当归抽薹开花过程中赤霉素代谢水平及其关键酶基因克隆与表达分析
2023-01-10 06:36崔秀文黄天苗李美玲栗孟飞魏建和
中草药 2023年1期
刘 迪,崔秀文,黄天苗,李美玲,栗孟飞*,魏建和
• 药材与资源•
当归抽薹开花过程中赤霉素代谢水平及其关键酶基因克隆与表达分析
刘 迪1,崔秀文1,黄天苗1,李美玲1,栗孟飞1*,魏建和2*
1. 甘肃农业大学生命科学技术学院 干旱生境作物学国家重点实验室,甘肃 兰州 730070 2. 中国医学科学院北京协和医学院药用植物研究所,北京 100193
对当归16个赤霉素(gibberellins,GAs)进行含量测定、GAs代谢酶基因筛选并进行生物信息学分析、关键基因克隆及表达验证,以期为调控当归抽薹开花提供理论依据。利用HPLC-MS/MS对不同材料中GAs含量进行测定与分析,基于当归全长转录组筛选直接参与GAs代谢酶基因,利用在线工具进行生物信息学分析,并对关键酶基因和表达水平进行qRT-PCR检测。在所测定的16个GAs中,8个GAs含量在早薹植株中高于非早薹植株,13个GAs含量随植株发育时期延长逐渐增加。当归全长转录组中含有9个直接参与GAs代谢的酶基因,可分为6个亚家族,共含有6个保守基序,亚细胞定位于细胞质。和基因克隆片段由于插入序列与全长转录组测序片段存在长度差异。基因在抽薹植株、随种苗春化作用和植株发育时期延长呈现低表达,而在冷冻规避春化作用种苗中呈现高表达,基因则相反;和基因在叶和茎中相对于根均呈现高表达。当归含有9个直接参与GAs代谢的酶基因,各成员理化性质和结构存在一定的差异,和基因表达水平与当归抽薹开花生理调控一致。
当归;赤霉素;赤霉素代谢基因;生物信息学;基因克隆;抽薹开花
当归(Oliv.) Diels为我国大宗中药材,不仅具有补血活血、调经止痛、润肠通便等传统功效[1],还具有消炎、抗癌和治疗心脑血管疾病等现代药理学作用[2]。目前,当归年种植面积达4.35万hm2[3-4];然而,栽培生产过程中高达50%的植株早薹开花,使得肉质根木质化,不能入药[5-6]。
赤霉素(gibberellins,GAs)是一类二萜类化合物,其合成和降解在植物生长发育过程中起到重要调节作用[7-8]。目前,在已鉴定出的136种GAs中,GA1、GA3、GA4、GA5、GA6和GA7等具有生物活性[7];比如,GA4通过调控基因诱导花发育[9],GA5和GA6参与花器官分生组织分化[10]。研究发现,GAs生物合成主要分为3个阶段,在第1阶段,牻牛儿牻牛儿焦磷酸(geranylgeranyl diphosphate,GGPP)先后通过内根-古巴焦磷酸合成酶(-copalyl-diphosphate synthase,CPS)和内根-贝壳杉烯合成酶(-kaurene synthase,KS)环化为内根-贝壳杉烯(-kaurene)。在第2阶段,内根-贝壳杉烯先后通过内根-贝壳杉烯氧化酶(-kaurene oxidase,KO)和内根-贝壳杉烯酸氧化酶(-kaurenoic acid oxidase,KAO)形成GA12-7-醛(GA12-7-aldehyde)。在第3阶段,以GA12-7-醛为中心主要由2条途径合成其他GAs;在第1条途径中,GA12-7-醛首先被氧化成GA12,其在GA20ox酶的作用下依次形成GA15、GA24和GA9,在GA13ox和GA20ox酶的作用下先后形成GA53、GA44、GA19和GA20;然后,GA9和GA20在GA3ox酶的作用下分别形成具有生物学活性的GA4和GA1,继而在GA2ox酶的作用下分别形成非活性的GA34和GA8;另外,GA9在GA3ox酶的作用下也可形成GA7,GA20在GA2ox酶的作用下也可形成GA29、或在GA3ox酶的作用下形成GA5;在第2条途径中,GA12-7-醛先后在GA7ox酶的作用下形成GA14-7-醛和GA14,后者也可形成有活性的GA4[7, 11-12](图1)。以上途径表明,作为GA生物合成的限速步骤,GA2ox和GA20ox酶分别为失活和活性的关键酶,GA2ox6酶可以抑制-基因的表达延迟开花[13];GA20ox1酶可以催化GA12转变为GA9,继而促进活性GA4和GA7的生物合成促进茎的伸长[14]。
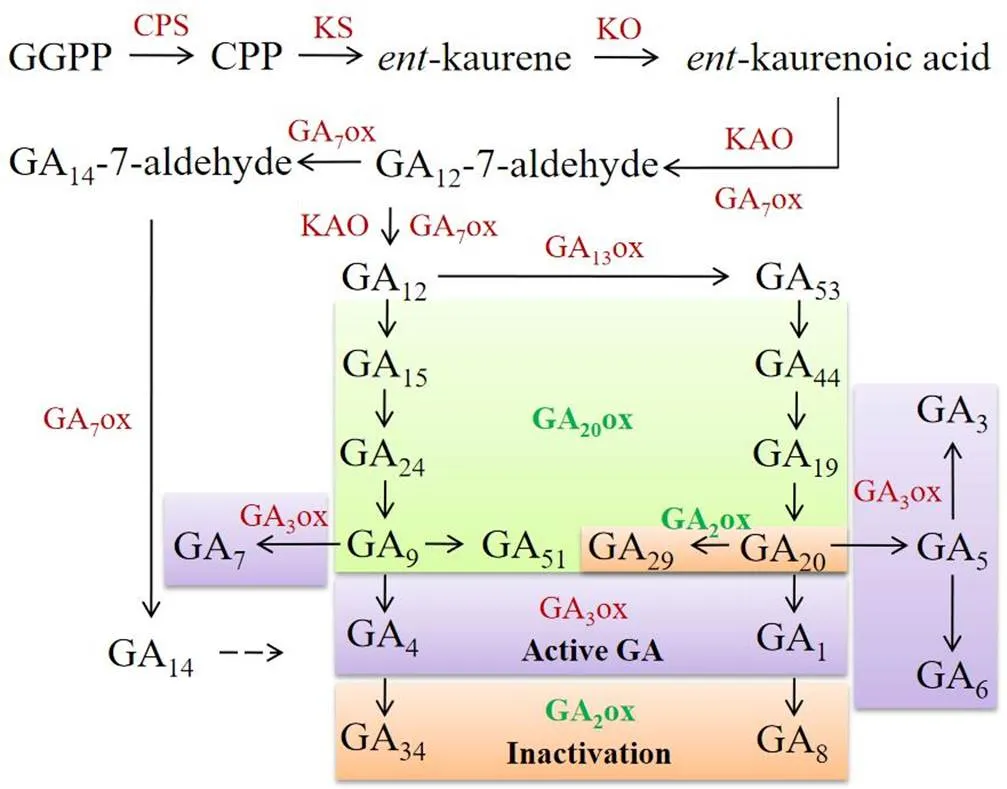
图1 GAs生物合成途径及其相关酶
前人研究发现,外源喷施GA3或抽薹开花过程中,当归植株GA3含量存在显著变化[15-16]。对当归抽薹开花分子调控机制研究发现,约70个基因通过5条途径参与抽薹开花,包括春化作用、光周期、自主、年龄、以及GA途径[17-21]。到目前为止,对当归抽薹开花过程中多个GAs代谢水平和酶基因生物信息学分析、关键酶基因克隆与表达等方面的研究鲜见报道。因此,本研究基于前期当归全长转录组测序结果,开展了16个GAs代谢水平测定、直接参与GAs代谢酶基因生物信息学分析、关键酶基因(和)克隆及表达验证,旨在深入揭示GAs代谢的生物学功能,为有效抑制抽薹开花提供理论基础。
1 材料与仪器
1.1 材料与试剂
样品取自本实验室−80 ℃保存的岷归1号两年生早薹(EB)与非早薹(Un-EB)植株、三年生不同时期[营养生长期(S1)、营养生长到生殖生长过渡期(S2)、抽薹初期(S3)和抽薹伸长期(S4)]植株的叶片和侧根,样品原植物均由甘肃农业大学栗孟飞教授鉴定为当归(Oliv.) Diels,植株生长环境及样品信息详见课题组前期实验[18-19]。GA1、GA3、GA4、GA5、GA6、GA7、GA8、GA9、GA14、GA15、GA19、GA20、GA24、GA44、GA51、GA53对照品购自Sigma公司,质量分数均大于98%,色谱甲醇购自Tedia公司。
1.2 仪器
Aglient 1290型高效液相色谱仪,美国Aglient公司;QTRAP 6500型质谱仪,美国AB公司;UYC-200型全温培养摇床,上海新苗医疗器械制造公司;氮吹仪,杭州米欧仪器有限公司;台式高速离心机,德国SORVAL公司;ABI QuantStudio 5实时荧光定量PCR仪,美国ABI公司;超微量分光光度计,上海宝予德科学仪器有限公司。
2 方法
2.1 GAs含量的测定
2.1.1 赤霉素对照品溶液制备 分别称取GA1、GA3、GA4、GA5、GA6、GA7、GA8、GA9、GA14、GA15、GA19、GA20、GA24、GA44、GA51、GA53对照品,以甲醇为溶剂配制梯度为0.1、0.2、0.5、2、5、20、50和200 ng/mL标准溶液。每个质量浓度2次重复。
2.1.2 供试品溶液制备 称取新鲜样品1.0 g,研磨后加入10 mL乙腈溶液4 ℃震荡8 h,4 ℃、13 000 r/min离心5 min取上清,沉淀再次加入10 mL乙腈溶液,重复上述操作取上清,合并2次上清液,避光,以氮气吹干有机相,400 µL甲醇溶解。4 ℃保存备用。
2.1.3 HPLC-MS/MS条件 使用高效液相色谱仪(Aglient 1290)-质谱仪(QTRAP 6500,AB SCIEX)。(1)HPLC条件:色谱柱为Poroshell 120 SB-C18反相色谱柱(150 mm×2.1 mm,2.7 µm);柱温30 ℃;进样量2 µL;流动相为甲醇/0.1%甲酸(A)-水/0.1%甲酸(B);梯度洗脱(0~1 min,20%A;1~9 min,20%~80% A;9~10 min,80% A;10~10.1 min, 80%~20% A,10.1~15 min,20% A);体积流量1 mL/min。(2)MS条件:采用电喷雾(electron spray ionization,ESI)离子源负离子电离模式,扫描方式为多反应监测(multiple reaction monitoring,MRM)模式,雾化器压力447.85 kPa,辅助气压力482.30 kPa,雾化温度400 ℃,气帘气103.35 kPa,毛细管电压4500 V。
2.1.4 标准曲线的绘制及含量测定 将“2.1.1”和“2.1.2”项中标准溶液和供试样品溶液用0.22 µm滤膜滤过,利用HPLC-MS/MS分别检测标准溶液和供试品溶液的峰面积。以色谱峰峰面积为纵坐标(),GAs质量浓度为横坐标()进行线性回归计算。根据16个GAs线性回归方程和相关系数,表明对照品在0.1~200 ng/mL内线性良好(表1)。
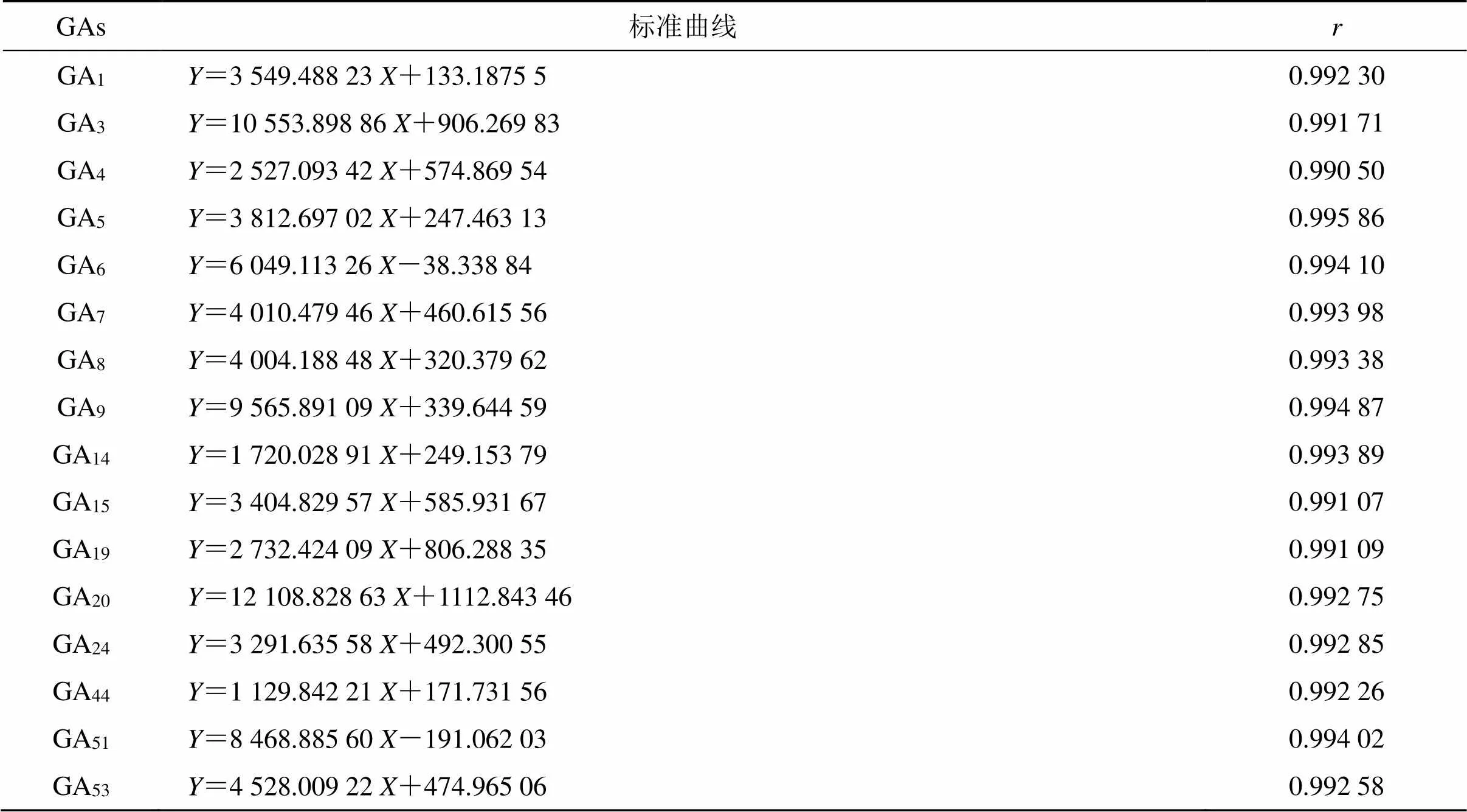
表1 16个GAs的标准曲线
按照公式计算赤霉素含量()。
=×/
为GAs的质量浓度,为样品最终溶解时所用的溶液体积,为称取的样品质量
2.1.5 统计分析 每个实验重复3次,运用SPSS软件(检验)进行数据差异显著性分析(<0.05);使用Origin 2021软件进行数据绘图。
2.2 GAs关键酶基因的生物信息学分析
2.2.1 基因序列来源 数据库来源于不同品种(岷归1号和岷归2号)全长转录组(NCBI access:PRJNA782300)[22];岷归1号种苗春化作用过程中[T1(0 ℃,14 d;未通过春化)、T2(0 ℃,60 d;通过春化)和T3(−3 ℃,125 d;低温规避春化)]全长转录组(NCBI access:PRJNA789039)[17]。利用关键词“gibberellins”和“GA”直接检索,初步筛选GA代谢相关基因,去除重复;然后利用UniProt数据库进行功能鉴定,去除非相关基因。
2.2.2 直接参与GAs代谢相关蛋白质序列鉴定及理化性质分析 蛋白质序列(包括氨基酸长度、相对分子质量和理论等电点)分析利用ExPASy在线工具;亚细胞定位利用Cell-PLoc在线工具;蛋白质二级结构预测利用PRABI-Gerland在线工具;蛋白质3D建模利用Phyre 2在线工具。
2.2.3 直接参与GAs代谢相关蛋白系统发育树构建 在NCBI数据库选择10个物种(拟南芥(L.) Heynh.黄胡萝卜Hoffm.、蓝果树Oliv.、可可L.、芝麻L.、陆地棉L.、茶树(L.) O. Kuntze、葡萄L.、烟草L、番茄L)中置信度较高的82个蛋白,利用MEGA11.0软件邻接法(neighbor-Joining,NJ)构建系统发育进化树(重复次数1000次,其他参数为默认值)。蛋白质保守基序分析利用MEME软件,并利用TBtools进行可视化。利用DNAMAN软件进行蛋白质多序列比对(深蓝色表示相似度100%、粉色>75%、浅蓝色>50%)。
2.3 当归GA2ox6和GA20ox1基因克隆
以岷归1号温室栽培植株功能叶片为材料(T2期,移栽生长40 d)[21]。总RNA提取利用Plant RNA Kit R6827试剂盒,其纯度和浓度检测使用超微量分光光度计;RNA反转录使用First-Strand cDNA Synthesis SuperMix试剂盒得到cDNA;利用NCBI Primer-BLAST设计引物(表2)。扩增产物利用1%TAE琼脂糖凝胶电泳进行检测;胶回收使用琼脂糖凝胶纯化试剂盒TIANgel Midi Purification Kit试剂盒。基因克隆使用平末端克隆试剂盒pHANDY®-Blunt Cloning Kit试剂盒;引物合成及阳性克隆测序由兰州天启基因生物科技有限公司完成。

表2 GA2ox6和GA20ox1 PCR扩增和qRT-PCR验证的引物序列
2.4 当归GA2ox6和GA20ox1基因表达分析
样品取自岷归1号两年生大田EB和Un-EB植株、三年生不同时期(S1~S4)植株、不同春化期(T1~T3)根茎顶端分生组织[17-19]、以及不同器官(根、茎和叶)。利用NCBI Primer-BLAST设计和基因qRT-PCR表达引物(表2),以作为内参基因[24]。RNA提取及反转录参照利用SuperReal PreMix Plus(SYBR Green)进行qRT-PCR 检测。利用2–∆∆Ct法计算和基因的相对表达水平[25]。
3 结果与分析
3.1 当归两年生EB和Un-EB植株中GAs含量的变化
通过对两年生EB和Un-EB植株中16个GAs进行HPLC-MS/MS测定(图2),以及含量变化进行分析(图3),结果显示,除GA1、GA4、GA7和GA24外,12个GAs含量存在显著差异。EB植株中4个GAs(GA6、GA9、GA14、GA20)含量显著高于Un-EB植株,增加1.32(GA7)~22.4(GA14)倍;其他8个GAs(GA3、GA5、GA8、GA15、GA19、GA44、GA51和GA53)含量显著降低,降低0.23 (GA44)~0.67倍(GA8)。

图2 当归EB和Un-EB植株提取液HPLC-MS/MS色谱图

*表示同一GAs在EB与Un-EB植株之间 P<0.05水平下达到显著性差异
3.2 当归三年生不同生长期植株中GAs含量的变化
通过对三年生不同生长期(S1、S2、S3和S4)植株中16个GAs进行HPLC-MS/MS测定(图4),以及含量变化进行分析(图5),结果显示,16个GAs含量也存在较大差异,在S1~S4时期,GA19含量最高(9.34 ng/g),依次为GA6(3.78 ng/g)和GA24(3.45 ng/g)。S4相对于S1,14个GAs(GA1、GA3、GA4、GA5、GA6、GA8、GA9、GA15、GA19、GA20、GA24、GA44、GA51和GA53)增加1.35(GA4)~7.18(GA9)倍,而GA14和GA7分别下降0.36和0.96倍。除GA3、GA7和GA14外,13个GAs含量随生长时间的延长呈现逐渐增加且在S4时期达到最大,而GA3、GA7和GA14分别在S2、S3和S2时期达到最大(图5)。
3.3 参与当归GAs代谢的关键酶基因生物信息学
3.3.1 当归GAs代谢相关基因鉴定及理化性质 通过对当归不同品种叶片和叶柄、种苗春化作用过程中根茎顶端分生组织的全长转录组进行筛选,发现有9个基因直接参与和18个基因间接参与GAs代谢(表3)。进一步对直接参与GAs代谢基因所编码的蛋白质序列和亚细胞定位分析,结果显示,9个蛋白质序列长度为189(GA2ox1-2)~552(GA2ox8)、相对分子质量为21 550~63 400、等电点为5.47(GA3ox2)~8.68(GA2ox8),亚细胞均定位于细胞质(表4)。
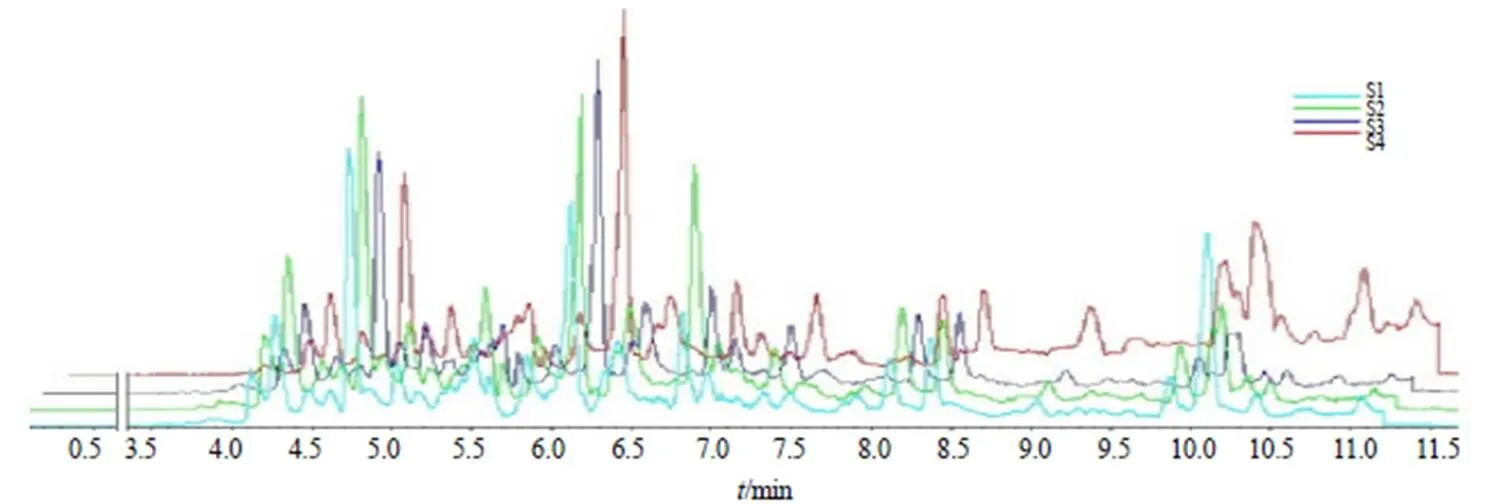
图4 当归三年生不同生长期植株提取液HPLC-MS/MS色谱图
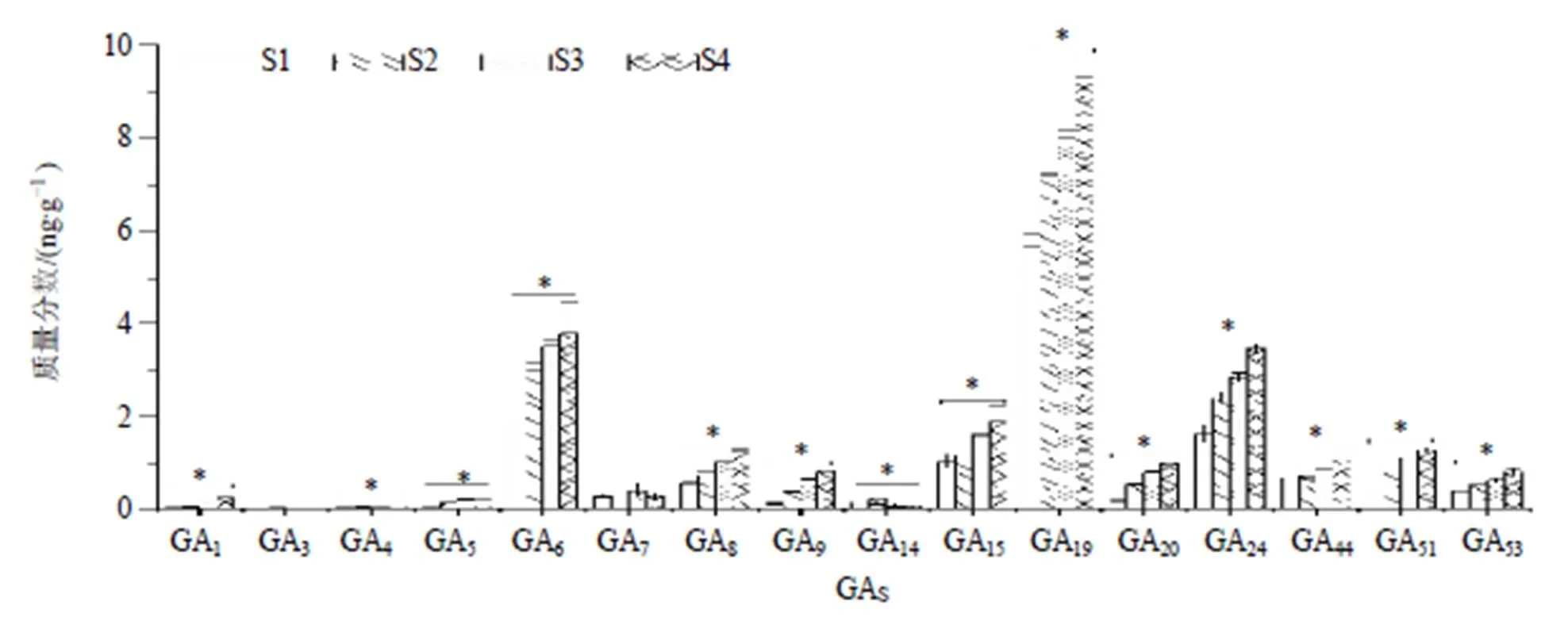
*表示同一GAs在S1和S4之间P<0.05水平下达到显著性差异
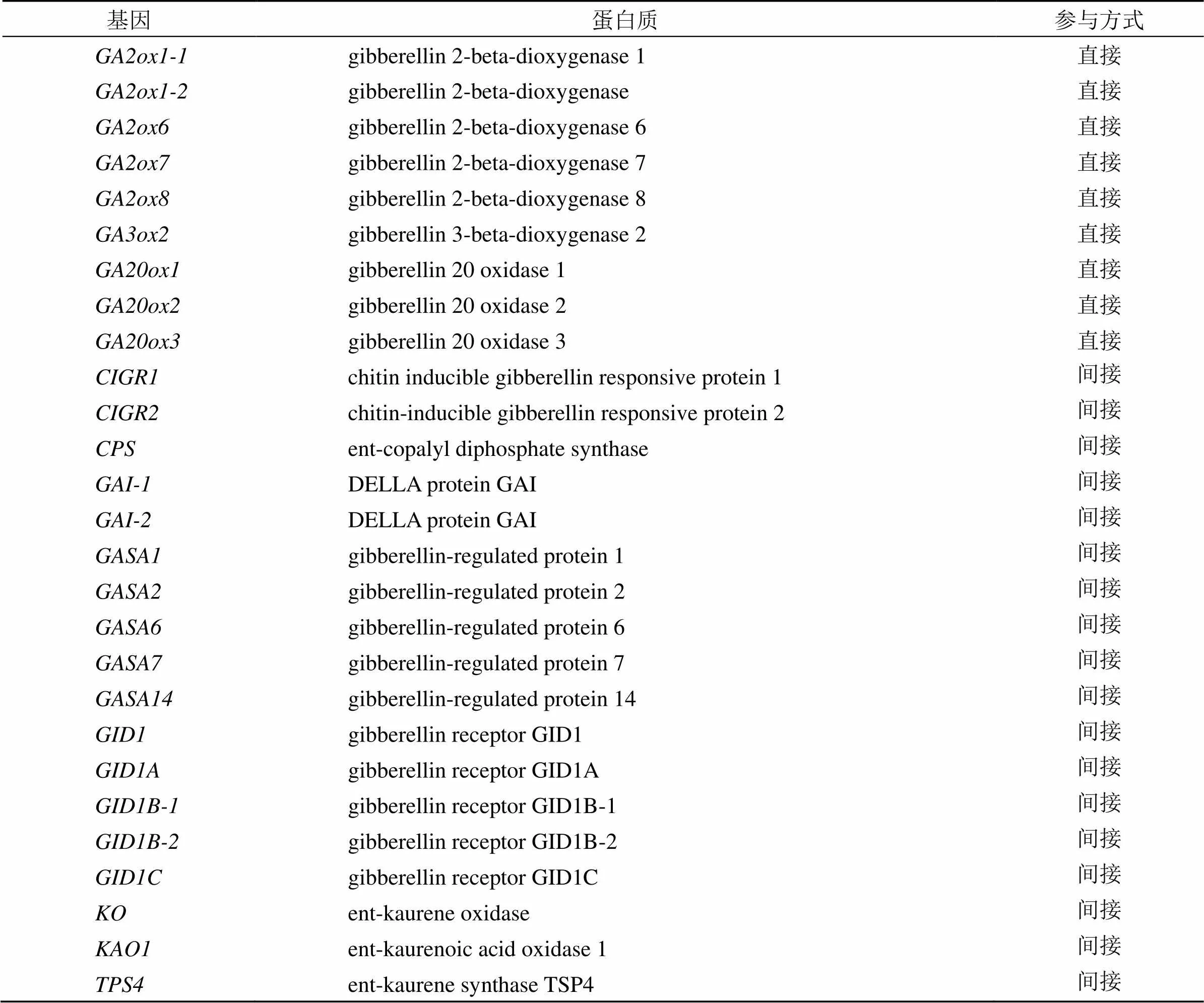
表3 当归全长转录组中直接和间接参与GAs代谢的基因

表4 直接参与当归GAs代谢蛋白质的特征及亚细胞定位
3.3.2 直接参与当归GAs代谢蛋白质的结构分析 通过对9个直接参与GAs代谢基因所编码的蛋白质进行二级结构预测以及3D建模,结果显示,蛋白质二级结构由α螺旋、延伸链、β转角和无规则卷曲组成(表5),无规则卷曲是构成二级结构的主要组成部分,占比30.25%(GA2ox8)~45.56%(GA2ox1-1),其次是α螺旋和延伸链,分别占比27.76%(GA2ox7)~41.12%(GA2ox8)和15.20%(GA3ox2)~22.22%(GA2ox1-2);有些蛋白质三级结构较为相似(如GA20ox1和GA20ox3),但有些空间结构差异较大(如GA2ox1-2、GA2ox6和GA20ox1等)(图6)。

表5 直接参与当归GAs代谢9个蛋白质的二级结构

图6 直接参与当归GAs代谢9个蛋白质的三级结构
3.3.3 直接参与当归GAs代谢蛋白质的进化树构建 通过对9个直接参与当归GAs代谢蛋白质、以及模式植物拟南芥等10个物种的82个GAs代谢蛋白质进行系统进化树构建,基于蛋白质序列相似性,这91个蛋白质被分成6个亚族Group I~VI,其中GA2ox6和GA20ox1分别位于GroupI和GroupIII亚族(图7)。
3.3.4 直接参与当归GAs代谢的蛋白质基序 通过对以上91个蛋白质进行结构域和基序分析,结果发现,这91个蛋白质分为6个亚族,与系统进化树关系一致(图7),序列含有6个保守基序(表6);直接参与当归GAs代谢的同一亚家族蛋白质基序具有高度相似性,其中,GA2ox6亚族均含有Motif 1~Motif 4和Motif 6;GA20ox1亚族均含有Motif 1~Motif 6;而不同亚族蛋白质基序存在较大差异,比如,Motif 5只存在于Group III亚族(图8)。
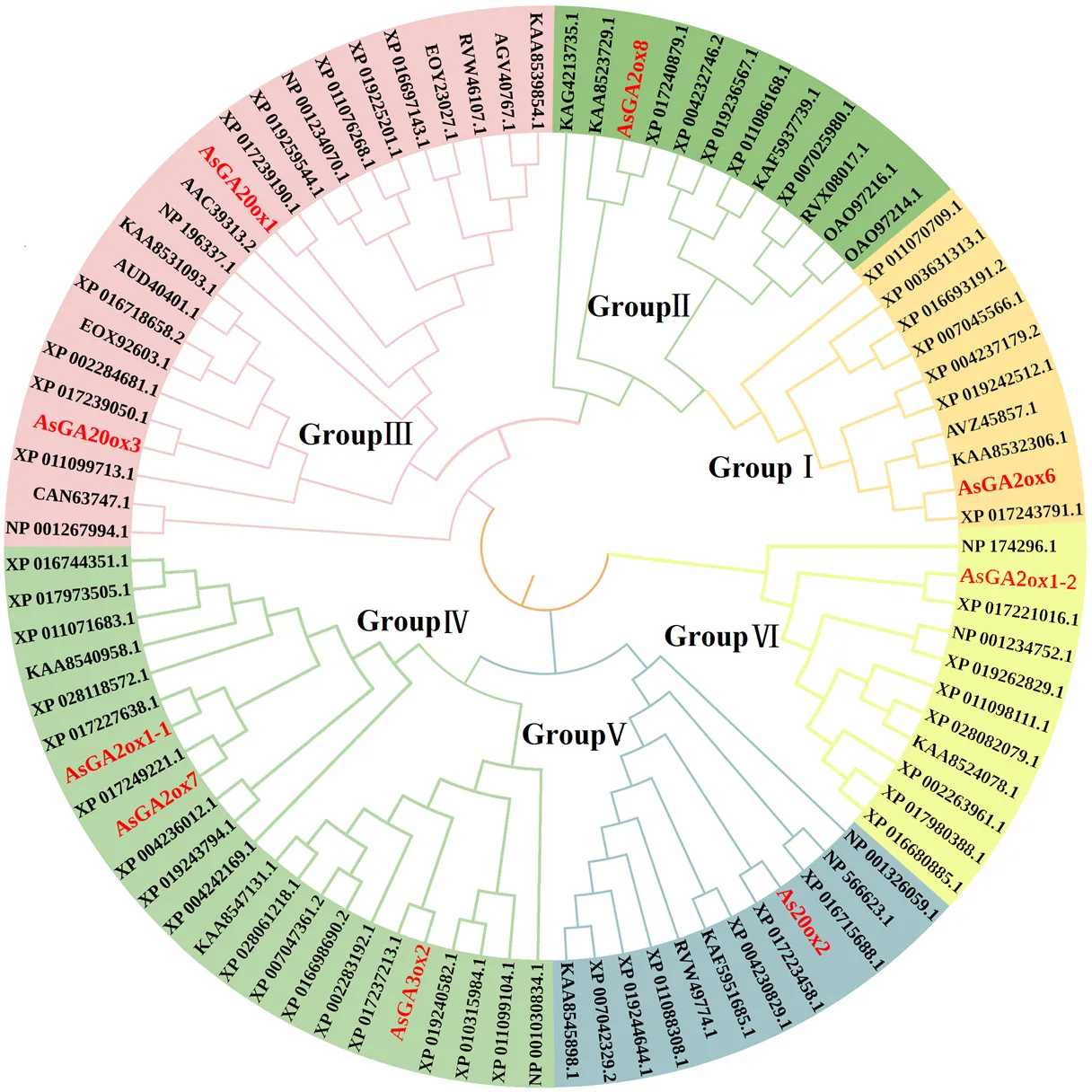
图7 当归和拟南芥等11个物种GAs代谢蛋白质的系统进化树

表6 直接参与GAs代谢蛋白质6个保守基序及其序列
3.3.5 当归GA2ox6和GA20ox1蛋白质多序列比对 通过将当归AsGA2ox6和AsGA20ox1与NCBI中拟南芥和黄胡萝卜等物种中同源性较高的5个蛋白质进行多序列比对,结果显示,GA2ox6与同科黄胡萝卜同源性最高,相似性达到89.74%(XP_017243791.1),依次为蓝果树71.55%(KAA8532306.1)、茶树68.84%(AVZ45857.1)、烟草66.57%(XP_019242512.1)和番茄65.98% (XP_004237179.2)(图9-A);GA20ox1也与黄胡萝卜同源性最高,达到82.95%(XP_017239190.1),依次为番茄69.13%(NP_001234070.1)、芝麻67.94%(XP_011076268.1)、蓝果树67.09%(KAA8539854.1)和烟草66.33%(XP_019225201.1)(图9-B)。

图8 当归和拟南芥等11个物种GAs代谢蛋白质的保守基序
3.4 当归GA2ox6和GA20ox1基因克隆
为了保证全长转录组中当归和基因碱基序列的准确性,基于全长转录组中和基因碱基序列设计扩增引物,以岷归1号功能叶片中RNA反转录所得的cDNA为模板,进行和基因克隆。琼脂糖凝胶电泳显示,和基因扩增片段大小为1000~2000 bp(图10);产物回收与碱基测序显示,基因克隆长度为1477 bp(458 bp和777 bp处插入了95 bp和370 bp);基因克隆长度为1455 bp(558 bp和880 bp处插入了108 bp和199 bp);通过与全长转录组测序获得的和进行序列比对,除了插入2个片段外,其他相似度为100%(图11)。

图9 当归GA2ox6(A)和GA20ox1(B)蛋白质与5个其他物种蛋白质的多序列比对

图10 当归GA2ox6和GA20ox1基因扩增产物
3.5 当归GA2ox6和GA20ox1基因表达分析
为了进一步验证和基因的生物学功能,对当归不同材料中和基因的表达水平进行了qRT-PCR检测与分析,结果(图12)显示,EB相对于Un-EB,呈现低表达,而呈现高表达;不同生长期植株中,表达水平随着时间延长呈现逐渐降低,而呈现逐渐增加;不同春化期种苗根茎顶端分生组织中,T2相对T1,表达水平降低,而T3相对T1表达增加,而则相反;不同器官中,和在茎和叶相对于根均呈现高表达。

图11 当归GA2ox6(A)和GA20ox1 (B) 基因克隆与全长转录组测序的序列比对

图12 当归GA2ox6和GA20ox1基因在不同材料中的相对表达水平
4 讨论
目前,EB开花导致根木质化不能入药仍是困扰当归生产、质量和效益提升的重大难题[4]。大量研究表明,GA调控植物生长发育过程——种子萌发、茎的伸长、花的发育、果实的形成和种子的发育,其含量的变化受到特定环境因素和生物合成酶基因的影响[7]。本研究发现,16个GAs含量在当归EB相对于Un-EB植株、不同生长发育时期植株中存在显著差异;基于当归全长转录组,发现9个基因直接参与GAs代谢;另外,克隆获得的关键酶基因和表达水平与相应的GAs代谢水平一致。
大量研究表明,GA最显著的作用是通过促进细胞伸长和分裂进而诱发植物茎的伸长[7]。前期研究发现,外源喷施GA3可显著促进当归提前抽薹开花[15];在抽薹分化前期植株中含量较高[16];在种苗春化作用过程中显著升高,而冷冻回避春化作用显著降低[17]。本研究发现,GA3含量在EB植株低于Un-EB植株,生长发育过程中S1~S4时期呈“上升-下降-上升”趋势,这与前人的研究结果存在一定差异。可能原因是:S1~S2时期植株处于生长旺盛阶段,植株营养物质积累有利于GA3的生物合成,进而诱发抽薹,当植株开始抽薹,较多的GA3转运至茎,促进花器官形成和果实发育[26]。另外,研究证实在菠菜中喷施赤霉素抑制剂BX-112,导致了GA1和GA8水平降低从而积累了GA20,抑制菠菜茎的伸长。这与本研究结果一致,在当归发育过程中GA1、GA4和GA20逐渐升高。较高的GA1通过促进细胞分裂来增加顶端分生组织的生命活力使节间数增多,同时促进微管组织和叶片发育,从而加速植物生长速率促进抽薹[27]。然而,GA4在调控抽薹开花过程中,集中在花器官中,调控雄蕊的发育等,因此叶片和侧根GA4含量可能低于其它GAs[9]。此外,活性GAs(如GA1、GA4、GA6和GA7)在EB植株中显著高于Un-EB植株,而非活性GAs相反;在不同生长期16个GAs中,除GA3、GA7和GA14外,13个GAs随生长发育时间延长含量逐渐增加。
为了进一步了解GAs参与当归抽薹开花的机制,本研究对9个直接参与GAs代谢基因进行了生物信息学分析和关键基因克隆及表达验证。研究发现,拟南芥含有16个GA氧化酶基因(和)[28];苹果含有41个GA氧化酶基因(,和)[29];葡萄含有24个GA氧化酶基因(、和)[30]。本研究基于当归全长转录组,发现有9个GA氧化酶基因(、、、和-3)。本课题组获得的9个GA氧化酶与拟南芥、苹果和葡萄等[28-30]氧化酶二级结构基本一致;苹果、葡萄和黄瓜等[29-31]其他物种亚细胞定位于细胞核或细胞质,本研究中的有9个GA氧化酶均定位于细胞质。
通过对拟南芥、苹果和葡萄等[28-30]物种研究发现,GA2ox、GA3ox和GA20ox基因家族根据结构的差异分布在不同的亚族中,表明植物GA氧化酶基因进化关系相近的保守结构相似,同一亚族在进化上具有一定的保守性,且进化关系越近的亚族,其保守基序的同源性越高。本研究基于系统发育树发现,GA20ox1和GA20ox3位于GroupⅢ亚族,而GA2ox1-1、GA2ox7和GA3ox2位于GroupⅣ亚族,其中,GA20ox1和GA20ox3含有相同的保守基序(Motif 1~6),而GA2ox1-1、GA2ox7和GA3ox2含有相同的保守基序(Motif 1~3,Motif 6)。同时,本研究中当归GA2ox6和GA20ox1蛋白质序列与同科植物胡萝卜的相似度最高,亲缘关系最近。另外,基因克隆获得的和序列与转录组结果存在一定差异,表明本研究所获得的序列可能尚不完整。
在当归EB相对Un-EB植株研究中发现,基因表达量较高,而基因表达量没有显著变化[21];本研究发现,基因在EB植株中低表达,这与GA8在EB植株中含量低于Un-EB植株的结果一致;基因在EB植株中高表达,这与GA1、GA4、GA9和GA20在EB植株中含量低于高于Un-EB植株的结果一致。在植物不同器官研究中发现,甘蓝型油菜中基因主要在根和叶中表达[32],拟南芥中基因在叶中表达量显著高于根[33],枳橙中主要在节间、叶片和种子中表达[34],黄瓜中基因在叶中表达量显著高于根和茎[31],这与本研究中当归和基因表达量在叶>茎>根的结果一致。同时,在本研究不同生长期发现,基因表达量随植株生长发育时期延长和在种苗春化作用中显著降低,在冷冻回避春化作用种苗中显著升高,而基因表达量相反。大量研究表明,GA2ox6通过催化GA20、GA4和GA1分别形成非活性的GA29、GA34和GA8调节植株抽薹开花[32];因此,本研究不同材料中参与GAs酶基因表达量的变化,会直接引起相应GAs含量水平的变化,最终呈现出抽薹开花的生理差异。
综合以上研究表明,本实验首次对当归抽薹开花过程中16个GAs含量进行了测定、对筛选的9个直接参与GAs代谢的酶基因进行了生物信息学分析、并对关键酶基因(和)进行了克隆及表达验证。但对关键酶基因(和)片段的完整性、以及其它GAs生物合成相关基因(如、和)的生物学特性等还需要进一步研究。
利益冲突 所有作者均声明不存在利益冲突
[1] 中国药典 [S]. 一部. 2020: 139.
[2] Wei W L, Zeng R, Gu C M,.in China-A review of botanical profile, ethnopharmacology, phytochemistry and chemical analysis [J]., 2016, 190: 116-141.
[3] 晋玲. 当归生产加工适宜技术[M]. 北京: 中国医药科技出版社, 2018: 236.
[4] 栗孟飞, 康天兰, 晋玲, 等. 当归抽薹开花及其调控途径研究进展 [J]. 中草药, 2020, 51(22): 5894-5899.
[5] Zhang H Y, Bi W G, Yu Y,.(oliv.) Diels in China: Distribution, cultivation, utilization and variation [J]., 2012, 59(4): 607-613.
[6] Li M L, Cui X W, Jin L,. Bolting reduces ferulic acid and flavonoid biosynthesis and induces root lignification in[J]., 2022, 170: 171-179.
[7] Lincoln Taiz, Eduardo Zeiger. 植物生理学 [M]. 北京: 科学出版社, 2015: 569-596.
[8] Wilson R N, Heckman J W, Somerville C R. Gibberellin is required for flowering inunder short days [J]., 1992, 100(1): 403-408.
[9] Eriksson S, Böhlenius H, Moritz T,. GA4 is the active gibberellin in the regulation of LEAFY transcription andfloral initiation [J]., 2006, 18(9): 2172-2181.
[10] King R W, Evans L T. Gibberellins and flowering of grasses and cereals: Prizing open the lid of the “florigen” black box [J]., 2003, 54: 307-328.
[11] Ait-Ali T, Frances S, Weller J L,. Regulation of gibberellin 20-oxidase and gibberellin 3beta-hydroxylase transcript accumulation during De-etiolation of pea seedlings [J]., 1999, 121(3): 783-791.
[12] Fleet C M, Yamaguchi S, Hanada A,. Overexpression of AtCPS and AtKS inconfers increased ent-kaurene production but no increase in bioactive gibberellins [J]., 2003, 132(2): 830-839.
[13] Yan J D, Liao X Y, He R Q,. Ectopic expression of GA 2-oxidase 6 from rapeseed (L.) causes dwarfism, late flowering and enhanced chlorophyll accumulation in[J]., 2017, 111: 10-19.
[14] Schomburg F M, Bizzell C M, Lee D J,. Overexpression of a novel class of gibberellin 2-oxidases decreases gibberellin levels and creates dwarf plants [J]., 2003, 15(1): 151-163.
[15] 张珍芳. 当归早期抽薹相关激素及其它生理生化指标分析 [D]. 兰州: 兰州大学, 2011.
[16] 黄珊. 当归早期抽薹分化和内源性激素变化研究 [D]. 杨凌: 西北农林科技大学, 2020.
[17] Luo M M, Liu X X, Su H Y,. Regulatory networks of flowering genes induring vernalization [J].(), 2022, 11(10): 1355.
[18] Li M F, Li J, Wei J H,. Transcriptional controls for early bolting and flowering in[J].(), 2021, 10(9): 1931.
[19] Li J, Li M L, Zhu T T,. Integrated transcriptomics and metabolites at different growth stages reveals the regulation mechanism of bolting and flowering of[J].(), 2021, 23(4): 574-582.
[20] Gao X, Guo F X, Chen Y,. Full-length transcriptome analysis provides new insights into the early bolting occurrence in medicinal[J]., 2021, 11(1): 13000.
[21] Yu G, Zhou Y, Yu J J,. Transcriptome and digital gene expression analysis unravels the novel mechanism of early flowering in[J]., 2019, 9(1): 10035.
[22] Zhu T T, Zhang M H, Su H Y,. Integrated metabolomic and transcriptomic analysis reveals differential mechanism of flavonoid biosynthesis in two cultivars of[J]., 2022, 27(1): 306.
[23] Liu X X, Luo M M, Li M F,. Depicting precise temperature and duration of vernalization and inhibiting early bolting and flowering ofby freezing storage [J]., 2022, 13: 853444.
[24] Xu R, Xu J, Li Y C,. Integrated chemical and transcriptomic analyses unveils synthetic characteristics of different medicinal root parts of[J]., 2020, 12(1): 19-28.
[25] Willems E, Leyns L, Vandesompele J. Standardization of real-timegene expression data from independent biological replicates [J]., 2008, 379(1): 127-129.
[26] 周琴, 张思思, 包满珠, 等. 高等植物成花诱导的分子机理研究进展 [J]. 分子植物育种, 2018, 16(11): 3681-3692.
[27] Zeevaart J A, Gage D A, Talon M. Gibberellin A1 is required for stem elongation in spinach [J]., 1993, 90(15): 7401-7405.
[28] 石永春, 邓晓旭, 刘卫群. 拟南芥GA20氧化酶家族的生物信息学分析 [J]. 河南农业大学学报, 2010, 44(4): 453-455, 461.
[29] 董凤, 樊胜, 马小龙, 等. 苹果赤霉素氧化酶基因GA2ox、GA3ox和GA20ox家族全基因组鉴定及表达分析 [J]. 园艺学报, 2018, 45(4): 613-626.
[30] He H H, Liang G P, Lu S X,. Genome-wide identification and expression analysis of GA2ox, GA3ox, and GA20ox are related to gibberellin oxidase genes in grape (L.) [J]., 2019, 10(9): 680.
[31] 庞保亚, 李强, 任仲海. 黄瓜CsGA20ox1异源表达促进拟南芥植株发育 [J]. 山东农业大学学报: 自然科学版, 2018, 49(4): 578-584.
[32] 廖晓英. 甘蓝型油菜中GA2ox家族基因克隆及BnGA2ox6功能研究 [D]. 长沙: 湖南大学, 2015.
[33] 段秋红, 赵小英, 贺热情, 等. 拟南芥GA2ox基因家族启动子分析 [J]. 生物信息学, 2013, 11(4): 275-281.
[34] 袁飞荣, 李芳, 蒋巧巧, 等. 转基因枳橙中GA20ox1与rol基因互作关系的研究 [J]. 中国农学通报, 2011, 27(19): 214-221.
Analysis on gibberellins metabolic level, key enzyme genes clone and expression pattern during bolting and flowering plants in
LIU Di1, CUI Xiu-wen1, HUANG Tian-miao1, LI Mei-ling1, LI Meng-fei1, WEI Jian-he2
1. College of Life Science and Technology/Key Laboratory of Aridland Crop Science, Gansu Agricultural University, Lanzhou 730070, China 2. Institute of Medicinal Plant Development, Chinese Academy of Medical Sciences & Peking Union Medical College, Beijing 100193, China
To determine the contents of 16 gibberellins (GAs), screen the genes involved in GAs metabolism and analyze their bioinformatics, as well as clone the key genes and validate their expression level, so as to provide theoretical basis for regulating bolting and flowering of.The GAs contents in different materials were determined by HPLC-MS/MS. The genes directly involved in GAs metabolism were screened based on the full-length transcriptomes of, the bioinformatics was analyzed using the online tools, and the expression levels of theandgenes were validated by qRT-PCR.Among of the 16 GAs, the contents of eight GAs were higher in bolted plants than unbolted plants, and the contents of 13 GAs were increased with the extension of plant development. A total of nine genes directly involved in GAs metabolism were identified from, with six subfamily classified, six motifs included, and the sub-cellular located in the cytoplasm. There was a certain difference in the base sequences between the cloned fragment ofandwith the full-length transcriptomes, due to the inserted fragments in the sequence ofand. The expression levels ofgene were down-regulated in the bolted plants compared to unbolted plants, and with the extension of seedlings vernalization and plants development, while up-regulated in the seedlings with frozen-avoided vernalization; but thegene showed opposite results; and the expression levels ofandgenes in leaves and stems were higher than roots.There are nine genes directly involved in GAs metabolism in, and certain differences in physical and chemical properties and structure among the nine members; the expression levels of theandgenes are in accordance with the physiological regulation of bolting and flowering of.
(Oliv.) Diels; gibberellins; gibberellins metabolic gene; bioinformatics; gene clone; bolting and flowering
R286.12
A
0253 - 2670(2023)01 - 0222 - 13
10.7501/j.issn.0253-2670.2023.01.024
2022-05-06
国家自然科学基金项目(32160083);干旱生境作物学国家重点实验室(甘肃农业大学)基金项目(GSCS-2021-Z03);道地药材生态种植与质量保障项目(202103003);财政部和农业农村部:国家现代农业产业技术体系(CARS-21)
刘 迪(1997—),女,山西运城人,硕士研究生,主要从事当归抽薹开花调控方面研究。E-mail: liudi9728@163.com
通信作者:栗孟飞(1980—),男,河南驻马店人,博士,教授,主要从事药用植物学研究。E-mail: lmf@gsau.edu.cn
魏建和(1970—),男,福建建阳人,博士,教授,主要从事药用植物栽培、分子育种和次生代谢产物调控研究。E-mail: jhwei@implad.ac.cn
[责任编辑 时圣明]
猜你喜欢
蔬菜(2022年1期)2022-11-11
麦类作物学报(2022年4期)2022-05-23
南方医科大学学报(2022年3期)2022-04-13
浙江大学学报(农业与生命科学版)(2021年3期)2021-07-10
中国预防兽医学报(2020年8期)2020-11-05
三农资讯半月报(2020年15期)2020-08-25
支部建设(2019年24期)2019-09-28
金桥(2019年12期)2019-08-13
文苑(2019年14期)2019-08-09
江苏农业科学(2018年11期)2018-06-29