
果蔬尾菜酵素发酵条件与生物活性探究
2022-12-29 07:31王一贺田少腾靳金博南松剑
湖北农业科学 2022年20期
王一贺,田少腾,靳金博,南松剑
(中国农业大学烟台研究院,山东 烟台 264670)
尾菜是指蔬菜在生长、采收、加工等环节中产生的废弃叶、根、茎等,也称“蔬菜废弃物”。2020年,中国蔬菜种植面积约为21.3万km2,蔬菜产量约为7.22亿t。随着中国果蔬种植的发展,尾菜也在不断增加,各类蔬菜在生产过程中尾菜的平均比例为33.6%,厨房加工过程中平均废弃率为20.5%[1]。目前,产地尾菜的处理方法以深埋、远倒为主,也有沼气处理、沤肥等[2],但由于处理技术存在不足,与生产实际差距较大,并且投入的人力和物力过多,应用推广难度较大[3]。尾菜若不及时收集处理,会导致资源浪费以及环境污染[4]。通过堆肥发酵处理,可使其作为废弃物得以资源化利用[5,6],发挥其具有的生态效益、经济效益和社会效益[7]。利用发酵的手段将果蔬尾菜转化为酵素,也是尾菜资源化利用的一个较好办法,发酵制得的酵素中不仅含大量的氨基酸、矿物质、功效酶等成分[8,9],而且含有大量作物所需的营养物质和生长调节物质[10]。因酵素具有的良好生物活性[11],经实践发现,酵素在种植业、养殖业都可以发挥重要作用[12-14]。
研究表明,桑椹酵素发酵过程中添加混合菌种群可以显著提高特征酶活力[15],有助于加速发酵过程,提高发酵质量[16];施用EM菌剂有效促进番茄、草莓生长,提高产量及品质[17,18];添加蔬菜类酵素发酵的滤液中含有较多霉菌,水果类酵素中含有多种酶类,其成分含量有所不同[19]。本研究通过对果蔬废弃物进行不同条件下的发酵试验,探究不同发酵条件下对产物细菌、真菌和大肠杆菌菌落总数,pH、密度等指标,纤维素酶、蛋白酶、脂肪酶和淀粉酶等功效酶活性的影响,分析果蔬酵素制备影响因素,从而优化发酵工艺,确定最优发酵因子组合,为尾菜发酵在生产中的合理应用提供参考。
1 材料与方法
1.1 试验材料
试验所用果蔬尾菜来源于牟平某农贸市场,包括蔬菜废弃物(因货架销售要求需整形、包装产生的废弃蔬菜茎、叶、根,主要品种为菠菜、油菜)、水果废弃物(红富士苹果坏果、果皮);红糖(EM菌培养适用型)、EM菌剂(混合型添加剂,地衣芽孢杆菌Ⅵ型)购于河南农富康生物科技有限公司。
1.2 试验设计
试验开始前将收集的果蔬尾菜清洗后,置于通风处自然晾干水分,消毒处理并切成1~3 cm的小块(段),装入清洗消毒过的不锈钢发酵装置中,按照料液比3∶10进行处理,加入发酵底物、添加物(EM菌剂、红糖)发酵。搅拌混合均匀后,添加不同果蔬尾菜发酵底物、添加物(EM菌剂、红糖)后密封发酵。试验开始后每2天将果蔬尾菜酵素摇匀1次,发酵过程中控制发酵温度为25℃,恒温发酵,发酵产生气体自动通过U型管排出发酵装置。
试验共分10组处理,1组为空白处理,其余试验处理具体成分和试验条件见表1。

表1 试验分组设计
1.3 项目测定
分别于发酵试验开始后的7、14、21、28、35、42、49 d取样,使用移液管量取果蔬尾菜酵素上清液50 mL,置于消毒玻璃瓶,4℃冰箱保存并进行项目检测,项目测定方法见表2。

表2 项目测定方法
1.4 数据处理
数据统计分析用Excel 2019、SPSS 18.0软件,采用LSD法作多重比较分析(P<0.05)。
2 结果与分析
2.1 酵素发酵过程中pH、密度的变化
发酵过程中,pH的变化是判断发酵规律和效果的依据之一,发酵过程中pH的变化见图1。发酵初期,各发酵处理pH变化幅度大,除T8外,其余发酵处理7 d内酵素pH均快速降至4.50以下。在后续发酵过程中,各发酵处理pH变化幅度大幅下降,T1、T6、T7、T8的pH略微升高,T2、T3、T4、T5、T9的pH略微下降,空白处理总体保持稳定,pH基本不变。仅添加EM菌剂的发酵处理发酵过程中pH上升,仅添加红糖及同时添加红糖、EM菌剂发酵处理发酵过程中pH下降。其原因可能是菌群生长过程中会消耗分解果蔬尾菜中的有机酸等物质,导致pH升高,添加糖可以增加酵素中的有机酸等物质含量,减缓消耗速度,抑制pH的升高;同时添加红糖、EM菌剂的T2、T4、T5分别为相同发酵底物中pH最低的发酵组,说明同时添加红糖、EM菌剂时,发酵效果最佳。
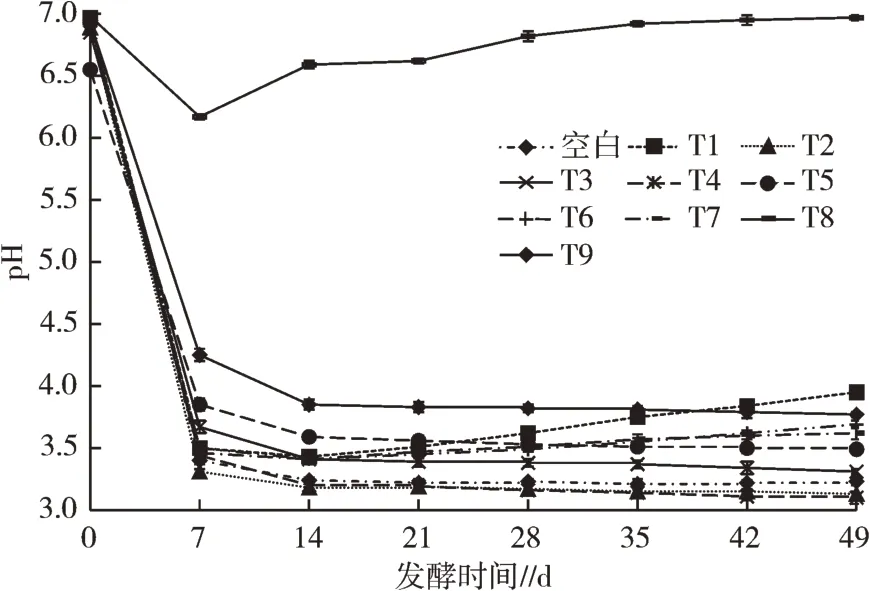
图1 发酵过程中pH变化
不同添加物对底物为蔬菜废弃物的发酵处理影响见图2,对比T5、T8、T9,发酵7 d后,T8的pH从6.17上升至6.59,而T5、T9的pH分别从3.85、4.25下降至3.59、3.85。分析发现,相同发酵底物(蔬菜废弃物)情况下,T8未添加红糖,发酵过程中pH显著高于其他处理,推测该发酵处理中EM菌剂与蔬菜发生反应生成的物质、环境抑制微生物的生长和产物的合成,导致其发酵效果不佳。
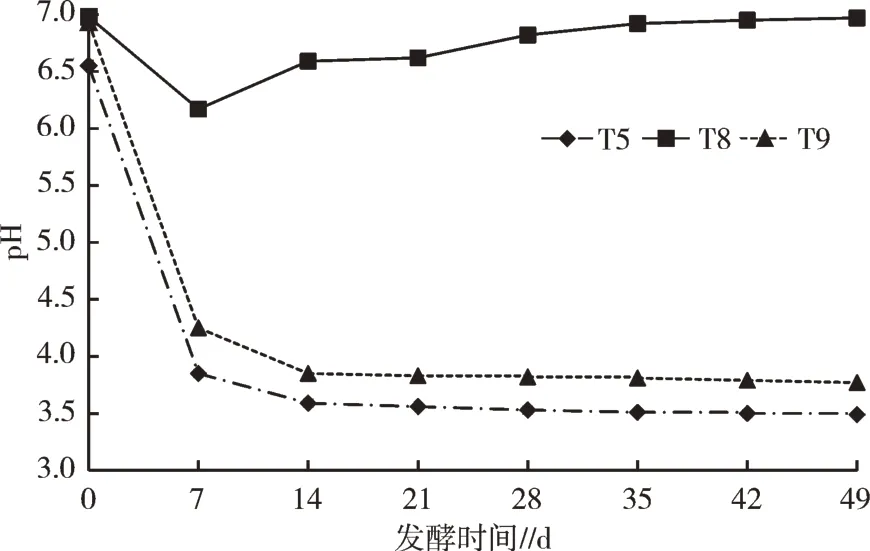
图2 添加物对pH的影响
发酵过程中酵素密度变化见图3,除空白处理缓慢下降外,不同发酵处理密度随时间变化均呈明显下降趋势,49 d时,T2、T3、T4、T5、T8、T9均明显低于1.000,推测果蔬尾菜酵素发酵过程中产生的物质密度小于1.000,可能产生乙醇等密度较小的发酵产物,说明这些发酵处理效果良好。

图3 发酵过程中酵素密度变化
2.2 酵素发酵过程中菌落总数变化
菌落总数是在一定条件下单位体积或者单位质量样品生长出的细菌、真菌、大肠杆菌等菌落总数。果蔬尾菜酵素制备时,可通过测定菌落数量表示其发酵程度。发酵过程中各处理组真菌菌落总数变化见图4,发酵过程各处理真菌总数不断上升,49 d发酵结束后,T2真菌菌落总数显著高于其余组处理,为3.25×105cfu/mL,较空白处理增幅为157.94%。说明料液比为3∶10,添加红糖(100 g/L)、EM菌剂(3 mL/L)时发酵效果最佳。49 d时,T1、T3真菌菌落总数分别较空白处理提升57.14%、30.95%,提升效果显著,说明添加EM菌剂或红糖均对发酵过程有显著促进作用,添加EM菌剂较添加红糖对发酵过程中真菌菌落总数的促进作用更为显著;T2、T4、T5真菌菌落总数分别为空白处理的257.94%,143.65%,77.46%,说明发酵底物为水果废弃物时发酵效果最佳;比较T1、T6、T7可知,发酵过程中真菌菌落总数与EM菌剂添加浓度呈负相关,推测EM菌剂添加浓度增大会抑制其他真菌的增长。同时添加红糖、EM菌剂(3 mL/L)的发酵处理(T2、T4、T5)为相同发酵底物真菌菌落总数最高的处理组,说明添加红糖、EM菌剂时发酵效果最佳。添加EM菌剂的发酵处理(T1、T6、T7、T8)酵素真菌菌落总数在35 d内快速上升,随后上升速度减缓,42~49 d维持稳定,菌落总数基本不变或略有下降,说明添加EM菌剂能够有效减少发酵时间,发酵至42 d时,各处理菌落总数基本稳定,为适宜发酵时长。添加红糖的发酵处理T3、T9在发酵35~49 d时较T1、T6、T7、T8菌落增速不断增加,菌落总数快速上涨,说明添加红糖能够有效增加发酵时间。
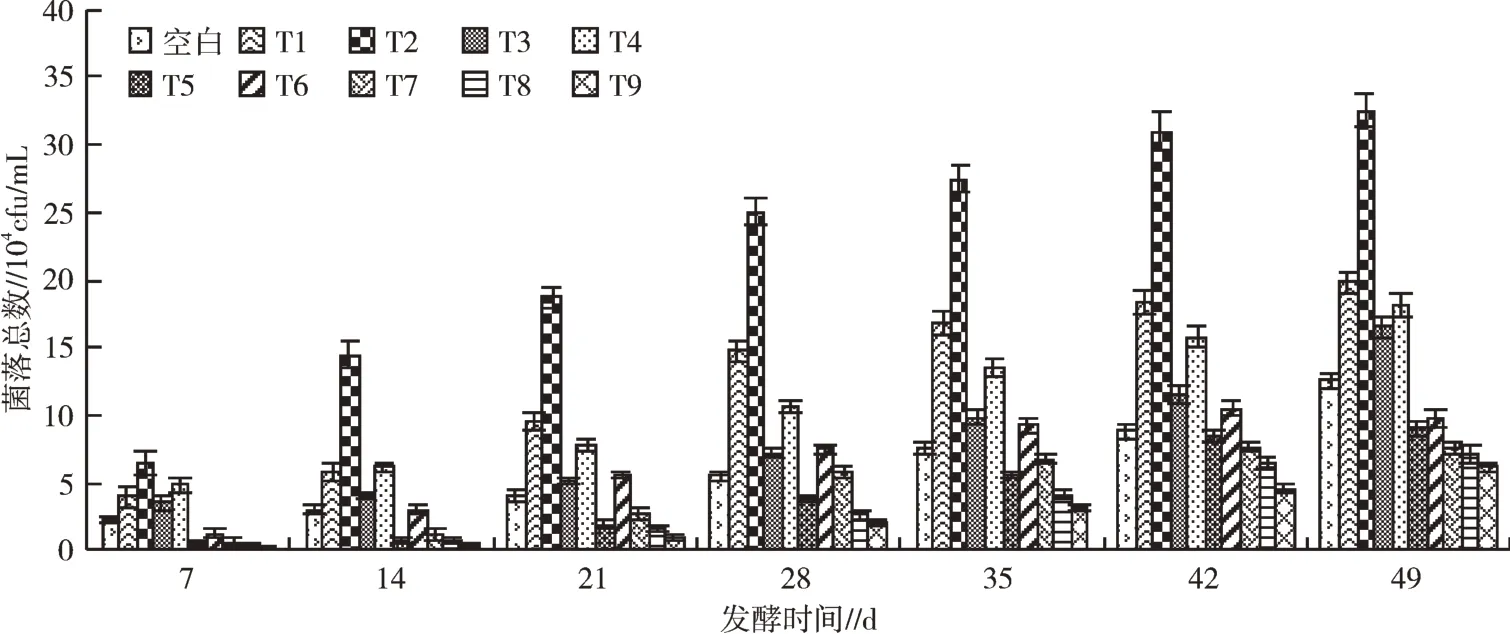
图4 发酵过程中真菌菌落变化
发酵过程中各处理细菌菌落总数变化见图5,果蔬尾菜酵素含有的细菌菌落总数较真菌菌落总数更多。添加EM菌剂的发酵处理42~49 d细菌菌落总数持续增加,无增速下缓趋势,与真菌菌落总数变化有差异。49 d发酵结束后,T2即添加红糖(100 g/L)和EM菌剂(3 ml/L)发酵效果最佳,细菌菌落总数较空白处理增幅为215.20%。T1、T3细菌菌落总数较空白处理分别提升61.01%、52.83%,提升效果显著;T2、T4、T5细菌菌落总数分别为空白处理的316.98%,137.74%,64.78%,说明发酵底物为水果废弃物时发酵效果最佳,细菌菌落总数为发酵底物为蔬菜的489.32%;比较T1、T6、T7可知,随着EM菌剂添加浓度的增加,发酵过程各时期细菌菌落总数呈下降趋势,推测EM菌剂添加浓度增大会抑制其他细菌的增长。
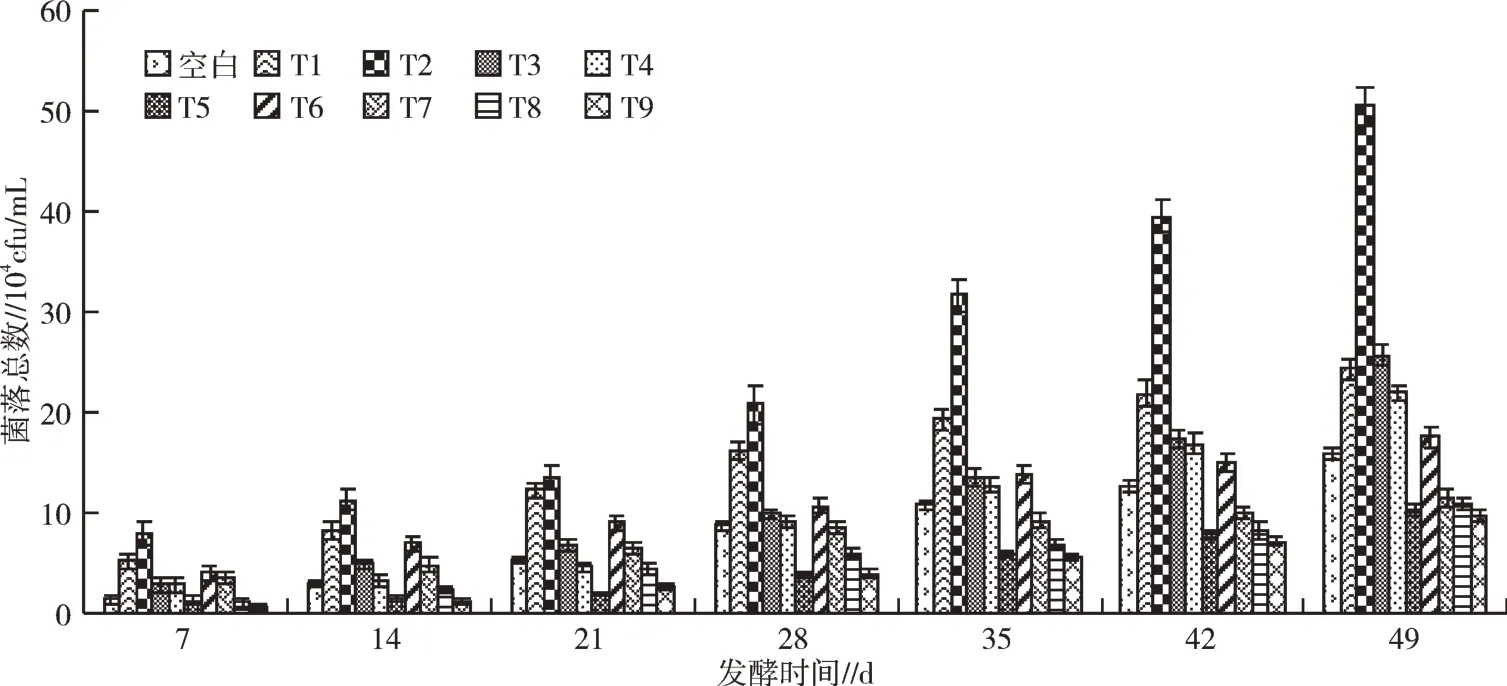
图5 发酵过程中细菌菌落变化
发酵过程中各处理大肠杆菌菌落总数变化见图6,除T8检测出含有大肠杆菌外,其余发酵处理均未检测出大肠杆菌,结合后续研究推测,因发酵过程中只有T8处理的pH为6.0~7.0,剩余9组处理pH均小于4.5,说明果蔬尾菜酵素环境绝大部分不利于大肠杆菌的生长。

图6 发酵过程中大肠杆菌菌落变化
2.3 酵素发酵过程中功效酶活性的变化
果蔬尾菜酵素中含有多种功效酶,随着发酵过程中功效酶酶活性不断上升,酵素作用效果不断提升。随着发酵不断进行,纤维素酶、脂肪酶、淀粉酶、蛋白酶活性均不断上升,发酵结束后各处理酶活性见表3。

表3 发酵结束后各处理的酶活性(单位:U/mL)
发酵过程中纤维素酶活性变化见图7,49 d发酵结束后,T2纤维素酶活性显著高于其余9组处理,为31.83 U/mL,较空白处理增幅为81.26%,说明T2发酵效果最佳。T1、T3发酵效果较好,较空白处理增幅分别为51.03%、54.67%,说明添加EM菌剂或红糖均对发酵过程有显著促进作用,且对于纤维素酶活性影响作用相似;T1、T6、T7纤维素酶活性与EM菌剂添加浓度呈负相关,推测EM菌剂添加浓度增大会抑制酶活性的增长;发酵底物为蔬菜废弃物的T5、T8、T9添加不同添加物后,发酵过程中酶活性均低于空白处理,纤维素酶活性仅为空白处理的84.00%、77.22%、55.30%,说明不同发酵底物种类对于酶活性的影响强于添加物对其影响,水果废弃物发酵优于蔬菜废弃物发酵。

图7 发酵过程中纤维素酶活性变化
发酵过程中脂肪酶活性变化见图8,49 d发酵结束后,T2脂肪酶活性显著高于其余组处理,为0.31 U/mL,较空白处理增幅为138.46%,说明T2发酵效果最佳。T1、T3发酵效果较好,较空白处理增幅分别为76.92%、130.77%,说明添加EM菌剂或红糖均对发酵过程有显著促进作用,添加EM菌剂较添加红糖对发酵过程中脂肪酶活性的促进作用更为显著;T1、T6、T7脂肪酶活性与EM菌剂添加浓度呈负相关,推测EM菌剂添加浓度增大会抑制酶活性的增长;酵底物为蔬菜废弃物的T5、T8、T9添加不同添加物后,发酵过程中酶活性均低于空白处理,脂肪酶酶活性仅为空白处理的92.30%、61.54%、69.23%,说明不同发酵底物种类对于酶活性的影响强于添加物对其影响,水果废弃物显著优于蔬菜废弃物发酵。但考虑到脂肪酶活性较低,仅为0.08~0.30 U/mL,测量精确度较低,可能导致产生误差,后续可进行更深入的研究,以期提高精确度。

图8 发酵过程中脂肪酶活性变化
发酵过程中α-淀粉酶、蛋白酶活性变化见图9、图10,49 d发酵结束后,T1至T9相较于空白处理,均无显著提升α-淀粉酶、蛋白酶活性,除T5、T8、T9外,不同处理α-淀粉酶活性均在3.0~4.0 U/mL,T1至T4处理较空白处理增幅均在20%以内,T5、T8、T9的α-淀粉酶活性显著低于空白处理,仅为空白处理的59.78%、34.92%、32.96%,说明不同发酵处理之间蛋白酶活性差距较小,蔬菜废弃物发酵不利于酵素蛋白酶活性增长。49 d发酵结束后,不同发酵组蛋白酶活性基本处于5.0~6.0 U/mL,T2处理较空白组增幅最大,也仅为17.1%,各处理组总体差距不大,说明不同发酵底物、不同添加物对于果蔬尾菜酵素α-淀粉酶及蛋白酶活性影响较小。

图9 发酵过程中α-淀粉酶活性变化

图10 发酵过程中蛋白酶活性变化
3 讨论
发酵过程中pH的变化可以反映发酵过程是否正常,同时也能反映酵素品质,是判断发酵成功与否的标志[21]。除T8外,49 d发酵结束时,剩余处理组pH均位于3.0~4.0并保持稳定,与蔡毅飞[21]、卜俊芝[22]的研究一致,说明发酵试验成功,果蔬尾菜酵素品质较好。发酵过程中7 d内酵素pH迅速下降,后续发酵过程中pH缓慢变化或保持稳定,说明发酵初期各项反应较为活跃,发酵速度较快,发酵中后期发酵环境基本保持稳定。发酵过程中未添加糖的发酵处理pH较高,符合张海燕等[23]的研究结果,说明酵素中糖添加量过低会影响微生物的生长和产物的合成。发酵过程中,酵素密度低于1.00,推测酵母菌发酵过程中产生醇类、醛类[24]、酸类[25]等物质,且随着醇类、醛类、酸类等物质浓度的增加,酵素密度不断降低[26]。
菌落总数为微生物指示性指标之一,可在一定程度上反映生产过程中的发酵程度。本研究通过试验发现,发酵结束后不同处理尾菜酵素中真菌、细菌菌落总数含量普遍达到7×105cfu/mL以上,该结论与文浩等[25]、何曼等[27]的研究结果相似,说明试验果蔬尾菜酵素发酵良好。添加EM菌剂处理组细菌、真菌菌落显著高于其他处理组,与位伯华[28]的研究结果一致,说明尾菜酵素添加EM菌剂可显著增加真菌、细菌菌落总数,发酵程度良好。
酵素通常因含有多种功效酶可有效进行资源化利用,本研究对酵素的纤维素酶、脂肪酶、蛋白酶及α-淀粉酶的酶活性进行测定,分析说明酵素中功效酶含量较高,与饶毅萍等[29]的研究一致。4种功效酶的含量差距较大,其中纤维素酶相对活力最高,脂肪酶相对活力最低,与周新萍等[19]、张海燕等[23]和陈爽等[30]的研究结果相似,而蛋白酶酶活力较低,与蔡毅飞[21]的研究结果蛋白酶的相对活性最高不一致,推测可能与试验采用的原料不同且试验所选用原料相对单一有关。
4 结论
果蔬尾菜酵素具有相对稳定的微生物生态环境,但不同发酵阶段中,微生物的种类和数量发生变化,各种反应和微生物之间的相互影响,导致各项指标在一定区间内动态变化。绝大部分数据显示,在发酵底物为水果废弃物,料液比为3∶10,添加红糖(100 g/L)与EM菌剂(3 ml/L)时发酵效果最佳,菌落总数、pH密度等多项指标较空白发酵组均有显著提升。对果蔬尾菜酵素发酵过程中功效酶活性(纤维素酶、蛋白酶、淀粉酶、脂肪酶)的变化进行研究,结果表明,酵素发酵过程中,在不同因素影响下,酵素密度不断下降,pH在一定区间内保持稳定变化;菌落总数不断上升,含有一定数量的真菌、细菌,大部分不含大肠杆菌;功效酶活性不断增加,纤维素酶活性较高,含有一定数量的α-淀粉酶及蛋白酶,脂肪酶酶活性较低。在发酵底物不同的情况下,酵素的微生物数量,酶活性存在较大差异。
猜你喜欢
娃娃乐园·综合智能(2022年3期)2022-04-19
农技服务(2021年6期)2021-12-05
今日农业(2021年16期)2021-11-26
农业技术与装备(2021年8期)2021-10-14
中老年保健(2021年10期)2021-08-24
祝您健康·文摘版(2017年5期)2017-04-06
中国蔬菜(2016年8期)2017-01-15
凤凰资讯报(2016年5期)2016-05-30
河北农业(2016年1期)2016-05-19
恋爱婚姻家庭·青春(2016年4期)2016-04-07
