
M3受体遗传变异与血压钠钾反应性的关联研究
2023-01-11 10:12胡桂霖牛泽家馨杜鸣飞张晓玉廖月圆王科科郭统帅高卫华闫睿晨罗文婧牟建军
西安交通大学学报(医学版) 2023年1期
张 玺 ,胡桂霖 ,牛泽家馨 ,杜鸣飞 ,邹 婷 ,张晓玉 ,王 兰,褚 超,廖月圆,马 琼,王 丹,王科科,贾 昊,陈 晨,严 瑜,孙 月,郭统帅,张 婕,高卫华,闫睿晨,高 可,罗文婧,牟建军,汪 洋,6
(1.西安交通大学第一附属医院心内科,陕西西安 710061;2.西北妇女儿童医院心内科,陕西西安 710003;3.西安国际医学中心医院心内科,陕西西安 710100;4.西安市人民医院心内科,陕西西安 710004;5.西安市第一医院心内科,陕西西安 710002;6.西安交通大学全球健康研究院,陕西西安 710061)
高血压是引起心脑血管疾病、慢性肾病和周围血管疾病的主要危险因素,是世界范围内最重要的单一死亡原因[1]。人体血压受复杂生理系统调节,同时受遗传和环境因素的共同作用[2]。研究表明,个体血压变化与钠盐摄入量呈正相关,高盐引起血压升高,低盐则引起血压下降,称为血压的盐敏感性[3-4]。此外,血压与膳食钾摄入量呈现明显的负相关性,且不同个体的血压反应性不尽相同[5-6]。既往研究提示,血压对钠钾摄入量反应的异质性可能取决于个体等位基因变异[7-8]。因此,识别血压钠钾敏感性的遗传变异有助于我们对血压调节机制的进一步理解。
M3毒蕈碱乙酰胆碱受体(M3受体)由CHRM3基因编码,是G 蛋白偶联受体家族的重要成员,参与第二信使信号通路传导[9-10]。M3受体广泛分布于心血管系统,在心率调节、肌力效应、心肌缺血损伤、血管舒缩及血压调节中发挥重要作用[11-13]。近来研究发现,M3受体基因变异与高血压的发生密切相关。DENG 等[14-15]发现,CHRM3基因的T1667C错义突变,可促使M3受体信号增强及再致敏,诱导肾上腺素分泌,进而致使血压升高。然而,以上研究未考虑基因和环境因素的相互作用,CHRM3基因变异引起的血压变化是否与膳食钠钾摄入量存在相关性。因此,本研究通过对既往建立的高血压队列进行慢性盐负荷及补钾干预实验,旨在讨论M3受体遗传变异与血压对膳食钠钾干预的反应性之间的关系。
1 对象与方法
1.1 研究对象
本研究通过对陕西省眉县农村地区18~60岁的成年人进行社区血压筛查,确定潜在先证者及其家系。收缩压(SBP)在130~160 mm Hg之间和/或舒张压(DBP)在85~100 mm Hg之间且未使用抗高血压药物的先证者及其兄弟姐妹、配偶和后代被招募参加本次研究。排除标准:继发性高血压,2 期高血压及以上,心血管病史或糖尿病史,使用抗高血压药物,怀孕,酗酒,正在低钠饮食或无法签署知情同意书者。最终纳入来自124个家庭的514名受试者,建立盐敏感性高血压队列并行基线调查,对其中的333名非父母者进行钠钾饮食干预研究。
1.2 饮食干预
慢性钠盐负荷及补钾干预方案参照本课题组既往研究[16-18]。首先为3 d基线调查,包括问卷调查、身高、体质量、血压、血尿生化参数等。此后依次为7 d低盐饮食(3 g氯化钠或51.3 mmol钠/日)、7 d高盐饮食(18 g氯化钠或307.8 mmol钠/日)、7 d高盐补钾饮食(高盐饮食基础上再加60 mmol/4.5 g氯化钾/日)。所有食物均为不加盐烹饪,此后由研究人员将预先分装好的钠盐及钾添加到每个受试者食物中。在整个研究期间,其他膳食营养摄入量保持不变。为了确保受试者对干预计划的依从性,其三餐(早餐、午餐和晚餐)被要求在指定餐厅进行,避免食用研究以外的食物。
1.3 24 h尿钠钾排泄量的测定
为确保受试者对饮食干预的依从性,在基线期的第3天及每个干预期的第7天收集24 h尿液标本,使用火焰光度计测定尿液中钠和钾的浓度。将所测得的钠、钾浓度乘以个体24 h尿液总容积,计算每个受试者的24 h尿钠、钾排泄量。
1.4 血压测量
受试者进行血压测量之前禁止吸烟、饮酒、进食刺激性食物及剧烈活动,静坐休息30 min后,由专业人员按照Korotkoff听音法测量右上臂坐位血压。于基线期每天和每阶段饮食干预期的第5、6、7天测量血压,每日测量3次,每次间隔1 min,最终将各阶段9次血压的平均值纳入研究。具体血压测量方法参照本课题组既往研究[16-18]。平均动脉压、脉压计算公式:平均动脉压=舒张压+1/3(收缩压-舒张压);脉压=收缩压-舒张压。将血压的钠钾反应性视为连续性变量,计算如下:低盐期血压反应性=低盐期血压-基线期血压;高盐期血压反应性=高盐期血压-低盐期血压;补钾期血压反应性=高盐补钾期血压-高盐期血压。
1.5 单核苷酸多态性(SNP)位点选择和基因分型
检索国家生物技术信息中心和基因组变异服务器数据库,按照以下标准筛选CHRM3基因的单核苷酸多态性(SNP)位点:①SNP频率分布符合Hardy-Weinberg平衡定律,P值不小于0.05;②次要等位基因频率(MAF)不小于0.05;③连锁不平衡系数r2不小于0.8。最终选取13个SNPs(rs10802764、rs997148、rs12034970、rs6429147、rs6692904、rs75804766、rs373288072、rs619288、rss58359377、rs114677844、rs663148、rs10802811、rs4072234)进行后续分析。使用Gold Mag-Mini纯化试剂盒提取受试者外周静脉血脱氧核糖核酸(DNA)。采用MassARRAY平台对CHRM3 SNPs进行基因分型。
1.6 统计学分析
连续变量以均数±标准差表示,分类变量以频率和百分比表示,连续性变量首先进行正态性检验,符合正态分布采用两独立样本t检验,不符合正态分布采用Mann-WhitneyU检验。各干预阶段指标比较采用重复测量的单因素方差分析,P<0.05为差异有统计学意义,以上分析通过软件SPSS 26.0进行。通过PLINK 软件对SNP 基因型进行Hardy-Weinberg 平衡检验和孟德尔一致性检验,同时计算MAF。运用PLINK 软件在加性模型、显性模型和隐性模型中对各SNP与血压钠钾反应性进行混合线性回归分析,同时校正年龄、性别、体质量指数和家系相关性等因素。通过R 软件(版本号:3.0.1)glmer函数,采用基于基因的分析法(gene-based analyses)评估单一性状与单个候选基因之间的关系[16]。运用截点乘积法(truncated product method,TPM)结合每个SNP关联分析的P值,评估CHRM3基因与血压钠钾反应的总体相关性[19]。
2 结果
2.1 基线特征及受试者血压的钠钾反应性
受试者基线特征及钠钾饮食干预的血压反应如表1所示。基线期先证者组的舒张压及平均动脉压在各组中最高,而收缩压低于父母组。在钠钾饮食干预过程中,各阶段血压均发生了变化,低盐期呈下降趋势,高盐期呈上升趋势,高盐补钾后的血压则再次下降。
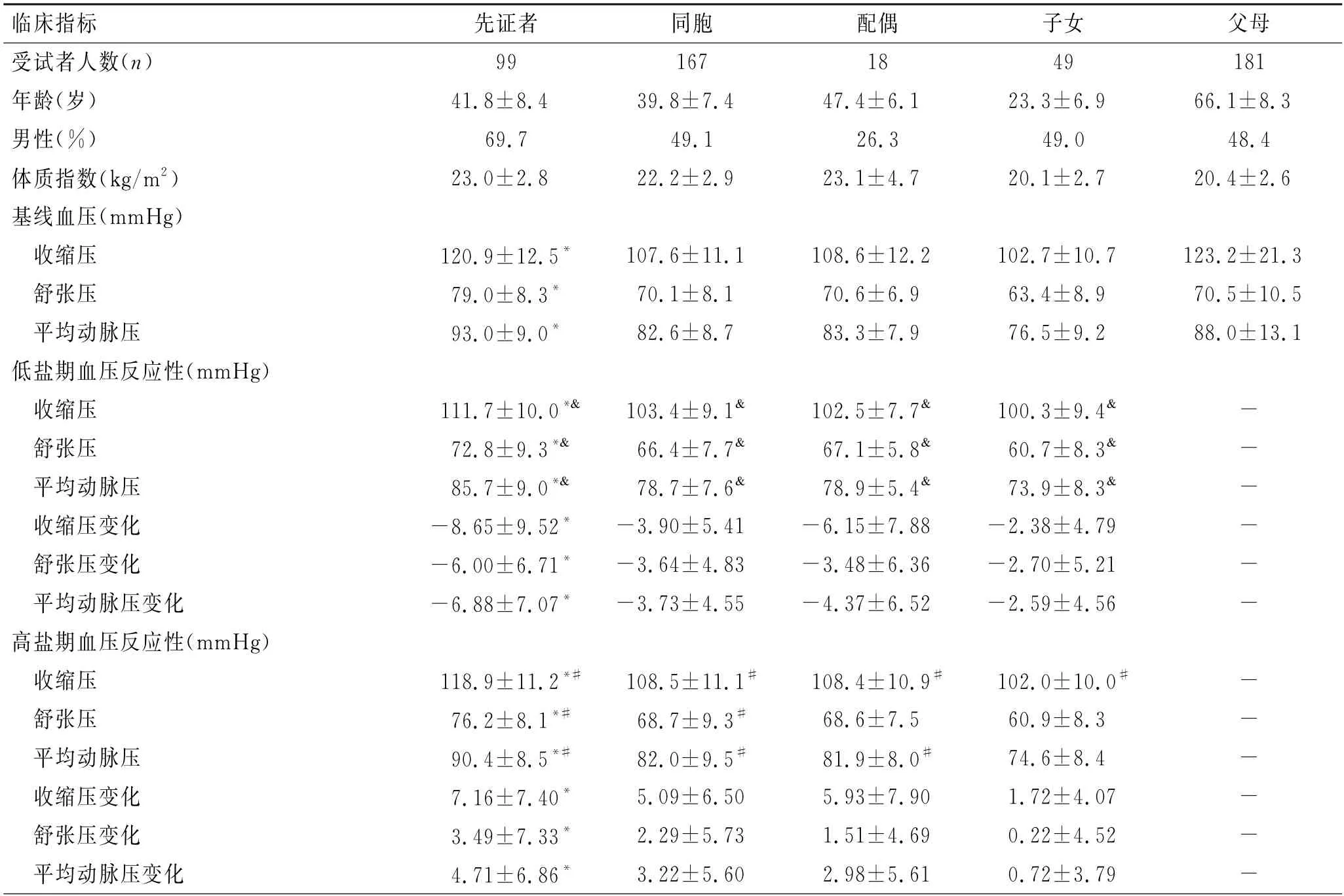
表1 基线特征及饮食干预下的血压反应性Tab.1 Baseline characteristics and BP responses to dietary intervention
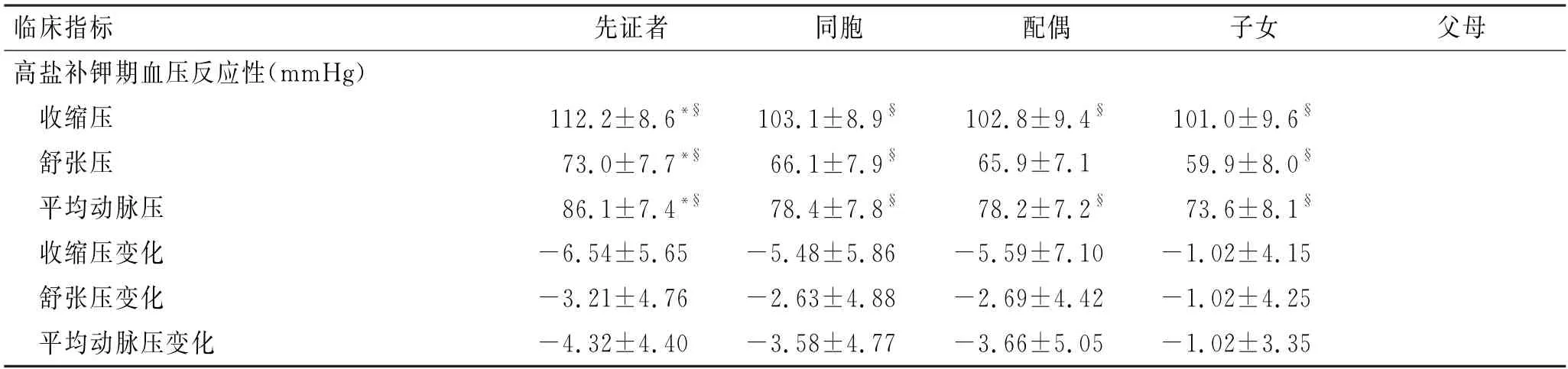
续表1
2.2 钠钾干预对受试者24 h尿钠和钾排泄的影响
各干预期受试者尿钠、钾排泄情况如表2所示,其与膳食钠钾摄入量的趋势平行。24 h尿钠排泄量在低盐期明显低于基线期,而在高盐负荷后则显著增加;此外,补钾可显著升高24 h尿钾排泄量(均P<0.05)。以上结果说明受试者对饮食干预的依从性良好。

表2 各干预期受试者尿钠、钾排泄量Tab.2 Urinary excretions of sodium and potassium in the subjects during the intervention
2.3 CHRM 3基因SNP位点信息
符合纳入排除标准的13个CHRM3基因SNP位点的基本信息如表3所示,包括基因型、位点位置、等位基因、次要等位基因频率和Hardy-Weinberg检验结果。各SNP位点基因型分布频率均符合Hardy-Weinberg平衡定律,说明受试人群具有群体代表性。
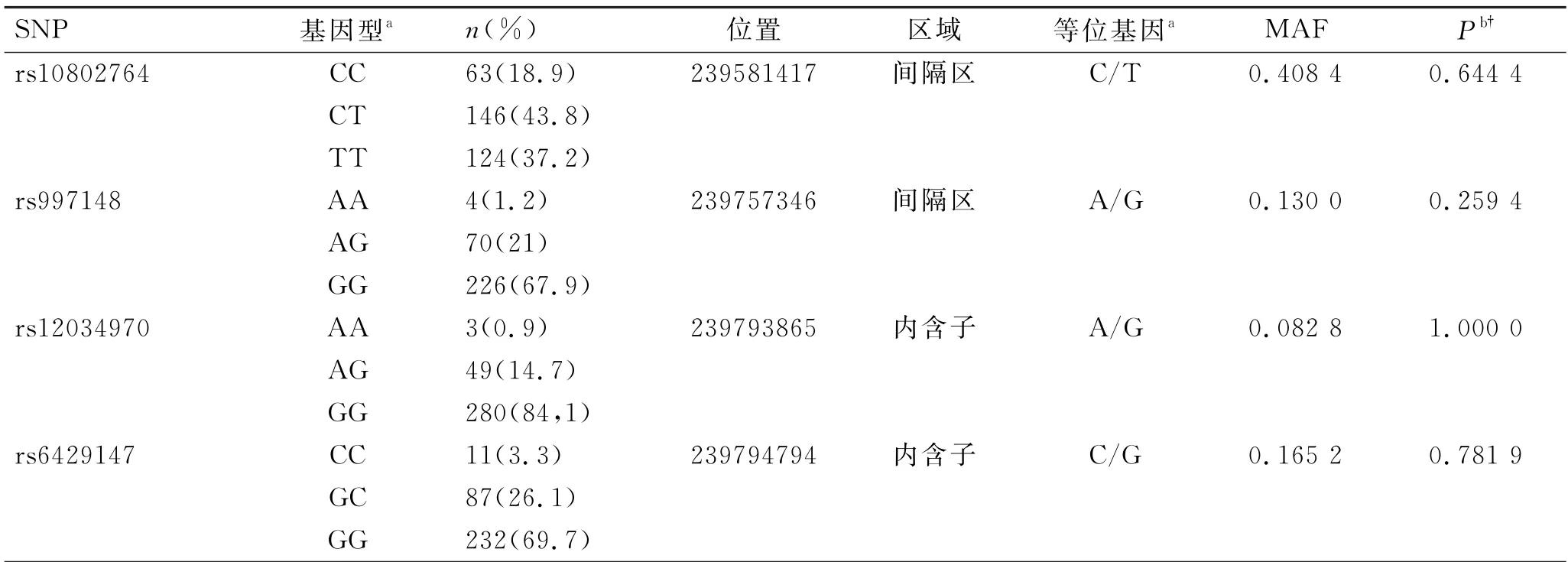
表3 CHRM 3基因SNPs基本信息Tab.3 Information on SNPs of CHRM 3

续表3
2.4 CHRM 3基因SNPs与血压钠钾反应性的关联分析
如表4所示,在低盐期,rs10802811位点与收缩压、舒张压和平均动脉压反应性显著相关,rs4072234位点与舒张压和平均动脉压反应性显著相关,rs58359377位点与平均动脉压和脉压反应性显著相关。在高盐期,rs6429147、rs114677844、rs663148位点均与收缩压和平均动脉压反应性显著相关,rs373288072位点与收缩压、舒张压和平均动脉压反应性显著相关,rs10802811与舒张压、平均动脉压和脉压反应性显著相关。在高盐补钾期,rs6692904位点与收缩压、舒张压和平均动脉压反应性显著相关。

表4 CHRM 3基因SNP位点与血压钠钾反应性的关系Tab.4 Associations of CHRM 3 SNPs with blood pressure responses to dietary intervention

续表4
进一步分析上述具有统计学意义的SNP位点,探讨其不同基因型与血压钠钾反应性的关系。如图1A、图1B所示,与野生型GG 相比,rs10802811 位点突变型AG 受试者低盐期及高盐期的舒张压及平均动脉压反应性均增高。在rs114677844位点中,突变的GG 纯合基因型受试者高盐期的收缩压反应性明显升高(图1C)。在高盐补钾期,与野生型CC 相比,rs6692904位点纯合突变型AA 受试者的舒张压及平均动脉压反应性均明显升高(图1D)。

图1 CHRM 3基因SNP位点基因型与血压钠钾反应性的关系Fig.1 Associations of CHRM 3 SNP genotypes with BP responses to dietary intervention
基于基因的分析(gene-based analyses)结果如表5所示,在高盐期,CHRM3基因与收缩压、舒张压和脉压反应性显著相关(PTMP分别为0.001、0.013、0.002)。而在低盐期及高盐补钾期,CHRM3基因与血压反应无明显相关性。

表5 CHRM 3基因与血压钠钾反应性的关系Tab.5 Associations of CHRM 3 with BP responses to dietary intervention
3 讨 论
M3毒蕈碱乙酰胆碱受体是G 蛋白偶联受体家族中五种毒蕈碱受体亚型(M1-M5)之一,广泛分布于心血管系统,在心律失常、心肌缺血、心肌肥厚、心房颤动、心力衰竭及其他非心血管疾病的发生发展中起到重要的生理功能和病理意义[11-13,20]。NISHIMARU 等[21]发现向离体小鼠心房中注入M3 选择性拮抗剂p-F-HHSiD 可显著降低左心房的心肌收缩力。ORII等[22]在兔肺动脉高压模型中发现M3 受体介导肺循环中乙酰胆碱诱导的血管舒张反应。BORELLA 等[23]发现,向大鼠侧脑室注射M3 受体拮抗剂4-DAMP可显著降低匹鲁卡品诱导的升压反应。然而,M3受体参与心血管功能调控的具体分子细胞学机制尚不清楚。M3 受体的编码基因CHRM3位于1号染色体q43,由一个外显子组成,具有>1100个SNP位点,在所有毒蕈碱受体基因中具有最丰富的遗传多态性[24-25]。在本研究中,我们发现多个CHRM3 SNPs与饮食钠钾干预的血压反应性显著相关,提示M3受体可能参与血压的调节,并与高血压的发生发展密切相关。
高血压是多基因遗传和环境因素相互作用导致正常血压调节失衡的结果,而个体遗传背景可能与盐敏感性高血压的病理生理学机制有关[26]。DENG等[27]在Dahl 盐敏感大鼠多基因高血压模型中发现,CHRM3基因末端结构域T1776C 错义突变可导致M3受体内吞延迟,引起细胞表面受体增加,功能信号延长,最终致使血压升高。为了进一步探索CHRM3基因突变在此升压过程中起到的关键作用,DENG 等[27]构建了CHRM3基因敲除DSS大鼠模型,发现CHRM3-/-组大鼠的血压明显低于CHRM3+/+组。此外,M3受体信号通路的增强可诱导肾上腺素分泌,进而影响盐敏性高血压的血管功能[14]。然而,CHRM3基因变异在人类盐敏感性高血压发展中的作用尚未被探索。本研究通过对陕西省农村人口进行慢性钠盐负荷实验,首次发现CHRM3基因SNP 位点rs10802811 与低盐期及高盐期舒张压及平均动脉压反应性显著关联,而rs6429147、rs373288072、rs114677844、rs663148 位点与高盐期收缩压及平均动脉压反应呈现显著相关性。而且,在低盐及高盐饮食干预下,rs10802811和rs114677844不同基因型之间的血压反应也存在差异。此外,通过基因分析发现,CHRM3基因与高盐期收缩压、舒张压和脉压反应的相关性显著,进一步提示M3受体遗传变异可能参与了人体血压盐敏感性的形成。
流行病学研究表明,膳食钾摄入量与血压呈负相关,且个体血压对钾摄入量的反应存在异质性[28-30]。GenSalt研究发现,APLN 和ACE2 基因SNP 位点与膳食钾补充后的血压反应显著相关[31]。此外,内皮素1(EDN1)、一氧化氮合酶3(NOS3)及E 选择素(SELE)基因多态性与补钾的降压反应均密切相关[7]。GU 等[32]提出,膳食钾补充引起的血压反应在中国人群中存在中等遗传力。本课题组既往队列研究发现抗衰老因子KLOTHO、脂肪素AdipoQ 及蛋白激酶SGK1基因多态性与补钾的血压反应性显著相关[33-35]。这些研究表明,遗传因素可能导致了血压对钾摄入量反应的变化。本研究首次讨论了M3受体基因多态性与补钾干预的血压反应性之间的关系,发现rs6692904位点与高盐补钾期的收缩压、舒张压和平均动脉压反应性显著相关,且突变型AA 基因型受试者的舒张压及平均动脉压反应性更高,提示M3受体遗传变异可能参与了血压钾敏感性的形成。
综上所述,本研究通过对具有相似遗传背景的陕西省汉族人群进行慢性盐负荷及补钾干预实验,发现CHRM3基因多态性与钠钾饮食干预的血压反应存在显著相关性,提示M3受体遗传变异参与血压钠钾敏感性的形成,为探索盐敏感性高血压的发生发展机制提供了新思路。
猜你喜欢
中国心血管杂志(2022年2期)2022-11-25
中国心血管杂志(2022年4期)2022-11-25
上海金属(2021年6期)2021-12-02
昆明医科大学学报(2021年3期)2021-07-22
中国心血管杂志(2021年6期)2021-01-02
心肺血管病杂志(2019年1期)2019-04-22
生物学通报(2019年3期)2019-02-17
中国心血管杂志(2019年3期)2019-01-04
电脑知识与技术(2018年19期)2018-11-01
中国新闻周刊(2017年16期)2017-06-15
