氧代谢及抗氧化酶活性的影响[J]中国农业科学,2009,42(1):64-71
[29] 山溪,秦文斌,张振超,等.低温胁迫对不同品系甘蓝幼叶ASA-GSH循环代谢的影响[J]南方农业学报,2018,49(11):2230-2235
[30] 张云婷,宋霞,叶云天,等.光质对低温胁迫下草莓叶片生理生化特性的影响[J]浙江农业学报,2016,28(5):790-796
[31] 刘福霞,刘乃森.不同光质对黄瓜幼苗生理生化特性的影响[J]湖北农业科学,2016,55(7):1722-1725
[32] 陈雄,吴冬秀,王根轩,等.CO2浓度升高对干旱胁迫下小麦光合作用和抗氧化酶活性的影响[J]应用生态学报,2000, 11(6):881-884
[33] 任红旭,陈雄,吴冬秀.CO2浓度升高对干旱胁迫下蚕豆光合作用和抗氧化能力的影响[J]作物学报,2001, 27(6):729-736
[34] 彭长连,林植芳,林桂珠.加富CO2条件下水稻叶片抗氧化能力的变化[J].作物学报,1999,25(1):39-43
[35] 庄明浩,李迎春,郭子武,等.大气CO2浓度升高对毛竹叶片膜脂过氧化和抗氧化系统的影响[J]生态学杂志,2012, 31(5):1064-1069
[36] 李德全,邹琦.抗旱性不同的冬小麦品种渗透调节能力的研究[J]山东农业大学学报(自然科学版),1991,22(4):377-383
[37] 朱维琴,吴良欢,陶勤南.干旱逆境下不同品种水稻叶片有机渗透调节物质变化研究[J]土壤通报,2003,34(1):25-28
[38] 李小玲,贺广祥,华智锐.外源渗透调节物质对黄芩幼苗抗旱能力的影响研究[J]江西农业学报,2021,33(11):20-27
[39] Högy P, Wieser H, Khler P,. Effects of elevated CO2on grain yield and quality of wheat: results from a 3‐year free‐air CO2enrichment experiment [J]. Plant Biology, 2009,11:60-69
[40] Li SH, Li YM, He XR,. Response of water balance and nitrogen assimilation in cucumber seedlings to CO2enrichment and salt stress [J]. Plant Physiology and Biochemistry, 2019,139:256-263
[41] Stitt M, Krapp A. The interaction between elevated carbon dioxide and nitrogen nutrition: the physiological and molecular background [J]. Plant, Cell & Environment, 1999,22(6):583-621
[42] Gupta M, Cuypers A, Vangronsveld J,Copper affects the enzymes of the ascorbate-glutathione cycle and its related metabolites in the roots of Phaseolus vulgaris [J]. Physiologia Plantarum, 2010,106(3):262-267
[43] Pukacka S, Ratajczak E. Antioxidative response of ascorbate-glutathione pathway enzymes and metabolites to desiccation of recalcitrant Acer saccharinum seeds [J]. Journal of Plant Physiology, 2006,163(12):1259-1266
[44] 刘宁,勾启萤,韩莹琰,等.不同氮素配比对生菜抗坏血酸—谷胱甘肽循环的影响[J]北京农学院学报,2020,35(2):59-64
[45] 魏国芹,陶吉寒,付全娟,等.硫化氢对低温胁迫下甜樱桃柱头和子房线粒体功能的影响[J].应用生态学报,2020,31(4):1121-1129
[46] Wu XX, He J, Ding HD,Modulation of zinc-induced oxidative damage in Solanum melongena by 6-benzylaminopurine involves ascorbate-glutathione cycle metabolism [J]. Environmental and Experimental Botany, 2015,116:1-11
[47] 韩敏,曹逼力,刘树森,等.低温胁迫下番茄幼苗根穗互作对其抗坏血酸—谷胱甘肽循环的影响[J].园艺学报,2019,
46(1):65-73
[48] 苗卫东,王萌,高换超,等.外源褪黑素对低温胁迫下不同葡萄品种抗氧化酶活性和ASA-GSH循环的影响[J]江苏 农业科学,2021,49(23):133-138
[49] 杨庆贺,郑成淑.低温弱光胁迫下外源ASA与CaCl_2对菊花叶片AsA-GSH循环的影响[J].山东农业大学学报(自 然科学版),2018,49(3):495-499
Effects of CO2Enrichment and LED Light on the Antioxidant Properties of Cucumber Seedlings
JIANG Wei-li1, KANG Hong-qi1, SUN Qi-ying1, ZHOU Xuan1, KONG Jing-wen2, SUN Po1, LI Qing-ming2*, LIU Bin-bin3*
1.271018,2.610299,3.271018,
A split-plot experimental design was adopted for this experiment using 'Jinyou 35' cucumber as the test material, with the main plot factor being the CO2concentration treatment, divided into two levels of atmospheric CO2concentration (≈400 μmol·mol-1) and enriched CO2concentration (≈800 μmol·mol-1). The split-plot factor was the light recipes treatment, which was divided into three light recipes: white light (W), red and blue light (RB=7:3), and red and blue light plus UV-A (RBU=7:3+20 μmol·m-2·s-1). The effect of CO2enrichment and LED light recipes on the antioxidant properties of cucumber seedlings was investigated. The experimental results showed that the CO2enrichment and RBU treatment significantly increased the activities of antioxidant enzymes such as SOD, POD, CAT, APX, GPX, GR and DHAR and the contents of osmoregulatory substances such as soluble sugars, free amino acids, soluble proteins and proline. It also increased ASA and GSH content and ASA/DHA and GSH/GSSG ratios, promoting the ASA-GSH cycle, thus reducing the content of MDA and ROS under RBU treatment and slowing down the oxidative damage caused by the addition of UV-A to cucumber seedlings. In summary, the CO2enrichment and RBU treatment can reduce the accumulation of reactive oxygen species by increasing the activity of antioxidant enzymes as well as the content of osmoregulatory substances, and improve the antioxidant properties of plants, thus contributing to the development of strong seedlings.
Cucumber; CO2enrichment; LED light recipes; antioxidant
S642.2
A
1000-2324(2022)05-0763-08
2022-01-17
2022-02-25
国家自然科学基金项目(31872154,31902092);中国农业科学院科技创新工程(34-IUA-03)
姜玮莉(1997-),女,硕士研究生,研究方向:设施蔬菜生理生态. E-mail:jiangwl97@163.com
通讯作者:Author for correspondence.E-mail:liqingming@caas.cn; lbroom@163.com
猜你喜欢
现代农业科技(2022年5期)2022-12-14
中老年保健(2021年5期)2021-08-24
当代水产(2021年6期)2021-08-13
今日农业(2020年16期)2020-12-14
智慧健康(2020年9期)2020-12-03
小学生学习指导(低年级)(2020年10期)2020-11-09
中成药(2019年12期)2020-01-04
天然产物研究与开发(2019年1期)2019-03-01
孩子(2016年7期)2016-07-11
中国粮油学报(2016年5期)2016-01-23
