
有机脂类分子重建海洋古温度研究进展
2023-01-10 07:34杨迎雪李丽郁金勇贺娟贾国东
海洋地质与第四纪地质 2022年6期
杨迎雪,李丽,郁金勇,贺娟,贾国东
同济大学海洋与地球科学学院海洋地质国家重点实验室,上海 200092
20世纪以来全球气温持续变暖,最近几年暖冬、酷暑、冷夏等自然灾害频繁发生,气候异常给人类带来了许多灾难性的后果,全球气候变化已成为关系人类生存发展的重大问题。未来气候发展如何,不仅为科学界所关注,而且受到了各国政府及社会公众的普遍关注,而目前的气候模型预测结果非常发散,究其原因是对过去气候变化的认识和了解不够充分。温度是气候变化中一个敏感和关键的因子,是气候模拟试验中一个重要的边界条件,也是古气候、古海洋学研究中一个重要的基本参数,如何获得可靠的古温度记录一直备受科学界关注。
古温度重建为我们了解过去的气候变化提供了宝贵信息,多年来陆地和海洋古温度重建工作取得了很大的进展,所用工具也从早期基于微体古生物属种形态统计的转换函数法、现代类比法等发展到无机元素比值法和同位素指标法(浮游有孔虫的δ18O和Mg/Ca比值)[1-2]。但基于无机指标的古温度重建存在一定的不确定性,以有孔虫为例,存在生物栖息深度、外壳保存状况、海水组成变化等影响因素[3]。相对而言,有机脂质生物标志物来源于活的生物体,对外部环境变化反应灵敏且具有相对较高的保存潜力,在地质演化过程中具有一定的稳定性,具有分析精度高、分析速度快、人为干扰少、所需样品量少等优点。尤其在深海或湖泊等化石记录有缺失的环境中,它们对恢复古生态环境有重要作用[4]。因此,有机温标已被广泛应用于海洋、湖泊及土壤的古温度重建。
海洋约占地球表面积的70%,在全球气候和环境演变过程中发挥着重要作用,而且海洋沉积物具有沉积连续、干扰因素少的特点,是古气候研究的重要载体。随着高效液相色谱-质谱联用技术(HPLC-MS)和气相色谱-质谱联用技术(GC-MS)等现代分析测试方法的高速发展,新的有机脂质温度指标不断被开发出来,开展现有的海洋有机脂类温度指标的系统总结与评述,对今后的古气候研究具有借鉴意义。因此,本文按照指标建立的时间顺序总结了6种古海洋温度有机代用指标的研究概况,包括长链烯酮古温度指标、四醚膜脂古温度指标TEX86、梯烷脂古温度指标NL5、长链烷基二醇古温度指标LDI、羟基四醚膜脂古温度指标RIOH和3-羟基脂肪酸古温度指标RAN13,讨论了它们在古温度重建中的潜力,以及不同指标重建温度存在差异的可能原因,这是由于它们有各自独特的生物来源,重建结果潜在地反映了生物源的生长环境温度,存在季节性和深度差异,同时非生物因素也不同程度地影响了重建结果,进而说明多指标对比可以减小误差,提高结果的准确性,最后,提出了未来研究的展望。
1 长链烯酮古温度指标
长链烯酮(Long-chain alkenones, LCAs)是碳链长度为35——42,且具有2——5个不饱和双键的甲基(Me)和乙基(Et)酮的脂肪族化合物,均呈反式构型[5],分子结构如图所示(图1a)[6],通过气相色谱(GC)和气相色谱-质谱(GC-MS)技术可以很容易地在亚纳克量级上被识别出来。Boon等[7]首次报道在非洲西南沃尔维斯湾海岸的硅藻泥中检测到该类物质,随后发现其广泛分布于全球的淡水与海洋环境中[8]。由于其对环境因子变化较敏感,且能够在沉积物中长久保存[9],所以可用于重建古温度、古生产力、古大气CO2分压,是古环境研究的重要工具。

图1 长链烯酮分子结构(a),不同温度下E.huxleyi培养实验中长链烯酮的代表性色谱图(b)[6,14]Fig.1 Structures of long-chain alkenones (a) and representative chromatograms showing distribution of long-chain alkenones in cultures of E.huxleyi grown at different temperatures(b)[6,14]
长链烯酮来源于定鞭藻(Haptophyte)。根据18S rRNA的基因序列分析[10],产长链烯酮的定鞭藻可分为3组(Groups I——III),每一组适宜的生存环境不同。I组包括未能分离培养的定鞭藻,它们主要出现在高纬度淡水和低盐湖泊中[11-12]。II组物种最具普遍性,在低盐、高盐湖泊和沿海地区都有发现,包括等鞭金藻Chrystila lamellosa和Isochrysis galbana[13]。III组主要生存于海洋环境的上层水体,包括颗石藻Emiliania huxleyi和Gephyrocapsa oceanica,它们是现代海洋长链烯酮的主要生产者[10]。
Brassell等[15]于1986年分析了不同纬度第四纪海洋沉积物样品中长链烯酮的分布特征,发现其不饱和度与温度密切相关,具体表现为不饱和度随温度的降低而增加(图1b)[14],基于此提出了烯酮不饱和度指标(U是不饱和度,K是烯酮,37是碳数;表1)。随后,Prahl和Wakeham[6]的研究发现,当温度高于12℃时,含有4个双键的C37:4型烯酮含量极低,在中低纬地区通常不存在或无法检测到,因此剔除C37:4烯酮,提出了适用于中低纬区域的优化指标并通过培养实验得到了校准公式(表1-4)。随后,Prahl等[16]增加了两组培养数据,又得到了略有不同的校正公式(表1-5)。此后Müller等[17]通过对大西洋、印度洋和太平洋的370个表层沉积物中长链烯酮分布进行研究(60°S-60°N),建立了的全球海洋表层温度(sea surface temperature,SST)的校准公式(表1-6),证明了古温度指标的普遍适用性。而对于极地高纬地区,Bendle和Rosell-Melé[18]提出了适用于估算样品中C37:4值过高时古温度重建的替代指标(表1)。
表1 长链烯酮古温度指标()及温度校准公式Table 1 Paleotemperature index (derived from long-chain alkenones) and the temperature calibration equations

表1 长链烯酮古温度指标()及温度校准公式Table 1 Paleotemperature index (derived from long-chain alkenones) and the temperature calibration equations
注:-表示文献中未给出相关数据,下表相同。
images/BZ_136_1210_2464_1252_2502.png指标序号 公式指示意义 参考文献1images/BZ_136_751_2594_1089_2664.png海水表层温度[15]2images/BZ_136_799_2680_1041_2751.png[6]3images/BZ_136_749_2766_1091_2837.png[18]温度校准公式样品来源 公式 适用范围 样品数 r2 标准误差 参考文献4培养实验images/BZ_136_886_3013_1111_3054.png8~25℃ 20 0.994 - [6]5images/BZ_136_880_3087_1117_3128.png8~25℃ 22 0.994 - [16]6全球大洋表层沉积物images/BZ_136_880_3161_1117_3203.png0~29℃ 370 0.958 1.5 [17]
2 四醚膜脂古温度指标TEX86
早期气相色谱样品制备需要采用冗长的化学裂解步骤,并带有一定危险性,因而古菌类脂研究受到一定程度的限制。随着液相色谱-质谱(LCMS)的发展,Hopmans等通过大气压化学电离源可以方便快速地检测古菌类脂,特别是甘油二烷基甘油四醚脂(Glycerol Dialkyl Glycerol Tetraethers,GDGTs)[33],此后这类四醚膜脂化合物受到了广泛关注。该物质是一类碳链与甘油分子通过醚键连接的单分子层生物膜脂化合物,中间的烷链骨架有类异戊二烯结构和甲基支链结构两种主要类型。基于此GDGTs可分为两类:类异戊二烯GDGTs(isoprenoid GDGTs, iGDGTs)和 支 链GDGTs(branched GDGTs, bGDGTs)。iGDGTs的碳链骨架一般具有0——4个五元环结构,其中泉古菌醇(Crenarchaeol)较为特殊,具有1个六元环和4个五元环结构[34],且立体异构体(Crenarchaeol')与其伴生出现(图2a)[35]。
iGDGTs来源于古菌,包括泉古菌(Crenarchaeota)、奇古菌(Thaumarchaeota)和广古菌(Euryarchaeota)3个主要类群。早期学术界曾认为,古菌只生存在以高盐、高温和缺氧等极端条件为特征的生态环境中,然而,21世纪初通过对环境样品中16S rDNA和脂质分析发现,iGDGTs的来源比预期要广泛得多,普遍存在于海洋、湖泊以及土壤等各种各样的环境中[36]。iGDGTs和不同类群古菌之间一般没有特定的对应关系,同一种iGDGTs可由不同的古菌门类产生,同一古菌门类可产生多种iGDGTs,这种多源性使沉积物中iGDGTs的来源复杂化。如,GDGT-0生物来源较为广泛,除嗜盐古菌外,三类古菌几乎都能合成它[37],它在高含量时常被用来指示产甲烷作用[38]。GDGTs 1-3的生物来源有奇古菌、泉古菌以及部分广古菌(包括产甲烷古菌、热原体目、厌氧甲烷氧化菌等)[37,39]。而Crenarchaeol目前被认为仅由奇古菌产生,在海洋环境中是海洋奇古菌的标志物[40]。它的立体异构体Crenarchaeol'一般认为也来源于奇古菌[41],但也存在不同观点,至今未有定论。
古菌培养实验表明,iGDGTs的构成对温度变化非常敏感,当温度升高时,它会增加碳链上五元环的个数以维持细胞膜脂的稳定性(图2b)[42]。根据这一机制,Schouten等[43]2002年对全球海区15个站点的44个表层沉积样品进行了统计分析,建立了含有86个碳原子的古温度指标TEX86(TetraEther indeX of 86 carbon atoms,表2),并提出了TEX86与SST的线性回归方程(表2-9)。方程中剔除了丰度较高的GDGT-0和Crenarchaeol,以避免对TEX86产生过大影响。随着研究海域的扩大,研究人员不断提出新的TEX86-SST校准公式。 Sluijs等[44]对北冰洋古新世——始新世极热事件期(55 MaBP)沉积物样品研究中发现,GDGT-3含量出现异常高值,推测GDGT-3含量增加是由陆源输入引起的,于是提出了TEX86的修正版本及-SST校准公式(表2、2-6),去掉了原式中的 GDGT-3,以消除陆源输入影响。Wuchter等[36]对Crenarchaeota在5~35℃条件下进行了15个培养实验,同样发现iGDGTs五元环结构数目与培养温度呈显著线性相关(表2-5)。之后Kim等[45]分析了全球287个海洋表层沉积物样品,在剔除年均温度在5℃以下的海域及红海地区的非线性关系数据后,建立了一个更广泛的全球校准公式(表2-7)。红海的TEX86值之所以偏离全球校准,是因为在高盐、高温和低营养水平的极端环境中,iGDGTs分布可能会不同,所以需要进行区域校准(表2-8)[46]。但Liu等[47]提出了另一种非线性校准方法,把上述全部的287个样品(表2-10)都包括在内。随后Kim等[48]又提出采用两种不同的校准模型以扩大iGDGTs古温度指标的区域适用性(表2、2-11):对于(亚)极地海洋区域,Crenarchaeol'在温度适应方面几乎不发挥作用,因此剔除Crenarchaeol',建立了(L代表低温地区);排除(亚)极地海洋数据后,TEX86的对数函数与海温的相关性最好,据此建立了(H代表高温地区)。这两个指标可以分别用于重建低温(15℃以下)和高温(15℃以上)环境的SST。这些研究结果推进了TEX86作为重建古温度指标的有效性。
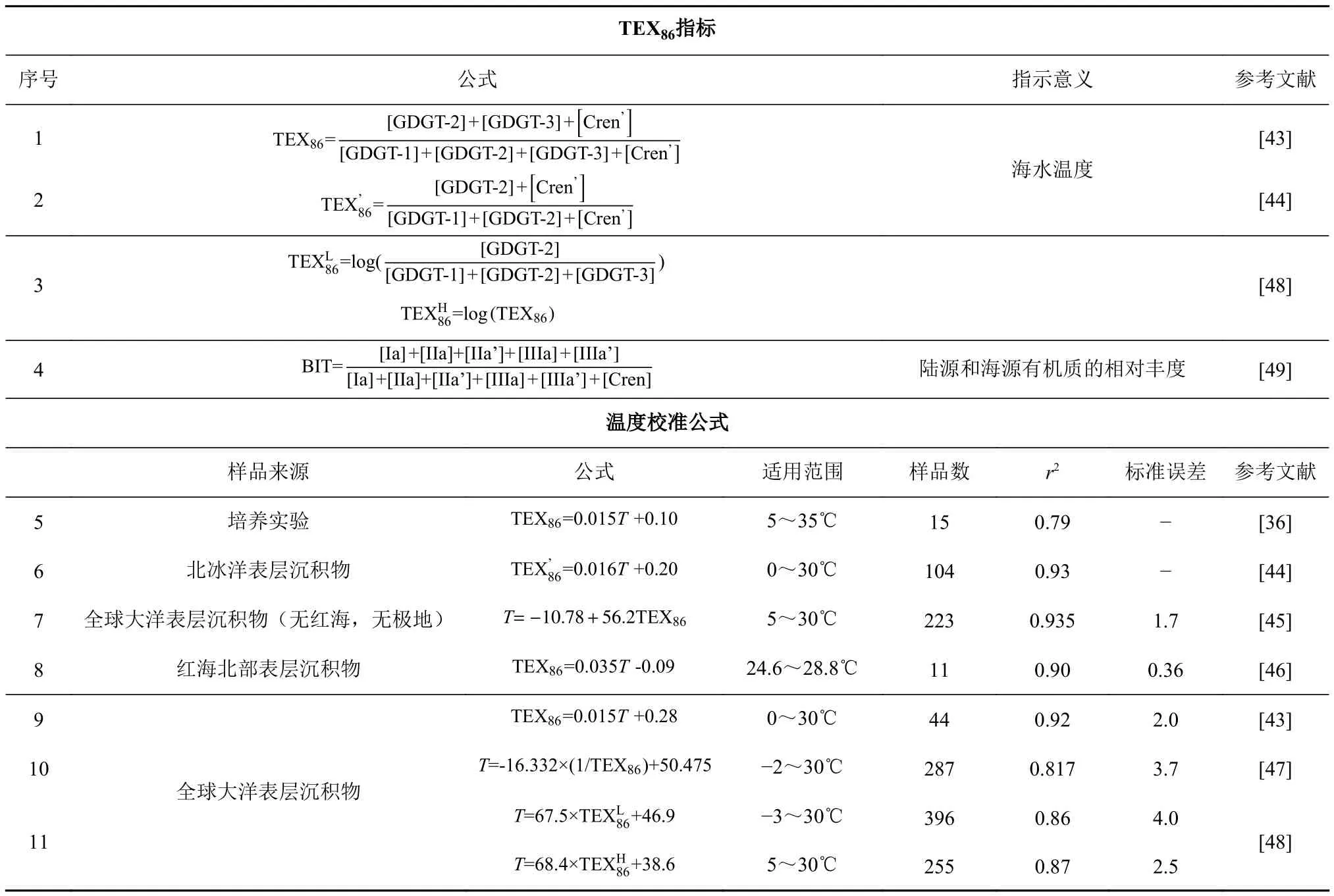
表2 四醚膜脂古温度指标(TEX86)及温度校准公式Table 2 Paleotemperature TEX86 index (derived from iGDGTs) and the temperature calibration equations
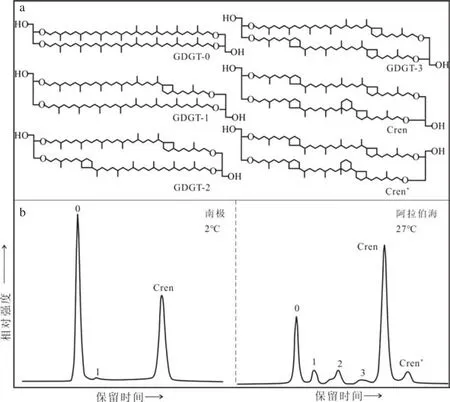
图2 iGDGTs分子结构(a)与不同温度下海洋沉积物样品中iGDGTs的代表性色谱图(b)[35, 42]Fig.2 Structures of iGDGTs (a) and representative chromatograms showing distribution of iGDGTs in marine sedimentary sample at different temperatures (b)[35, 42]
随后的研究进一步表明,TEX86指标受氧化还原条件、营养盐、pH等因素与早期成岩作用的影响较小[50],因此已成功用于全球不同海洋环境中的水温重建。例如,发现太平洋海沟表层沉积物中TEX86重建温度与实测上层海温一致[51],表明iGDGTs在向超深渊传输过程中没有被选择性降解,能够准确记录深渊海域的上层水体温度。GDGTs早期的地质记录可追溯至侏罗纪地层中[52],因此,TEX86在古老地层的温度重建方面发挥了更为重要的作用。如运用TEX86成功重建了墨西哥湾始新世——渐新世过渡期(EOT,33~34 Ma)的 SST((26±2.2)℃ 至 (33±2.2)℃)[53]及白垩纪超级暖期(125~88 Ma)赤道地区的SST(Albian早期和Cenomanian晚期——Turonian早期为32~36℃)[54]。此外,TEX86也能适用于一些大型湖泊,Powers等首次将它应用于湖泊研究中,随后该指标陆续成功应用于不同湖泊的古温度重建。如Tierney等利用TEX86合理重建了末次冰期至全新世的非洲马拉维湖表层温度变化[55]。
尽管TEX86-SST指标建立以来已广泛应用于各类环境、不同时期的古温度重建,但研究表明,古菌生长、陆源输入、地理环境等非温度因素也会对TEX86的古温度重建产生重要影响。如①古菌生长季节和水深的影响:北海沉积物的重建温度主要反映了冬季海温[56],而地中海沉积物的TEX86指标则反映的是夏季海温[57],表明了海洋古菌生长季节的区域性差异对古温度重建的影响。虽然TEX86温度指标是基于表层温度推演而得,但大量研究表明,奇古菌的最大丰度出现在次表层水体,因此,TEX86也可能更多地反映了次表层水温[58]。②陆源输入的影响:TEX86公式中的iGDGTs来源于海洋奇古菌GroupI.1a,而土壤奇古菌中GroupI.1b含量较高,相同温度下GroupI.1b会比GroupI.1a生成更多的Cren'[59],导致陆源输入明显的区域(如湖泊、河口、边缘海)TEX86值会更高。bGDGTs主要来源于陆地细菌,由地表径流输送至近海区域,因此,Hopmans等 提 出BIT(branched and isoprenoid tetraether index)指标用于指示陆源输入:当BIT<0.3时,则陆源输入对TEX86指标重建海温的影响可以忽略(表2——4)[49]。但已有研究表明,bGDGTs也存在水环境下的原位产生[60],因此,BIT作为评判陆源输入对TEX86指标影响的评判标准还存在一定争议。③特定环境的不适用。在缺氧富甲烷海域,甲烷厌氧氧化古菌在缺氧环境下会产生大量GDGT1-3,影响TEX86温度信号的准确性[61]。另外,在成熟度过高的沉积有机质中,可能存在差异性降解等,TEX86-SST会偏低[62]。湖泊体系中,不同湖泊中奇古菌类型可能不同,目前尚缺乏普适于全球湖泊的校准公式,需对TEX86进行区域校准。
3 梯烷脂温度指标NL5
厌氧氨氧化作用(anaerobic ammonium oxidation:anammox)是海洋氮循环中的重要一环,是水体缺氧区、缺氧沉积物、甚至海水中氮移除的重要途径。该过程发生在厌氧氨氧化菌(AnAOB)的厌氧氨氧化体(Anammoxoxome)这一特殊的细胞器内。Sinninghe Damsté等[63]最早在厌氧氨氧化菌的培养实验中发现了厌氧氨氧化体膜中的特殊磷脂结构,由于其形似梯子,故而命名为梯烷脂(ladderane lipids),该类物质可借助高效液相色谱-质谱技术(HPLC-MS)分析检测。梯烷脂包括完整梯烷磷脂(intact ladderane phospholipids)和 梯 烷 核 心 脂(ladderane core lipids)。梯烷核心脂由3或5个线性连接的环丁烷(其中3个环丁烷相连时会再与一个环己烷相连),通过烷基链以醚键和酯键与甘油骨架相连;完整梯烷磷脂由梯烷核心脂和极性头基连接而成,甘油骨架的sn-3位连接极性头基(磷脂酰乙醇胺、磷脂酰胆碱和磷脂酰甘油),sn-1位可连接多种不同的基团(以醚或酯形式连接的烃基、梯烷结构、直链或甲基支链的烷烃等)[64],其中梯烷脂脂肪酸结构如图3a[65]所示。梯烷脂广泛存在于土壤、沼泽、湖泊、海洋、极地地区、温泉和深海热液喷口等环境中[66],产梯烷脂的细菌可以通过调节膜流动性以维持外部环境和细胞内部之间的生理稳态[67],在古环境研究中具有潜在应用价值。
梯烷脂来源于厌氧氨氧化菌,专属性强[68]。厌氧氨氧化菌属于浮霉菌门(Planctomycetes),包括Candidatus Brocadia、Candidatus Anammoxoglobus、CandidatusKuenenia、CandidatusScalindua、Candidatus Jettenia5个属[69],且不同种属厌氧氨氧化菌中梯烷脂的组分有明显差异[67]。对前4个属的厌氧氨氧化菌的分子生物学研究发现,“Candidatus A.propionicus”、“Candidatus B.fulgida”的梯烷脂肪酸主要是C20[5]结构;“Candidatus K.stuttgartiensis”、“Candidatus Scalindua”的梯烷脂肪酸主要是C20[3]结构;在4个属中,C18[5]结构浓度都是最低的,只有“Candidatus Scalindua”培养物含有C18[3]结构[70]。
自然环境中厌氧氨氧化菌生存的温度范围很大,从-2.5℃的海冰到85℃的深海热液环境中都能检测到它们的活动。Rattray等[65]于2008年分析了实验室培养、自然环境中厌氧氨氧化菌群的梯烷脂,发现C20[5]脂肪酸的含量与温度成正比,C18[5]脂肪酸的含量与温度成反比(图3b[65])。脂肪酸链长的变化可能是一种“膜内黏性调试”(Homeoviscous adaptation),即改变双层膜的组成来应对温度变化引起的膜黏度变化。基于此,Rattray等提出了NL5指标(index of ladderane lipids with 5 cyclobutane rings)来量化梯烷脂链长随温度的变化,并发现其与温度呈四阶S型回归关系(表3-1、3-2),在12~20℃之间变化显著,而在这个温度之外NL5不再随温度变化而变化。随后Rattray等[71]扩大了研究温度范围,提出了修正的温度转换函数(表3-3)。

表3 梯烷脂古温度指标(NL5)及温度校准公式Table 3 Paleotemperature NL5 (ladderane lipids) index and the tempreature calibrations equations
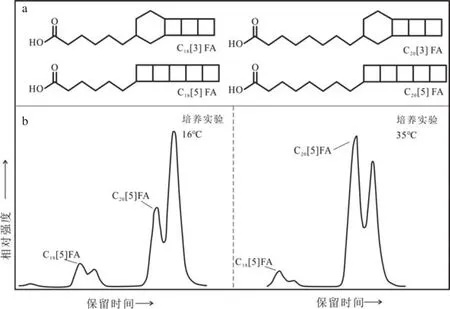
图3 梯烷脂脂肪酸分子结构(a,FA: fatty acid)和不同温度下Candidatus B.fulgida培养实验中梯烷脂脂肪酸的代表性色谱图(b)[65]Fig.3 Structures of ladderane fatty acids (FA: fatty acid) (a) and representative chromatograms showing distribution of ladderane fatty acids in cultures of Candidatus B.fulgida grown at different temperatures(b)[65]
使用NL5可以确定沉积物中梯烷脂的来源,即来自较温暖的上层水体还是相对寒冷的沉积物原位生产。这种方法已在一些海域得到成功应用,如圣巴巴拉盆地表层沉积物NL5为0.72[65],对应温度约为15℃,这远高于当地表层沉积物的温度(6℃),因此推测这些梯烷脂应该是生物分子残留物,主要来源于较温暖的上层水体。类似地,中国长江口附近的东海沉积物中NL5指数为0.54~0.89,对应的温度为15.9~23.3℃,也高于底层海水的年均温度(12~17℃)[72]。而在非洲西北部大陆架和陆坡沉积物中NL5指标出现0.11~0.22的低值,反演温度在10℃以下,与原位底层水温度(2.5~10℃)吻合,因此,该区域梯烷脂可能来源于沉积物原位生产[73]。
然而随着研究的深入,研究者逐步发现梯烷脂含量与组成在一定程度上也受其他环境因素的影响,如营养盐[74]、溶解有机碳浓度[71]、水深[73]、氧含量[75]等。而且该指标的温度适用范围较小(12~20℃),在更低或更高的温度下梯烷脂肪酸链长均无明显变化[65]。同时NL5和温度之间的关系并没有像其他温度指标一样存在很好的校准,只能用来估算温度范围。拜尔张力学说指出,正常的键角是109.5°,和正常的键角偏差越大,环的稳定性就越差,梯烷脂环丁烷的内角为90°,因此它具有高度张力[76],比较不稳定,其结构可能在早期成岩阶段发生变化,从而影响NL5值的准确性。此外,梯烷脂相对较难保存,在第四纪或更长时间尺度的温度重建中潜力不明。
4 长链烷基二醇温度指标LDI
在海洋沉积物中,除长链烯酮、GDGTs等时常被检测到之外,长链烷基二醇也是海洋沉积中常见的一类脂肪醇化合物。该类化合物分为饱和与不饱和两类,饱和二醇化合物又称长链烷基二醇(Long-chain alkyl diols, LCDs),是一种由 C1位和中间链(C11-C19)位含有羟基基团的长烷基链组成的脂类化合物[77];不饱和二醇化合物则是长碳链上含有碳碳双键的化合物,它们的分子结构如图4a[35]所示。通常用气相色谱-质谱技术(GC-MS) 检测长链烷基二醇,目前常用的仪器是三重四极杆GC-MS,其检测模式包括单粒子扫描(SIM)与多反应扫描(MRM)两种,MRM模式通过两级离子选择排除大量干扰离子,降低背景噪音,从而使目标检测物的信号值显著提高,即使在二醇含量较低的海域也能获得有效数据[78]。长链烷基二醇最早发现于黑海沉积物中[79],随着实验室培养和野外调查的展开,在全球范围内的各种海洋和淡水环境中普遍检测到了该类物质。在海洋环境中主要存在C28和C301,13-diols、C28和C301,14-diols以及C30和C321,15-diols这6类长链烷基二醇[80]。研究发现,长链烷基二醇的组成分布与温度、营养条件、上升流等密切相关,因此其在古气候、古生产力重建中具有重要意义。
长链烷基二醇生物来源复杂,目前普遍认为1,14-diols来源于Proboscia硅藻和Apedinella radians异鞭藻,湖泊系统中1,13和1,15-diols来源于真眼点藻。Volkman等[81]对真眼点藻Nannochloropsis进行实验室培养,从中检测出了C30-C321,15-diols,提出了真眼点藻作为长链烷基二醇生物来源的可能性。随后,在淡水真眼点藻的培养实验中也检测到了1,13和1,15-diols[82]。由于培养实验中观察到的长链烷基二醇与海洋真眼点藻的长链烷基二醇分布不同[83],且真眼点藻主要存在于淡水环境,海洋中含量较低,因此,在海洋环境中1,13和1,15-diols的生物来源尚无法定论[84],未来还需进一步确定它们具体的生物来源。Sinninghe Damsté等[85]对上升流区的长链烷基二醇进行稳定单体碳同位素分析,发现Proboscia硅藻是C28和C301,14-diols的主要生物来源[85]。Rampen等[86]在Apedinella radians异鞭藻中也检测出了C28-321,14-diols,该物种主要存在于河口和微咸水非上升流区。
Rampen等[80]于2012年对大西洋209个表层沉积物样品的长链烷基二醇与SST进行相关性分析,发现在-3~27℃(年均温度)范围内,SST与表层沉积物中的C28和C301,13-diols和C301,15-diol三种化合物丰度之间存在良好的线性关系,其中,前两者与SST呈负相关,后者与SST成正相关(图4b)。在此基础上,构建了长链二醇指数LDI(Long chain Diol Index,表4),并建立了LDI与SST的转换函数(表4-2)。但是最近研究发现,当SST>27℃时,甚至大于30℃时,LDI也不会接近最大值1[87],而且它与温度的关系倒转,呈负相关。因此Yang等[88]2020年重新校准了LDI与SST的函数关系(表4-3),以适用于现代实测海表温度>27℃的环境。此外,鉴于LDI会受到淡水输入和Proboscia硅藻的影响,De Bar等[89]修正了LDI与SST的校正公式(表4-4),新公式的地理覆盖范围更广,且同时考虑了这两种因素的影响,但因为有几个研究区的LDI与年均SST相关性较差,具体原因目前尚不清楚,使得这个公式校准误差较大(3℃)。

表4 长链烷基二醇古温度指标(LDI)及温度校准公式Table 4 Paleotemperature LDI (long chain diol index) and the calibration equations
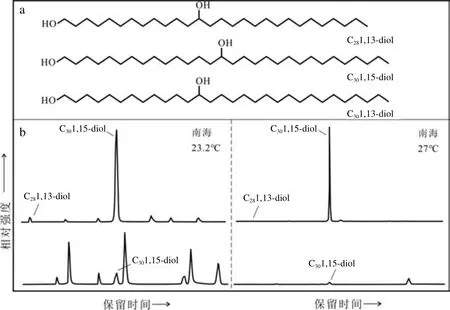
图4 长链烷基二醇分子结构(a)和不同温度下海洋沉积物样品中长链烷基二醇的代表性色谱图(b,m/z 313:C281,13二醇、C301,15二醇,m/z341:C30 1,13二醇)a改绘自[35],b未发表数据。Fig.4 Structures of long-chain alkyl diols (a) and representative chromatograms showing distribution of long-chain alkyl diols in marine sedimentary sample at different temperatures (m/z 313:C281, 13-diol, C301,15-diol, m/z341:C301, 13-diol) (b)a is modified after[35], and b is from unpublished data.
尽管这类化合物还没有确切的海洋生物来源,但由于其在地质环境中难以被降解,LDI指标已被广泛应用于古气候-环境重建工作。对南大西洋刚果河口区和北大西洋中纬度区域的沉积物应用LDI和两种指标,重建温度记录的相似性表明,LDI指标可以很好地重建大西洋的年均古温度[80,90]。粤东上升流区的LDI指标重建海水表层温度与SST几乎一致,同时也很好地记录了ENSO波动趋势,说明LDI是重建上升流区SST的有效指标[91]。此外,LDI在整个第四纪的研究中都具有很好的潜力,如澳大利亚东南部海域、地中海西部和海中部、鄂霍次克海中部和智利边缘海[92]。
影响长链烷基二醇构成和分布的因素不只有温度,淡水输入[93]和氧化降解[94]也会在一定程度上影响LDI反演结果的准确性。此外,长链烷基二醇尚不确定的生物来源也是重要的影响因素。Rampen等[83]对62个湖泊的LDI进行检验,发现LDI与SST的相关性较弱(r2=0.33),可能因为不同湖泊的真眼点藻种属不同,导致长链烷基二醇分布也不同。在受陆源输入影响较大的河口和沿海环境,高丰度淡水真眼点藻1,13-和1,15-diols的输入也会使LDI反演结果偏离预期值[93]。在冰岛附近的大西洋海域,LDI与SST的相关性也较弱,研究发现,此现象应该与Proboscia硅藻是该地区1,13和1,15-diols的重要生物来源有关[95]。同时LDI反演的SST也可能存在区域性差异,例如,在澳大利亚东南部海域,LDI温度指标更好地反映了该区域冬季海水温度,但在冲绳海槽,LDI重建的温度偏向于暖季。相比之下,在南海北部上升流区,LDI重建的SST很好地指示了年平均海温[96]。
5 羟基四醚膜脂古温度指标RI-OH
随着液相色谱-质谱技术的发展,越来越多GDGTs的化合物被识别,为新的气候指标的建立提供了思路。使用高效液相色谱-质谱技术(HPLC-MS)可以检测出羟基化类异戊二烯GDGTs(hydroxylated isoprenoid GDGTs, OH-GDGTs)。2009年,Lipp等[97]首次在海洋次表层沉积物样品中检测出该物质。随后Liu等[98]确定了OH-GDGTs的结构,即在具有0——2个五元环的iGDGTs的烷烃支链上又连接了1——2 个羟基(图5a)[35]。研究表明,OH-GDGTs广泛分布于海洋沉积物中,具有作为生物标志物的潜力。
北欧海[99]和波罗的海[100]表层沉积物中iGDGTs和OH-GDGTs之间的正相关性表明,它们可能有共同的古菌来源。已知海洋环境中奇古菌I.1a[101-102]和产甲烷广古菌[103]均能合成OH-GDGTs,底栖古菌也可能产生OH-GDGTs[104],同时也不能排除其他泉古菌或广古菌对海洋OH-GDGTs的潜在贡献,因此,古菌群落的多样性会导致OH-GDGTs生物来源复杂。此外,目前发现土壤中OH-GDGTs的含量较低,而且土壤奇古菌中含量最丰富的类群GroupI.1b无法合成OH-GDGTs[105]。
Huguet等[106]发现海洋表层沉积物中OHGDGTs的相对丰度随纬度的增加而增加,在高纬寒冷环境中尤为丰富,且与海表温度存在显著的相关性。Fietz等[99]发现OH-GDGTs中五元环的数量与亚极区和极区海温成正比(图5b),因此认为环的数量可以作为指示亚极区和极区海温的代用指标。Lü等[107]于2015年测定了54个中国近海表层沉积物样品中OH-GDGTs的分布丰度,构建了环化指数RI-OH(the ring index of OH-GDGTs)及其温度经验公式,并认为RI-OH可应用于全球海洋(表5)。但由于极地样品中缺乏OH-GDGT-2,导致基于Fietz等的数据计算出的RI-OH值均为1,因此Lü等修订了RI-OH,将极地样品中含量较高的OH-GDGT-0 纳入其中(RI-OH'),并建立了 RI-OH'与 SST 的相关性方程(表5)。当温度<15℃时,RI-OH'-SST的准确性更高(残差小),而当温度>15℃时,RI-OHSST的准确性更高。

表5 羟基四醚膜脂古温度指标(RI-OH)及温度校准公式Table 5 Paleotemperature RI-OH indices derived from OH-GDGTs and the tempreature calibration equations

图5 OH-GDGTs分子结构(a) 以及不同温度下海洋沉积物样品中OH-GDGTs的代表性色谱图(b)a改绘自文献[35],b未发表数据。Fig.5 Structures of OH-GDGTs (a) and representative chromatograms showing distribution of OH-GDGTs in marine sedimentary sample at different temperatures (b)a is modified after [35], and b is from unpublished data.
由于土壤中OH-GDGTs的含量较低,因此,RIOH指标可有效弥补在近岸海域由于陆源输入干扰导致重建古水温存在较大偏差的问题,成为重建近岸海水古温度的一个潜在重要指标。RI-OH指标已经成功应用于海洋环境中,如西太平洋边缘海[107]、南大洋[108]、弗莱姆海峡[109]、地中海[110]等地的古温度重建。
除温度外,RI-OH指标也受陆源输入[111]、季节性生长[112]等其他因素的影响,影响机制与TEX86类似,故不再赘述。值得注意的是,在南海区域的研究表明,高于25℃时RI-OH与温度呈负相关,与之前的研究结果相反[113],这说明OH-GDGTs对温度的响应机制很复杂,目前尚不明确,所以还无法确定RI-OH的全球适用性。
6 3-羟基脂肪酸温度指标RAN13
3-羟基脂肪酸(3-hydroxy fatty acids,3-OH-FAs)作为环境变化指标的研究才刚刚开始,最常用的检测方法是气相色谱-质谱技术(GC-MS)。3-羟基脂肪酸又称β-羟基脂肪酸,是革兰氏阴性菌外膜上脂多糖的主要成分[114],含10——18个C原子,并在距离羧基的第三个碳位上连接一个羟基(图6a)[115],分为正构(normal)、异构(iso)和反异构(anteiso)三种类型。典型3-羟基脂肪酸通过酯键或酰胺键与葡糖胺结合[116],在各种沉积环境中保存完好。作为环境中革兰氏阴性细菌的生物标志物,3-羟基脂肪酸广泛存在于各种环境中,如海洋、湖泊沉积物、洞穴、土壤等。
C10-C183-羟基脂肪酸主要来源于革兰氏阴性菌外膜[117-118]。已有的海洋沉积物、湖泊沉积物和土壤16S rRNA基因测序表明,在门水平上,海洋生产者以变形菌(Proteobacteria)(约94%)为主,湖泊生产者以变形菌(Proteobacteria)(约55%)、拟杆菌(Bacteroidia)(约 14%)和绿弯菌(Chloroflexi)(约10%)为主,土壤生产者以变形菌(Proteobacteria)(约 51%)和酸杆菌(Acidobacteria)(约 24%)为主;在纲水平上,生产者中变形菌的组成存在显著差异,海洋中可能是 α-和 γ-变形菌,湖泊中可能是 β-、γ-、δ-变形菌,土壤中可能是 α-、β-、γ-、δ-变形菌[119-120]。有研究表明,C10-C183-羟基脂肪酸还可能来源于革兰氏阳性乳杆菌[121]、厌氧甲基营养型细菌[122]、微藻[123]、蓝细菌[123]。3-羟基脂肪酸的生物来源至今尚不明确,未来还需要开展大量工作,包括培养实验和遗传分析,以确定陆地和水体环境中的3-羟基脂肪酸的准确生物来源。
Wang等[114]对神农架的26个土壤样品的羟基化膜脂进行分析,探讨了控制3-羟基脂肪酸相对分布的环境因素。脂肪酸中的支链可以增加细胞膜的流动性和通透性,为了使细胞内部的pH呈中性,支链脂肪酸比例会随pH升高而增加,基于此研究者提出了支链比(branching ratio)指标(表6-9)。该指标与土壤pH呈指数关系(表6),与年均气温、年均气压和土壤湿度无明显相关性,是一个独立的记录pH值的指标。又因pH为质子浓度的负对数,所以提出了分支比的负对数——RIAN指标(表6),它与pH之间呈线性关系(表6-10)。除支链比和RIAN外,研究还定义了两个与土壤pH相关的指标:支链指数与RIN(表6-11、6-12),其中支链指数值限定在0~1之间(支链比和RIAN无界),而RIN指标不需要测定反异构同系物。由于反异构脂肪酸比正构脂肪酸有更低的熔点,它能在低温条件下使细胞膜保持较好的流动性,同时研究发现C15和C173-羟基脂肪酸同系物的反异构/正构比与年平均气温(mean annual temperature,MAT)呈显著负相关,基于此,定义了RAN15和RAN17(the ratio of anteiso to normal)(表6),它们与温度线性相关(表6-13、6-14),且独立于pH。Huguet等[124]分别测定了热带和温带气候下坦桑尼亚西南部Rungwe山和意大利中部Majella山的土壤样品中3-羟基脂肪酸的浓度和丰度特征,结果表明,革兰氏阴性菌对不同纬度土壤温度变化的响应方式相同,但RAN15和MAT之间的相关性在截距上有显著差异,表明RAN15可能需要进行区域校准(表6-15、6-16)。与RAN15相比,RAN17和MAT之间的统计相似性表明可以为神农架、Majella山、Rungwe山样品建立一个统一的联合校准(表6)。Pierre等[125]利用168个全球分布的表层土壤数据集和精细的统计工具,研究3-羟基脂肪酸作为MAT和pH指标在全球范围内的适用性,结果证实,3-羟基脂肪酸的衍生环境指标(RAN15、RAN17和RIAN)与MAT 或pH 值之间的强线性关系只能在局部水平上获得,它们无法作为全球环境代用指标。与线性回归相比,多元线性回归、k-NN算法和随机森林算法得到的MAT和pH与3-羟基脂肪酸相对丰度之间存在很强的全球相关性,证实了3-羟基脂肪酸在全球水平上作为环境代用指标的潜力。革兰氏阴性菌是海洋中占据主导地位的原核生物[126],在海洋颗粒物和沉积物中均能检测到大量3-羟基脂肪酸[127],研究海洋沉积物中3-羟基脂肪酸的分布,进而可以明确基于3-羟基脂肪酸的环境指标能否应用于海洋环境。2020年,Yang等[120]分析了45个北太平洋表层沉积物样品中的3-羟基脂肪酸,发现土壤温度代用指标RAN15和RAN17与海温均无线性关系。与土壤相比,海洋沉积物中反异构C133-羟基脂肪酸同系物丰度更高,同样基于反异构/正构比与MAT呈负相关这一现象(图6b)[120],提出了一种新的基于3-羟基脂肪酸的海表温度指标RAN13(表6),它与SST呈指数关系(表6-18)。在土壤样品中RAN13与MAT没有相关性[114,124,128],因此RAN13是一个独立的海洋温度指标,即使在边缘海盆地,它也几乎不受陆源信号的影响。而后Yang等[118]又分析了24个中国碱性湖泊表层沉积物样品中3-羟基脂肪酸的组成和分布,发现基于海洋3-羟基脂肪酸的温度指标(RAN13)与MAT呈中度线性关系(r2=0.5)。由于陆源输入和湖泊原位生产的混合贡献,基于土壤3-羟基脂肪酸的温度指标(RAN15和RAN17)也不适用于湖泊环境,故而提出了一种新的湖泊环境温度代用指标 RIN17(the ratio of iso to normal)用于湖泊系统古温度重建(表6-19)。
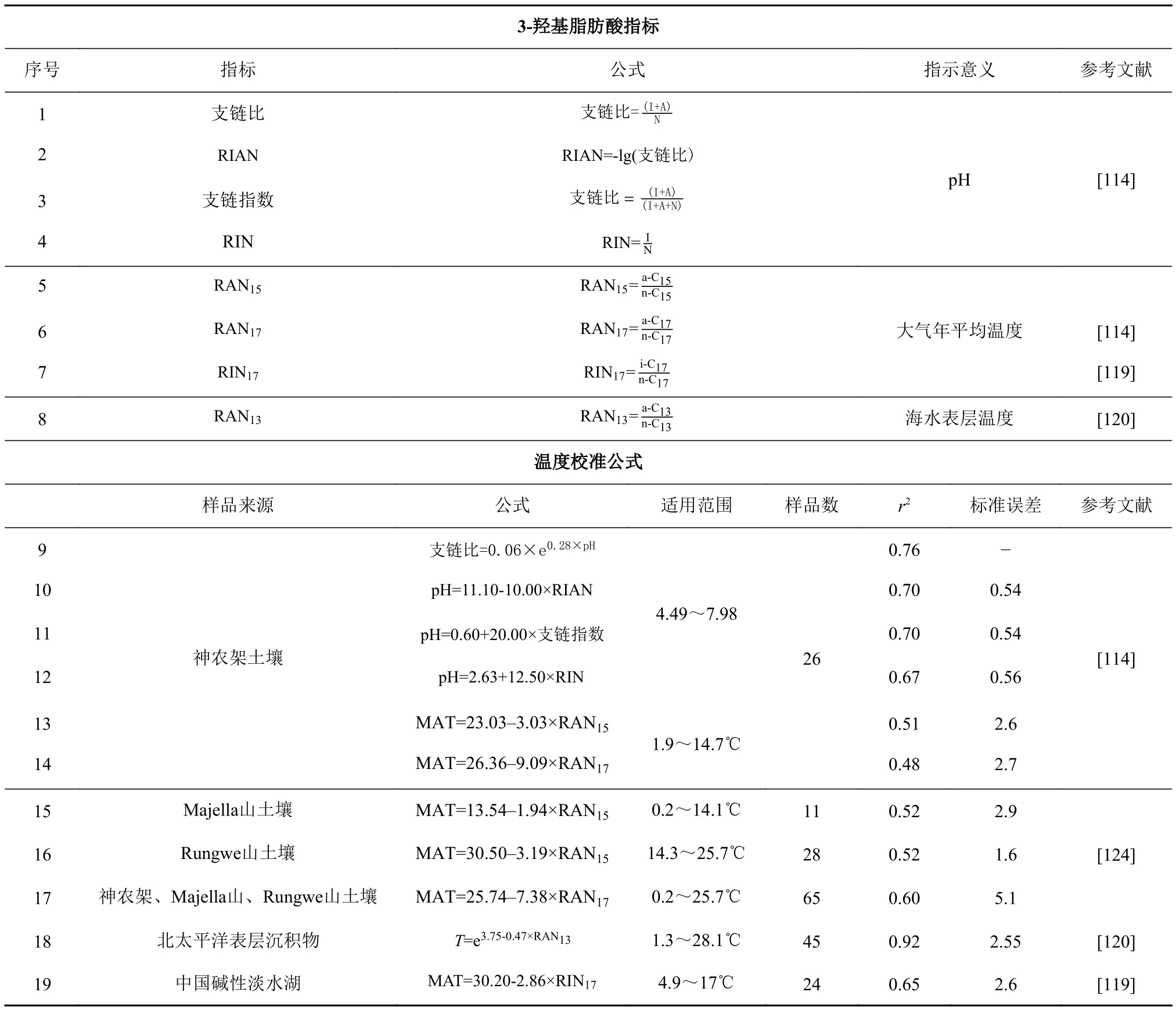
表6 3-羟基脂肪酸指标及温度、pH校准公式Table 6 The 3-OH-FAs index, and paleotemperature and pH calibration equations
3-羟基脂肪酸作为最近提出的温度指标在各种环境中都有广泛的适用性,对古气候和环境研究具有潜在意义。该指标不依赖于pH,可以独立用于重建温度,这在pH值高度可变且可能是混淆变量的环境中是一个优势,同时3-羟基脂肪酸化合物可以使用GC-MS和GC-FID识别和精确量化,所以可以利用少量的样品获得高分辨率的古气候记录。利用RAN15和RAN17重建9 kaBP以来长江中游和尚洞石笋的温度,记录显示与器测温度及δ18O记录一致[128],东海沉积物中RAN13的重建温度与过去58年来的器测海表温度相符[120],在湖泊环境中RIN17也显示出温度重建的潜力[119],这些结果都证实3-羟基脂肪酸作为古温度代用指标的可靠性。
基于3-羟基脂肪酸的温度指标也存在其局限性。应用该指标重建陆地温度时,默认土壤温度是大气温度的近似值,然而大气与细菌生长的土壤环境可能存在温差,同时后者还受太阳辐射和土壤水分等多重因素的影响[129-130]。此外革兰氏阴性菌的多样性、土壤固有的异质性、降水量/土壤水分、植被类型或季节性等其他因素也可能影响土壤3-羟基脂肪酸的相对分布,从而影响RAN15和RAN17值[124]。不同土壤之间微生物多样性存在差异,导致建立RAN15/RAN16-MAT的全球线性校准存在一定困难[131-132]。同时,3-羟基脂肪酸在某些环境中的生物来源复杂,可能混淆或掩盖其反映的环境信息[114],例如湖泊沉积物中的3-羟基脂肪酸可能来源于周围土壤。最后,该指标在海洋应用中的局限性还需要进一步研究来确定。
7 对比与展望
7.1 多指标对比
近年来,基于不同有机指标开展的古温度重建工作表明,每个指标因受到不同程度母源生物生长习性、选择性降解、环境变化等因素的影响,在实际应用中会显示出一定的差异(表7)。因此,在不同的环境背景下联合使用多种温度指标,既可以通过比较不同温标的时空性差异以提高结果的准确度,又能避免使用单一代用指标的局限性。

表7 不同脂质古温度指标综合对比Table 7 Comparison among different lipid-based temperature proxies
Yang等[120]利用RAN13反演了东海B3沉积岩芯过去58 年的温度,结果为16.6~23.3℃(平均20.2℃)。温度趋势与仪器监测海温一致,记录了1959——2017年的长期升温过程,20世纪70——80年代有约4℃的振荡,20世纪90年代以来海表温度持续上升。该结果与来自邻近站位的TEX86-SST重建结果对比表明,两个指标反演的平均温度接近,然而与RAN13相比,TEX86既没有重建仪器监测数据中的长期变暖趋势,又观察不到年代际振荡(图7a),这似乎表明西太平洋边缘海陆相有机质的季节性高输入并不会覆盖RAN13的温度信号,同时RAN13指标记录了更敏感的气候变化信号,突出了其作为边缘海温度指标的潜力。Zhu等[92]利用、以及LDI三种脂类古温度指标重建了南海北部海岸的温度记录(图7b)。该区域上升流活动较强,导致养分可利用性强,夏季定鞭藻大量繁殖,使得衍生海温偏夏季,同时上升流导致了夏季海温降低,因此衍生海温偏夏季但偏冷。奇古菌更适应于寡营养盐的冷水环境,这导致低估了海温。和分别反映了较高和较低的海温,如图7所示,LDI重建的温度在二者之间,说明LDI互补于这两个指标,提供了更全面的古海温演化信息。

图7 东海B3站位RAN13和指标反演年均温度对比(a),南海S101站位LDI和A9站位、指标反演年均温度对比(b)虚线为1959——2017年间的温度线性趋势,S101站位LDI-SST可当作A9站位的年均温度[92, 120]。Fig.7 Comparison in annual average SST between RAN13 and inversed ones in Site B3 (a) and among the LDI in S101,and and inversed ones in Site A9 (b)The dashed lines are linear trend from 1959 to 2017; Site S101 can be used to serve as a representative of annual mean SST in Site A9 [92, 120].
总之,每个指标都有其优缺点,没有一种指标可以适用于所有情况,采用多指标法可以获得更可靠的古温度变化记录,揭示气候演化历程。
7.2 总结与展望
脂质生物标志物来源于活的生物体,其对环境响应的灵敏度及准确度较高,虽然仅占沉积物中有机质的很少一部分,但其组成和结构相对稳定,保存了母源生物脂类物质的基本信息特征,特别是母源生物在生长代谢过程中气候环境因子的变化信息,是追溯有机质来源、重建区域气候环境演变的重要材料。本文从生物来源、古温度重建机制、影响因素等方面总结概述了6种有机脂类重建古海洋温度指标的研究进展。
随着检测技术的进步、灵敏度的提高,研究区域的扩展、程度的深入,有机脂类与环境间的相互关系不断被发现,有机脂类温度指标也在不断发展,并被应用于越来越广泛的区域,而各指标由于生物来源的差异分别适合不同时间尺度、区域范围,多指标联合使用以及跨学科研究特别是有机地球化学脂类与分子生物学技术相结合,从分子水平系统性了解生物、特殊脂类物质和环境间的相互关系,将有效推动有机古温度指标的发展和完善,得到准确、可信的古温度重建信息,从而提升古气候研究至一个新高度,这对分析气候演化规律、探讨全球气候变化特征及预测未来气候演变都具有重要意义。
猜你喜欢
海洋通报(2022年2期)2022-06-30
海洋石油(2021年3期)2021-11-05
昆明医科大学学报(2021年4期)2021-07-23
铁路通信信号工程技术(2021年3期)2021-04-03
河北环境工程学院学报(2021年1期)2021-03-19
肿瘤防治研究(2020年8期)2020-09-11
空间科学学报(2020年6期)2020-07-21
合成树脂及塑料(2020年3期)2020-01-16
中成药(2017年5期)2017-06-13
人间(2015年11期)2016-01-09
