
黄土高原生物土壤结皮发育过程中颗粒态和矿物结合态有机碳变化特征
2023-01-09 03:18杨娥女王宝荣姚宏佳黄懿梅安韶山
水土保持研究 2023年1期
杨娥女, 王宝荣, 姚宏佳, 黄懿梅, 安韶山,3
(1.西北农林科技大学 水土保持研究所, 黄土高原土壤侵蚀与旱地农业国家重点实验室,陕西 杨凌 712100; 2.中国科学院 水利部 水土保持与生态环境研究中心, 陕西 杨凌 712100;3.中国科学院大学, 北京 100049; 4.西北农林科技大学 资源环境学院, 陕西 杨凌 712100)
土壤有机碳库作为陆地生态系统最大的碳库[1],其微小波动将可能对大气二氧化碳(CO2)浓度甚至全球碳平衡产生重大影响[2]。最近的研究表明土壤有机碳在很大程度上其实是微生物残体、植物分子聚合物以及它们的降解产物所组成的复杂混合物。植物源有机碳和微生物源有机碳主要贡献有机碳的积累过程[3]。由于植物源有机碳和微生物源有机碳在形成、周转时间和功能方面的差异,最近的研究将土壤有机碳(Soil organic carbon,SOC)划分为颗粒态有机碳(Particulate organic carbon,POC)和矿物结合态有机碳(mineral-associated organic carbon,MAOC)两种形式[4-6]。POC以植物大分子物质为主,具有高的C∶N比和较快的周转速率,而MAOC则以微生物源组分为主,具有较低的C∶N和较慢的周转速率[5]。由于这两种碳库的有机碳来源不同及物理保护机制的差异,POC和MAOC的变化特征影响土壤有机碳的稳定性。POC和MAOC含量及其比例受植被类型影响。例如,Cotrufo等[4]关于欧洲草地和森林生态系统土壤POC和MAOC的研究表明,MAOC占土壤总有机碳的比例超过50%,草地和丛枝菌根林在MAOC中储存了更多的土壤有机碳,而外生菌根林在POC中储存了更多的土壤有机碳。而MAOC吸附的有机碳分子主体来源为微生物产物及其残体物质。土壤黏粒和粉粒对SOM的吸附作用是土壤有机质稳定的主要因素[7]。森林生态系统枯落叶是土壤有机碳的主体来源[8],不易分解的植物残体以物理迁移方式进入土壤主要贡献POC的增加。草地生态系统由于地上植物组分低的生物量,地下植被组分(如根系分泌物和细根)成为土壤有机碳的主体来源[9]。这些小分子的根系分泌物被微生物利用进而直接形成一系列MAOC主要贡献土壤有机碳的增加过程。
生物土壤结皮(Biological soil crusts,BSCs)是由藻类、地衣、苔藓等隐花植物、细菌、真菌与土壤表层颗粒胶结形成的复杂有机复合体,占沙漠生态系统活体覆盖的40%以上[10]。结皮定殖和发育过程有效地改善了土壤的物理、化学和生物学特性[11-12],主要由于BSCs中固碳微生物和初等绿色植物可以固定大气中的碳氮和捕获空气中悬浮的细土壤颗粒稳定地表,进而促进土壤发育[13]。从最初的CO2微生物碳固定过程到随后的初等维管植物发育进而贡献土壤有机碳和养分增加过程。随着国家退耕还林还草政策的实施,更有利于黄土高原BSCs定殖、发育和分布[14-16]。主要由微生物组成的生物土壤结皮成为退耕地上普遍存在的地被物,广泛分布于草原区,森林草原区,森林区,其中在草原区的盖度可以达到60%~70%[17],BSCs在提高土壤表面稳定性、增强土壤抵抗水蚀风蚀能力、改善土壤养分和促进土壤发育等方面发挥着重要作用[11]。
目前,关于BSCs对干旱和半干旱区生态系统功能作用已有大量研究[18],然而,在植被生产力受限和黏粒含量缺乏的沙地生物土壤结皮中POC和MAOC变化特征尚不清楚。本研究将土壤有机碳库划分为POC(>53 μm)和MAOC(<53 μm),探究黄土高原BSCs发育过程中POC和MAOC的变化特征及其对有机碳积累的相对贡献。试图阐明BSCs发育中沙质土壤POC和MAOC的形成过程,丰富干旱半干旱生态系统SOC形成和稳定过程的理论依据。
1 材料与方法
1.1 研究区概况
研究区地处陕西省榆林神木六道沟流域(38°46′—38°51′N,110°21′—110°23′E)。流域面积6.89 km2,海拔1 081~1 274 m,主沟道长 4.21 km,100 m以上的沟道密度达 6.45 km/km2,沟谷面积占流域面积的 38.1%[19]。年平均气温9~14℃,属于干旱半干旱季风气候,寒暑分明,气候比较干燥,年潜在蒸发量超过1 000 mm,降水年际变化较大,多年平均降水量为437 mm,且5—10月占全年降水的65%~85%。在该地区,生物结皮分布广泛,由藻类和以苔藓为主的结皮组成。土壤黏粒(<0.002 mm),粉粒(0.002~0.05 mm)和砂粒(0.05~2.0 mm)的含量分别为0.33%~0.78%,9.81%~21.68%和77.54%~88.96%。

表1 研究区沙地基本信息
1.2 样品的采集和处理
选择神木六道沟流域采集不同发育阶段的生物土壤结皮样品,包括藻结皮(algae),藻结皮(algae>80%)+少量藓结皮(moss <20%),藓结皮(moss>80%)+少量藻结皮(algae<20%),藓结皮(moss)4个发育阶段的生物土壤结皮和裸沙(Bare sand)作为研究对象。在每个结皮发育阶段随机选择5个40 m×40 m的样方(即5个野外重复),每个样方按“S”点选取12—15点,多点收集生物土壤结皮层,结皮层下0—2 cm,2—10 cm,10—20 cm共4个土壤层次的土壤样品。混合均匀带回实验室,剔除植物残体和其他杂物过2 mm筛,在室内风干后过0.15 mm筛。
1.3 颗粒态有机碳和矿物结合态有机碳的测定
颗粒态有机碳和矿物结合态有机碳分离采用六偏磷酸钠分散法[20],即称取过2 mm筛的风干土样20 g于100 ml的塑料瓶中,加入60 ml六偏磷酸钠(质量/体积=5%),手动摇匀后在摇床上震荡18 h。将混浊液过53 μm筛子,反复用去离子水冲洗,直至水流清澈为止。筛上粗质部分为POC(>53 μm),筛下细质部分为MAOC(<53 μm),用已知质量的500 ml烧杯收集上下组分,经60℃烘干称重并记录,计算其所占全土的质量占比。将烧杯中烘干土壤收集研磨过0.15 mm筛,用重铬酸钾—外加热法测量其SOC含量[21]。土壤中POC,MAOC含量及质量占比(%)计算公式如下:
POC质量分离比=m颗粒态/m
POC=SOC颗粒态×POC质量分离比
POC绝对量=SOC颗粒态×m颗粒态
式中:m颗粒态表示分离后颗粒态有机碳组分的质量;m表示分离前的总土壤质量;SOC颗粒态表示颗粒态组分测得的土壤有机碳;MAOC计算将上述POC替换为MAOC即可。
1.4 黏粒含量测定
本研究采用USDA分类系统标准(砂粒范围0.05~2 mm,粉粒范围0.002~0.05 mm,黏粒范围<0.002 mm)进行土壤砂粒、粉粒、黏粒的划分。具体的操作步骤为:将土壤样品风干后磨碎通过2 mm筛,用烘干法测定风干土壤的吸湿水含量。用万分之一天平称取10 g土样,倒入500 ml的三角瓶中,后加入10 ml的浓度为0.5 mol/L的六偏磷酸钠,并加蒸馏水至250 ml,充分摇匀,盖上小漏斗,放到205℃沙浴锅中煮沸 1 h,煮沸过程中用玻璃棒经常搅拌。冷却后全部洗入1 000 ml的量筒中,最后将量筒内的悬浮液加蒸馏水至1 000 ml。根据室内温度分别确定<0.05 mm,<0.02 mm和<0.002 mm的取样深度及沉降时间。在对应时间用吸管吸取对应的悬浊液后,移入铝盒,烘箱烘干后用万分之一天平测定其质量,最后得到各粒级土粒的比例。
1.5 统计分析
用Excel和SPSS 22.0软件进行统计分析,采用SPSS 22.0进行单因素和双因素方差分析,确定生物土壤不同发育阶段土壤颗粒态有机碳、矿物结合态有机碳的显著性(p<0.05)及双因素的交互作用,文中所有柱状图、散点图均由Origin 2018绘制,图表中数据为平均值±标准差。
2 结果与分析
2.1 生物土壤结皮发育过程中颗粒态有机碳和矿物结合态有机碳变化特征
BSCs中颗粒态组分(>53 μm)质量占比远大于矿物结合态组分(<53 μm)质量占比(图1),4个发育阶段的颗粒态组分均小于Bare sand对照,而矿物结合态组分均大于Bare sand对照。在相同发育阶段,颗粒态组分质量占比随着土壤土层加深而增加;而矿物结合态组分质量占比随着土壤土层加深而降低。
POC含量随着土层深度而降低(图2)。其中,在BSCs土层,随着BSCs发育,POC含量由大到小为MO(9.47 g/kg)>MO-AL(6.39 g/kg)>AL(6.31 g/kg)>AL-MO(3.56 g/kg)(图2A)。0—2 cm土层中,4个发育阶段POC含量都显著高于Bare sand对照(0.63 g/kg),其中AL阶段POC含量最高,为2.67 g/kg(图2B)。
MAOC含量随土层增加而降低(图2)。其中,在BSCs土层,MAOC含量由大到小为MO(12.57 g/kg)>AL(7.40 g/kg)>MO-AL(7.23 g/kg)>AL-MO(5.27 g/kg)(图2A)。与Bare sand对照相比,2—10 cm和10—20 cm土层,各发育阶段的MAOC含量都有所增加,在2—10 cm土层,MO-AL,MO,AL和AL-MO比Bare sand对照分别增加了82.6%,75.4%,74.9%和65.2%(图2C);在10—20 cm土层中,MO-AL,AL-MO,MO和AL比Bare sand对照分别增加了84.0%,55.5%,35.9%和25.7%。
在BSCs和0—2 cm土层,MAOC绝对量高于POC绝对量(图3A、图3B),在2—10 cm和10—20 cm土层,除MO-AL外,其他发育阶段POC绝对量高于MAOC绝对量(图3C、图3D),表明在BSCs层和结皮层下0—2 cm土层,微生物源有机碳-MAOC主要贡献SOC的积累;而在结皮层下2—10 cm和10—20 cm土层,植物源有机碳-POC主要贡献SOC的积累。
在BSCs土层,MO发育阶段POC和MAOC绝对量均显著高于其他发育阶段,随着生物土壤结皮发育,POC和MAOC绝对量是一个增加的过程(图3A);说明BSCs的定殖和发育促进了SOC的积累。随着土层加深,各发育阶段POC和MAOC绝对量趋于稳定(图3C,3D)。
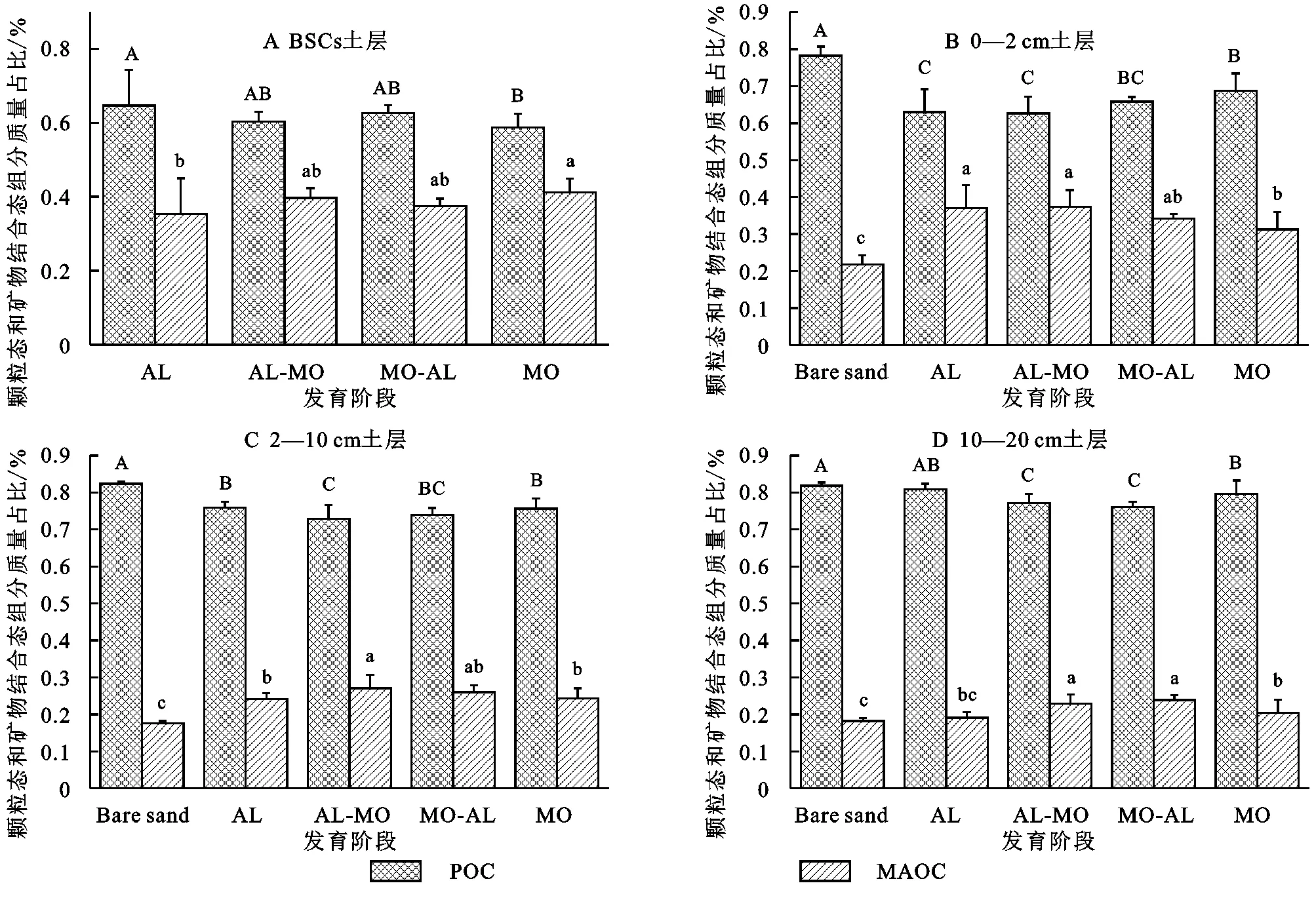
注:Bare,Al,Al-Mo,Mo-Al和Mo分别表示裸沙、藻结皮、藻结皮+少量藓结皮、藓结皮+少量藻结皮和藓结皮,误差线表示标准差;大写字母表示不同发育阶段POC的显著性,小写字母表示不同发育阶段MAOC的显著性(邓肯法,p<0.05),下同。
2.2 生物土壤结皮发育过程中颗粒态有机碳、矿物结合态有机碳与SOC的关系
随着BSCs发育,在BSCs土层,POC随黏粒含量增加而增加,拟合函数为y=3.91+1.65x,R2=0.06(图4A);MAOC随着黏粒含量增加而减少,拟合函数为y=12.17-2.66x,R2=0.15(图4B)。说明MAOC趋于饱和状态,随着黏粒含量增加,MAOC的增加速率变慢,而POC没有饱和。在BSCs和0—2 cm层土壤中,POC/MAOC与SOC的线性拟合斜率大于零(图5A),而在2—10 cm和10—20 cm层土壤中,它们的线性拟合小于零(图5B),表明与下层土壤相比,POC在表层土壤中增加更快,然而MAOC/POC与SOC的线性拟合趋势与POC/MAOC相反,表明在有机物输入相对较少的下层,MAOC增加量大于POC(图5C,5D)。

图2 生物土壤结皮发育阶段颗粒态有机碳和矿物结合态有机碳含量
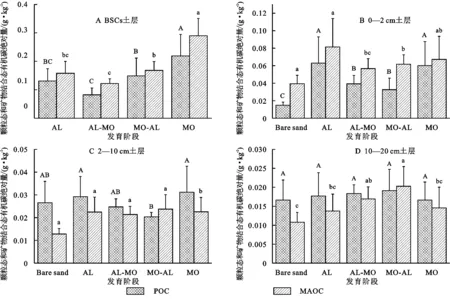
图3 生物土壤结皮发育阶段颗粒态有机碳和矿物结合态有机碳绝对量
表层土壤黏粒受限制,吸附在黏粒表面的MAOC饱和,不再吸附,更多的碳进入POC,所以POC随着黏粒增加而增加。POC/MAOC在表层随着SOC增加而增加,表明POC增加速率比MAOC快,因此在BSCs和0—2 cm土层,MAOC饱和,POC主要贡献SOC的增加(图4)。
在BSCS土层中,随着BSCs发育,POC/SOC和MAOC/SOC比值均先减少后增加。在0—2 cm土层,与Bare sand对照相比,POC/SOC的值在各发育阶段都有所增加,MO阶段增加最快;而MAOC/SOC的值都低于Bare sand对照,但随着发育进行(AL→AL-MO→MO-AL→MO),MAOC/SOC一直缓慢增加。2—10 cm土层,各发育阶段POC/SOC和MAOC/SOC值均低于Bare sand对照(表2)。线性拟合分析发现,在BSCs发育过程中POC和MAOC与SOC均存在显著的正相关关系,说明POC和MAOC的增加促进了土壤有机碳的积累(图6)。随着SOC含量增加,碳含量相对较低的土壤中,MAOC对SOC的贡献占主导地位,然而,MAOC有饱和的趋势,不再随SOC增加而增加。此时,SOC的增加通过POC的累积来实现。

图4 颗粒态有机碳和矿物结合态有机碳随黏粒含量的变化特征

图5 颗粒态有机碳和矿物结合态有机碳的比值与土壤有机碳的关系
2.3 生物土壤结皮发育阶段、土层及其交互作用对POC和MAOC的影响
为了探究生物土壤结皮发育阶段、土层及它们的交互作用对POC和MAOC的影响,我们进行了双因素方差分析。由表3可知,发育阶段、土层及发育阶段×土层交互作用的F值都大于p(p<0.05),说明发育阶段、土层及发育阶段×土层交互作用对POC和MAOC的影响均为显著,三者中土层的影响显著程度最高。其中,对POC影响的显著程度由大到小为土层(F=193.16)>发育阶段(F=12.56)>发育阶段×土层交互作用(F=9.83);对MAOC影响的显著程度由大到小为土层(F=391.34)>发育阶段(F=19.30)>发育阶段×土层交互作用(F=12.92)。
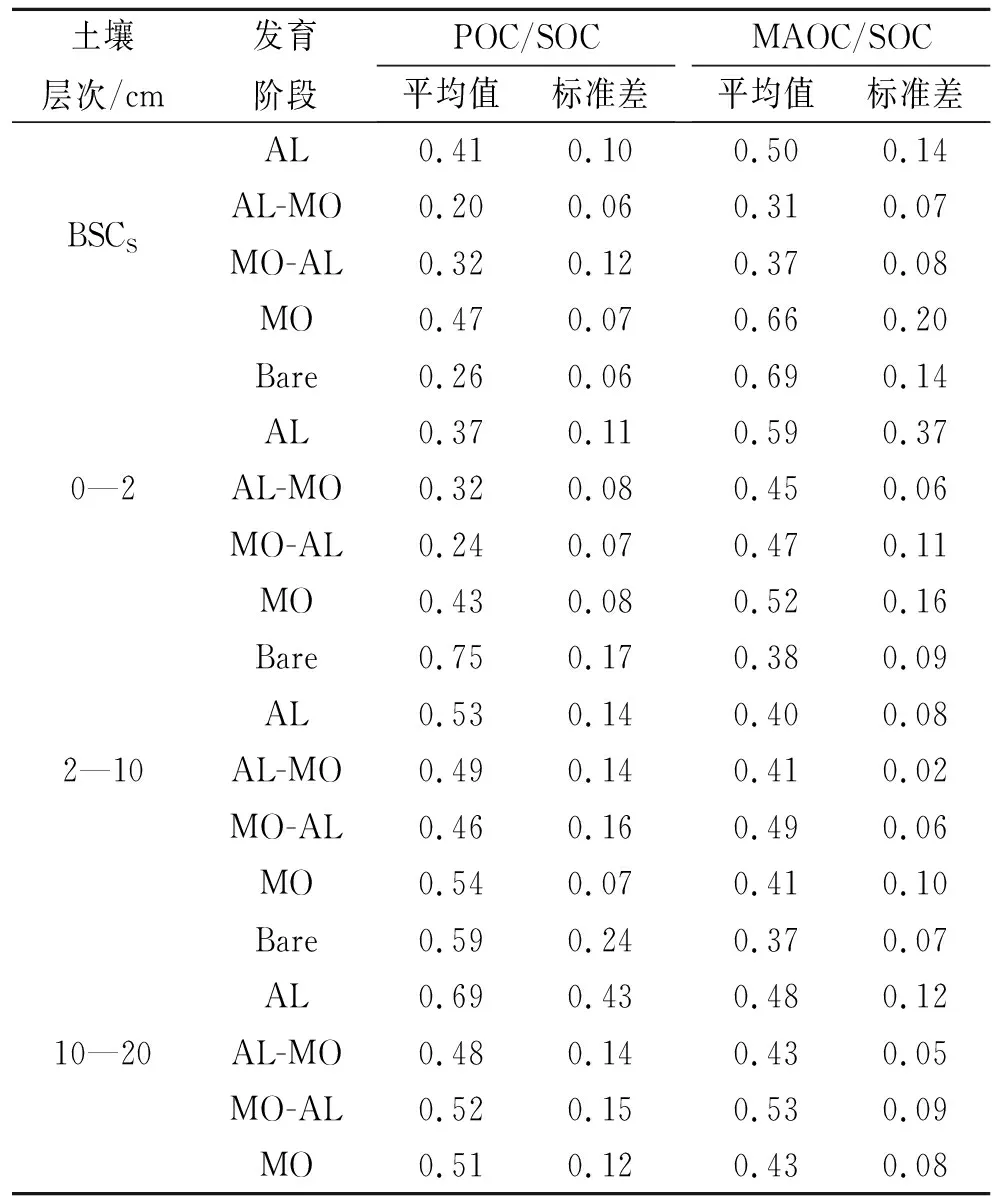
表2 颗粒态和矿物结合态有机碳与土壤有机碳的比值关系

图6 生物土壤结皮发育过程中颗粒态和矿物结合态有机碳与土壤有机碳的关系
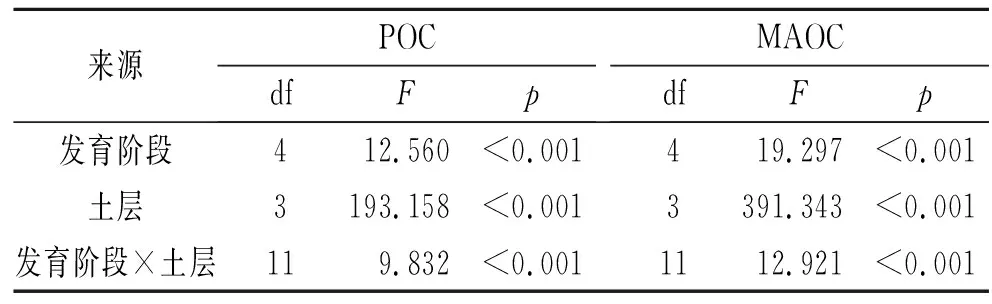
表3 双因素方差分析
3 讨 论
3.1 生物土壤结皮发育对颗粒态有机碳和矿物结合态有机碳的影响
同一发育阶段相同土层MAOC含量大于POC含量,这是由于MAOC受矿物保护,不易被微生物分解,是有机碳库中相对稳定的组分[22],这与梁爱珍等关于东北黑土区颗粒态有机碳和矿物结合态有机碳的研究结果相同[23]。而POC组分质量占比要远高于MAOC组分质量占比(图1),这是因为沙地中沙粒含量很高,大多数土壤颗粒粒径大于53 μm,小于53 μm的土壤颗粒占比较小,因此POC组分质量占比显著高于MAOC组分质量占比。
BSCs是荒漠生态系统的重要组成部分,也是干旱区碳积累和释放的关键因素[24-25]。BSCs的发育首先是抗干旱胁迫和干扰能力强的蓝藻拓殖并形成藻结皮,藻结皮的定殖改变了土壤的微环境,增加了土壤的持水能力,进而为喜湿的藓结皮发育创造条件。在BSCs层土壤中,随着结皮发育,POC和MAOC含量呈增加趋势,在MO阶段POC和MAOC含量最高(图2A,图3A)。其他研究结果也发现,BSCs的发育有利于SOC的积累[26-28]。Dümig等[29]发现由藻类结皮演替到藓类结皮,结皮层有机碳含量增加了1.8倍;Liu等[30]的研究发现随着BSCs的演替,土壤微生物丰度和多样性呈逐渐增加趋势。其次,结皮层作为空气和土壤连通的介质,其捕获大气降尘和颗粒及通过光合作用固定碳的能力随着结皮发育而增强[25,29]。此外,不同发育阶段生物土壤结皮的结构、功能和生物量存在显著差异,导致进入土壤的凋落物数量和质量存在差异[31],藓结皮阶段是生物结皮演替的高级阶段,输入的有机物多。吴丽等[32]研究了BSCs演替过程中生物量的变化,发现随着结皮的发育演替,结皮中的光合生物量呈逐步增加。
3.2 不同土层生物土壤结皮发育下颗粒态有机碳和矿物结合态有机碳
土壤有机碳的垂直分布与土壤质地和气候密切相关,SOC随黏粒含量和水分含量的增加而增加,随温度增加而降低;然而,植被覆盖对SOC的影响大于土壤质地和气候条件,覆盖植被凋落物在地上地下及根际之间的分配决定了SOC的垂直分布特征。本研究结果表明,POC和MAOC随着土层深度而降低。在BSCs层含量最高,0—2 cm土层含量次之,2—10 cm和10—20 cm土层含量要远少于BSCs和0—2 cm土层, POC和MAOC含量在不同土层间存在显著差异(p<0.05)(图2,图3)。因为生物结皮形成后,改善了BSCs层土壤的理化性质,使土壤有机质含量剖面分布呈表聚现象[33-34]。其次,结皮形成后,隐花植物和一些浅根系植物大量繁殖,他们的残体、分泌物等被微生物分解成腐殖质,导致该层有机碳的积累[35]。BSCs对表层SOC影响显著,对深层土壤影响不显著。而许多研究也有类似结果[14,36-37]。
BSCs和0—2 cm土层中MAOC绝对量高于POC绝对量,由以微生物源有机碳为主要来源的MAOC贡献SOC积累;而2—10 cm和10—20 cm土层POC绝对量高于MAOC绝对量,由以植物源有机碳为主要来源的POC贡献SOC积累(图3)。这是因为BSCs层和结皮层下0—2 cm土层微生物和土壤动物含量高[28],微生物和动物通过自身新陈代谢、坏死等残体累积有机碳,这也解释了MAOC主要是微生物源有机碳[38]。POC主要贡献2—10 cm和10—20 cm土层有机碳的原因是,一些小分子随水淋溶到下层土壤中,吸附在POC和MAOC,然而下层MAOC饱和导致更多的碳分子吸附在POC上。
3.3 颗粒态有机碳和矿物结合态有机碳与有机碳的关系
在干旱半干旱区,生物结皮广泛分布,改善了干旱半干旱的微环境,提高土壤养分,而土壤养分又促进结皮发育演替[39-40]。生物结皮在全球碳氮循环中扮演者重要角色,是干旱半干旱地区重要的碳、氮源[41-42],生物结皮层固N被认为是干旱区和N限制生态系统土壤N输入的主要来源[43],生物结皮中的隐花植物如蓝藻、地衣、苔藓等都可以固定大气中的N[44],隐花植物还通过光合作用将空气中的CO2固定在生物结皮土壤中[45]。
我们将POC/SOC和MAOC/SOC做了对比,发现在不同的发育阶段和不同土层深度,POC/SOC和MAOC/SOC没有明显的变化规律(表2)。这可能是随着生物土壤结皮发育,土壤的微环境逐渐改善,不同发育阶段的不同土层养分含量、微生物数量等分布不均匀导致的[32,39],同时,随着生物结皮发育,结皮厚度增加,结皮颜色加深,表面更加粗糙,输入土壤的凋落物数量和质量不同,所以POC/SOC和MAOC/SOC在不同发育阶段和不同土层深度没有明显的变化规律。
在我们的研究中,POC,MAOC和SOC均有显著的正相关关系(图5),表明生物土壤结皮定殖和发育显著促进了SOC积累,这与许多研究结果相似[23,46]。POC随着SOC的增加而增加,而MAOC随着SOC的增加会饱和。由于微生物生物量的生态限制(例如竞争,捕食等),土壤可能出现SOC饱和状态,随着土壤C输入增加,与矿物相关的有机碳形成率降低,矿物相关的有机碳由微生物直接产生的化合物(即微生物细胞和细胞外化合物)组成[47]。MAOC主要吸附在矿物表面,随着黏粒含量受限,矿物比表面积有限,矿物表面吸附的MAOC处于饱和状态。因此随着SOC的增加,MAOC有饱和的趋势,不再随SOC增加而增加。此时,SOC的增加通过POC的累积来实现。Cotrufo等[4]在欧洲草地和森林生态系统土壤研究中也得出了相同结果。因此,沙地土壤POC和MAOC与总SOC显著关系也体现了沙土POC和MAOC对SOC的积累和保持的重要贡献。
4 结 论
生物土壤结皮的定殖和发育能够显著增加土壤有机碳含量,在最初的土壤发育过程中,MAOC对SOC的贡献占主导地位,然而由于沙土黏粒含量的缺乏,当MAOC趋于饱和时,SOC的增加主要由POC的积累来实现,这表明黏粒含量是这种黄土高原砂质土壤中稳定碳积累的重要限制因素。在BSCs土层,由于MAOC的增加受到低粘土含量的限制, POC的累积速率比MAOC更快,MAOC处于饱和状态。在BSCs和0—2 cm土层,以微生物源有机碳为主导的MAOC主要贡献有机碳积累,在2—10 cm和10—20 cm土层,以植物源有机碳为主导的POC主要贡献有机碳积累。
猜你喜欢
新疆大学学报(自然科学版)(中英文)(2022年3期)2022-06-04
作物学报(2022年7期)2022-05-11
土壤(2021年5期)2021-11-17
水土保持通报(2020年6期)2020-03-10
农业环境科学学报(2019年12期)2019-12-23
江苏农业学报(2019年2期)2019-09-10
四川农业科技(2019年2期)2019-03-28
新疆大学学报(自然科学版)(中英文)(2018年4期)2018-02-10
科学与财富(2017年26期)2017-09-23
中国果业信息(2015年1期)2015-01-24
