
半枝莲提取物对肝癌HuH-7细胞自噬的影响及其作用机制
2022-12-28 08:00王娟霞鲁佳敏吴瑞郭佳敏门碧莹黄子龙
中国生物制品学杂志 2022年12期
王娟霞,鲁佳敏,吴瑞,郭佳敏,门碧莹,黄子龙
1.兰州大学第二医院感染科,甘肃 兰州 730030;2.兰州大学第二临床医学院,甘肃 兰州 730000
目前,肝癌是全球第六大恶性肿瘤,也是全球第三大癌症相关死亡原因[1]。尽管肝癌的诊断和治疗方法有了较大改进,但仅有10% ~20%的患者诊断为早期肝癌,导致肝癌仍难以治愈[2]。因此,进一步改进肝癌的诊疗方法和探索新的药物对于提高肝癌的治疗效果均具有重要意义。
半枝莲又名韩信草,是一种唇形目唇形科植物,全草入药,味辛性平,无毒,具有清热解毒、散瘀止痛、利湿等功能,主含黄酮类、二萜类和多糖等成分。研究表明,半枝莲与其他肿瘤联合组方可治疗肺癌、宫颈癌和肝癌等多种肿瘤[3]。同时,半枝莲单药提取物已被证明可显著抑制多种肿瘤细胞的增殖。如半枝莲提取物可通过诱导凋亡来抑制人结肠癌细胞增殖[4-5]。WONG 等[6]研究显示,半枝莲提取物可诱导前列腺癌细胞凋亡并抑制细胞存活。此外,半枝莲提取物可明显诱导肝癌细胞凋亡和细胞周期阻滞,从而抑制细胞增殖[7-8]。但其抑制肝癌生长的作用机制目前尚不清楚。
自噬是真核生物对细胞内物质进行周转的重要方式,通过这一途径,细胞内受损的蛋白质和细胞器会被囊泡吞噬,并与溶酶体融合形成自噬溶酶体,从而降解内容物以维持细胞本身的代谢需要和细胞器更新[9]。在自噬发生的初始阶段,错误折叠蛋白或待降解物与P62 蛋白和上游Atg 蛋白相互结合,随后LC3B 被募集至该结构周围形成吞噬泡,随着双层膜结构的不断延伸,P62 蛋白、待降解底物与LC3B 和Atg 蛋白结合形成自噬体[10-12]。研究表明,自噬在包括肝癌在内的多种肝脏相关疾病的发病和进展过程中发挥重要作用[13]。在黑色素瘤研究中,半枝莲提取物还可通过激活自噬抑制肿瘤的生长[14]。提示自噬可能在半枝莲提取物诱导肝癌细胞生长抑制中发挥重要作用。因此,本研究首先通过细胞活力试验确定半枝莲提取物的作用浓度后,将其作用于HuH-7细胞,以评价对自噬的影响,然后通过3-Methyla-denine(3-MA)抑制自噬,探讨自噬在半枝莲提取物影响HuH-7细胞生长中的作用,并进一步研究半枝莲提取物对HuH-7 细胞PI3K/AKT/mTOR 信号通路活性的影响,从而阐明半枝莲提取物调节自噬的机制。
1 材料与方法
1.1 细胞 HuH-7 细胞购自中国科学院典型培养物保藏委员会细胞库。
1.2 主要试剂及仪器 半枝莲提取物购自北京沃凯生物科技有限公司;3-MA购自美国MedChemExpress公司;DMEM购自美国Invitrogen公司;胎牛血清购自美国Gibco 公司;CCK-8 试剂盒购自上海生博生物医药科技有限公司;山羊封闭血清和RIPA 裂解液购自武汉博士德生物工程有限公司;兔抗人LC3B、P62、GAPDH、p-AKT、AKT、mTOR 和p-mTOR 单克隆抗体购自美国Abcam公司;Annexin V-FITC/PI 凋亡检测试剂盒和DAPI 染色液购自上海碧云天生物技术有限公司;山羊抗兔Alexa 荧光二抗购自美国Thermo-Fisher 公司;酶标仪购自北京普朗新技术有限公司;流式细胞仪购自美国BD公司。
1.3 细胞培养及分组 将HuH-7 细胞用含10%胎牛血清的DMEM培养基,于37 ℃,95%空气和5%CO2培养箱中培养,每3 d换液1次。收集对数生长期HuH-7 细胞,按5 × 104个/mL 的细胞密度接种96 孔板,100 μL/孔,培养箱培养24 h;更换成含100、200、500、1 000和1 500 μg/mL半枝莲提取物以及500 μg/mL半枝莲提取物+ 5 mmol/L 3-MA 的DMEM 培养基继续培养24 h,设不含半枝莲提取物对照组。
1.4 CCK-8试验 收集各组细胞,加入CCK-8溶液,10 μL/孔,培养箱培养2 h,上酶标仪检测A450值,并按下式计算细胞活力。试验重复3次。
细胞活力(%)=(加药组A450值-空白组A450值)/(对照组A450值-空白组A450值)×100%
1.5 激光共聚焦试验 收集各组细胞,弃培养基,PBS 洗涤3 次,4%多聚甲醛固定20 min;0.1%Triton X-100透化10 min;PBS洗涤3次,用10%山羊封闭血清处理30 min;加入兔抗LC3B 单克隆抗体(1∶300稀释),4 ℃孵育过夜;PBS 洗涤3 次,加入山羊抗兔Alexa 荧光二抗(1∶500 稀释),37 ℃孵育2 h;PBS 洗涤3 次,加入DAPI 染色液,室温避光孵育5 min;PBS洗涤3次,滴加抗荧光淬灭剂于玻片上,盖上盖玻片,于激光共聚焦显微镜下观察并拍照。试验重复3次。
1.6 Western blot分析 收集各组细胞,预冷PBS洗涤后,预冷RIPA裂解液处理细胞,16 000×g离心20 min,BCA 蛋白定量试剂盒测定蛋白浓度。按30 μg/孔蛋白上样12%SDS-PAGE凝胶孔中,100 V电泳1.5 h;转至PVDF 膜上,5%封闭液室温封膜1 h;加入相应的兔抗人LC3B(1∶2 000稀释)、P62(1∶1 000稀释)、GAPDH(1∶5 000稀释)、p-AKT(1∶5 000稀释)、AKT(1∶5 000 稀释)、mTOR(1∶5 000 稀释)和p-mTOR(1∶5 000 稀释)单克隆抗体,4 ℃孵育过夜;TBS 洗涤3 次,加入山羊抗兔Alexa 荧光二抗(1∶10 000 稀释),室温孵育1 h;TBS 洗涤3 次,化学发光显色。采用Image J 软件分析目的条带灰度值,并计算目的蛋白相对表达量。试验重复3次。
1.7 细胞凋亡检测 采用流式细胞术。收集各组细胞,PBS洗涤1次,用0.25%胰酶消化,1 000×g离心5 min;PBS 重悬后,进行细胞计数,每组取含5 × 104个细胞的悬液,1 000 ×g离心5 min;加入195 μL Annexin V-FITC 结合液重悬,加入5 μL Annexin VFITC 结合液混匀,加入10 μL PI染色液混匀,室温避光孵育15 min;上流式细胞仪检测各组细胞凋亡情况。试验重复3次。
1.8 统计学分析 采用IBM SPSS 19.0 软件进行统计学分析,试验数据以均数±标准差(x±s)表示,多组独立样本比较采用单因素方差分析(One-Way ANOVA),两组间比较采用Tukey检验,以P<0.05 为差异有统计学意义。
2 结果
2.1 不同浓度半枝莲提取物对HuH-7 细胞活力的影响 结果显示,与对照组相比,100、200、500、1 000和1 500 μg/mL 半枝莲提取物均可明显抑制HuH-7细胞活力(F分别为0.937、0.000、0.036、0.945、3.821,P均<0.01),随着药物浓度的增加,细胞活力逐渐降低,见图1。500 μg/mL浓度的半枝莲提取物抑制HuH-7 细胞活性40%左右,作用浓度适中,因此,选择该浓度处理HuH-7细胞,进行后续试验。

图1 半枝莲提取物对HuH-7细胞活力的影响Fig.1 Effect of Scutellaria barbata extract on activity of HuH-7 cells
2.2 半枝莲提取物对HuH-7细胞自噬的影响 结果显示,与对照组[(11.67±1.53)个]相比,500 μg/mL半枝莲提取物组细胞LC3B斑点数[(31.00±3.61)个]明显升高(F= 2.597,P<0.01),见图2。Western blot 分析显示,对照组和500 μg/mL 半枝莲提取物组细胞LC3B-Ⅱ/LC3B-Ⅰ值分别为(0.10 ± 0.04)和(2.24 ± 0.11);与对照组相比,500 μg/mL 半枝莲提取物组细胞LC3B-Ⅱ蛋白表达明显升高(F=2.127,P<0.01)。对照组和500 μg/mL 半枝莲提取物组细胞P62蛋白相对表达量分别为(0.85±0.07)和(0.15±0.04);与对照组相比,500 μg/mL 半枝莲提取物组P62 蛋白相对表达量明显降低(F=0.859,P<0.01)。见图3。

图2 激光共聚焦试验检测各组细胞LC3B斑点数Fig.2 Determination of acounts of LC3B spots of cells in various groups by laser scanning confocal microscopy
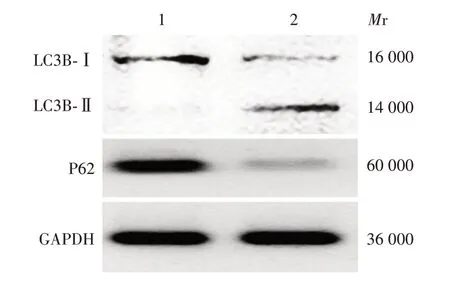
图3 Western blot 分析各组细胞中LC3B-Ⅰ/Ⅱ和P62 蛋白的表达Fig.3 Western blotting of expression of LC3B-Ⅰ/Ⅱand P62 proteins in cells of various groups
2.3 3-MA 逆转半枝莲提取物对HuH-7 细胞自噬的影响 结果显示,对照组、500 μg/mL半枝莲提取物组、500 μg/mL 半枝莲提取物+5 mmol/L 3-MA 组细胞LC3B斑点数分别为(9.67±5.03)、(36.33±4.04)和(11.33±2.01)个。与对照组相比,500 μg/mL半枝莲提取物组细胞LC3B斑点数明显升高(F=0.131,P<0.01);与500 μg/mL 半枝莲提取物组相比,500 μg/mL半枝莲提取物+5 mmol/L 3-MA组细胞LC3B斑点数明显降低(F=1.180,P<0.01)。见图4。Western blot 分析显示,对照组、500 μg/mL 半枝莲提取物组、500 μg/mL 半枝莲提取物+ 5 mmol/L 3-MA 组细胞LC3B-Ⅱ/LC3B-Ⅰ值分别为(0.13 ±0.04)和(2.45 ± 0.09)和(0.89 ± 0.17)。与对照组相比,500 μg/mL 半枝莲提取物组细胞LC3B-Ⅱ蛋白表达明显升高(F=3.273,P<0.01);与500 μg/mL半枝莲提取物组相比,500 μg/mL 半枝莲提取物+5 mmol/L 3-MA 组细胞LC3B-Ⅱ蛋白表达明显降低(F=2.315,P<0.01)。对照组、500 μg/mL半枝莲提取物组、500 μg/mL半枝莲提取物+5 mmol/L 3-MA组细胞P62蛋白相对表达量分别为(0.84±0.10)、(0.11 ± 0.05)和(0.53 ± 0.05)。与对照组相比,500 μg/mL半枝莲提取物组细胞P62蛋白相对表达量明显降低(F= 1.892,P<0.01);与500 μg/mL半枝莲提取物组相比,500 μg/mL 半枝莲提取物+5 mmol/L 3-MA 组细胞P62 蛋白相对表达量明显升高(F=0.153,P<0.01)。见图5。

图4 各组细胞LC3B斑点数的激光共聚焦显微镜观察Fig.4 Laser scanning confocal microscopy of acounts of LC3B spots of cells in various groups

图5 各组细胞中LC3B-Ⅰ/Ⅱ和P62蛋白表达的Western blot分析Fig.5 Western blotting of expression of LC3B-Ⅰ/Ⅱand P62 proteins in cells of various groups
2.4 3-MA 逆转半枝莲提取物对HuH-7 细胞活力的影响 结果显示,与对照组相比,500 μg/mL半枝莲提取物组细胞活力明显降低(F=3.231,P<0.01);与500 μg/mL半枝莲提取物组相比,500 μg/mL半枝莲提取物+5 mmol/L 3-MA组细胞活力明显升高(F=1.462,P<0.01)。见图6。

图6 3-MA逆转半枝莲提取物对HuH-7细胞活力的影响Fig.6 Effect of 3-MA reverse Scutellaria barbata extract on activity of HuH-7 cells
2.5 3-MA 逆转半枝莲提取物对HuH-7 细胞凋亡的影响 结果显示,对照组、500 μg/mL半枝莲提取物组、500 μg/mL 半枝莲提取物+5 mmol/L 3-MA 组细胞凋亡率分别为(9.49±0.94)%、(25.68±2.08)%和(13.21±1.60)%。与对照组相比,500 μg/mL半枝莲提取物组细胞凋亡比率明显升高(F= 1.284,P<0.01);与500 μg/mL 半枝莲提取物组相比,500 μg/mL半枝莲提取物+5 mmol/L 3-MA组细胞凋亡比率明显降低(F=0.119,P<0.01)。见图7。
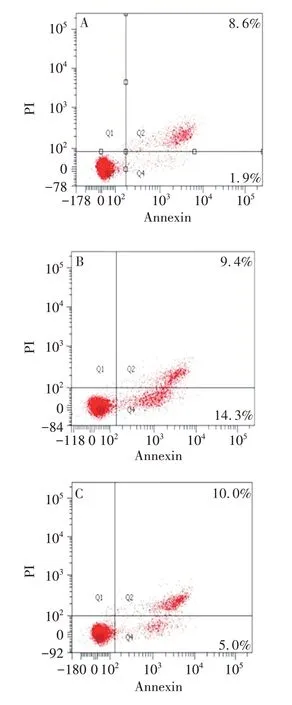
图7 3-MA逆转半枝莲提取物对HuH-7细胞凋亡的影响Fig.7 Effect of 3-MA reverse Scutellaria barbata extract on apoptosis of HuH-7 cells
2.6 半枝莲提取物对HuH-7细胞PI3K/AKT/mTOR信号通路的影响 结果显示,对照组细胞p-AKT/AKT 和p-mTOR/mTOR 值分别为(0.92 ± 0.04)和(0.56±0.06),而500 μg/mL 半枝莲提取物组细胞p-AKT/AKT 和p-mTOR/mTOR 值分别为(0.23 ±0.06)和(0.24±0.03)。与对照组细胞相比,500 μg/mL半枝莲提取物组细胞p-AKT 和p-mTOR 蛋白表达明显降低(F分别为0.945 和0.955,P均<0.01)。见图8。

图8 Western blot 分析半枝莲提取物对PI3K/AKT/mTOR信号通路的影响Fig.8 Analysis of effect of Scutellaria barbata extract on PI3K/AKT/mTOR signal pathway by Western blotting
3 讨论
近年来的研究表明,半枝莲具有抗炎、抗病毒、抗氧化和抗癌的作用[15]。DAI 等[16]研究表明,半枝莲提取物能明显抑制肝癌MHCC97H细胞的增殖、侵袭和转移。本研究通过CCK-8 试验也发现,半枝莲提取物可明显抑制肝癌HuH-7 细胞活力,并呈浓度依赖性。也有研究的动物实验结果表明,半枝莲提取物可明显抑制荷实体肝癌H22 小鼠肿瘤生长[17]。因此,半枝莲提取物对肝癌细胞具有重要的抑制作用,有望开发成为有效的抗肝癌药物。
自噬作为真核细胞溶酶体的主要降解途径之一,在细胞内分子和细胞器循环利用中发挥重要作用。近年来的研究表明,自噬在肿瘤中的作用比较复杂,在癌症形成早期阶段,自噬作为一种抗癌途径阻止正常细胞向癌细胞的恶性转化;另一方面,自噬参与了肿瘤进展和转移的整个过程[18]。有研究表明,Becn1缺失加速乙型肝炎病毒(hepatitis B virus,HBV)相关的肝细胞癌形成,证明了自噬基因Becn1在维持肝稳态和抑制肝癌形成中的重要性[19]。相反,在肝癌中,自噬可促进细胞生长,并与肿瘤的转移和耐药性相关[20]。因此,自噬发挥双重作用,在不同环境中发挥促进或抑制肿瘤生长和发展的作用。有研究表明,半枝莲总黄酮可抑制荷瘤小鼠黑色素瘤体积,并诱导肿瘤细胞凋亡和自噬,抑制PI3K/AKT/mTOR信号通路活性[10]。本研究也发现,半枝莲提取物能够增强肝癌细胞自噬。但自噬在半枝莲提取物抑制肝癌细胞生长中的作用尚不清楚。鉴于文献报道半枝莲提取物对PI3K/AKT/mTOR 信号通路活性的抑制作用[14,21-24],本研究通过3-MA(一种PI3K 抑制剂)抑制半枝莲提取物诱导的自噬增强作用,结果显示,自噬抑制后,半枝莲提取物对肝癌细胞活性的抑制以及凋亡的诱导能力均明显减弱,同时,半枝莲提取物明显抑制肝癌细胞中PI3K/AKT/mTOR 信号通路活性。因此,本研究结果表明,半枝莲提取物可通过抑制PI3K/AKT/mTOR 信号通路活性来诱导自噬,从而降低肝癌细胞的生长能力。这与自噬可促进肝癌进展的报道存在差异。但WANG等[25]研究发现,H2S 可通过调节PI3K/AKT/mTOR信号通路增强自噬来抑制肝癌细胞增殖、侵袭和细胞分裂。该报道与本研究结果一致,进一步证明了自噬的双重作用。
综上所述,半枝莲提取物可增强肝癌细胞自噬,而自噬的增加会导致细胞生长受到抑制,其机制可能与对PI3K/AKT/mTOR信号通路的抑制相关。
猜你喜欢
中老年保健(2022年2期)2022-08-24
中老年保健(2021年5期)2021-12-02
中老年保健(2021年12期)2021-11-30
昆明医科大学学报(2021年4期)2021-07-23
军民两用技术与产品(2021年10期)2021-03-16
中西医结合肝病杂志(2020年2期)2020-10-27
当代陕西(2020年9期)2020-08-04
癌变·畸变·突变(2015年4期)2015-02-27
中国民族民间医药·下半月(2014年2期)2014-09-26
浙江人大(2014年1期)2014-03-20
