
基于线粒体CO I 和Cyt b 序列的丹江口水库鳙群体遗传结构分析
2022-12-27 04:12汪鄂洲付昕石义付陈泽涛朱勇夫董薇薇郑海涛4阙延福李伟涛朱丽娅
渔业致富指南 2022年11期
○汪鄂洲付昕石义付陈泽涛朱勇夫董薇薇郑海涛,4阙延福李伟涛朱丽娅
(1.水利部水工程生态效应与生态修复重点实验室,水利部中国科学院水工程生态研究所 湖北武汉 430079;2.南水北调中线水源有限责任公司 湖北丹江口 442700;3.湖北省水产科学研究所 湖北武汉 430071;4.湖北中水长江生态保护研究院有限公司 湖北武汉 430014)
鳙(Aristichthys nobilis)属鲤形目(Cypriniformes)、鲤科(Cprinidae)、鲢亚科(Hypophthalmichthyinae),是我国传统的“四大家鱼”之一,其自然分布区主要在中国大陆长江和珠江流域,其他水体中也有分布。自二十世纪五十年代我国突破四大家鱼人工繁殖技术后,鳙逐渐成为我国乃至世界主要淡水鱼类养殖种类。世界上已有20多个国家和地区引进了鲢和鳙,或作为水产养殖对象供人们食用,或用于池塘、水库等水体浮游生物的生物控制。根据联合国粮农组织(FAO)的统计资料,2010年鳙为世界第七大水产养殖种类,根据我国2019年渔业年鉴统计数据,鳙的产量达到310.16万吨,居养殖产量第三位。由于受水利建设,过度捕捞以及水体污染等多种因素影响,我国几大天然水体中的鳙野生资源量呈急剧下降态势。
丹江口水库(110°34′47″-110°47′53″E,32°14′10″-32°58′10″N)位于汉江中上游,在湖北省丹江口市和河南省南阳市之间,水域横跨鄂、豫两省,是南水北调中线工程的唯一水源地,也是优化我国水资源配置的重大战略工程。为进一步改善水库水质,维护和恢复丹江口水库水生生物资源,近20年来丹江口库区多次开展包括“四大家鱼”在内的经济鱼类增殖放流活动。仅2019年丹江口市放流鲢、鳙、草鱼、鳊等鱼苗4250万尾。虽然大规模的增殖放流一定程度上对鳙资源的恢复起到作用,但放流苗种来源不一在很大程度上会对天然水域鳙群体带来影响,甚至会导致物种野生群体遗传多样性降低,造成遗传污染。因此,应加强增殖放流的遗传管理,在放流前开展遗传评估。冯晓婷等对2016-2018年长江江苏段主要参与增殖放流的良种场所采集的鳙亲鱼和长江江苏段各采样点的回捕鳙进行评估,结果显示增殖放流对长江江苏段野生群体的贡献率为6.11%。
遗传多样性不仅是物种多样性和生态系统多样性的基础,也是鱼类物种生存与进化的基础,开展鱼类遗传多样性研究,对渔业资源管理与养护、人工增殖放流等工作具有重要的指导意义。自上世纪九十年代以来,我国科研人员利用同工酶、微卫星、线粒体等不同的标记对长江流域鳙遗传多样性进行了研究,而有关丹江口库区鱼类的遗传多样性研究尚未见报道。丹江口库区有关鱼类的研究主要集中在库区鱼产力、鱼类资源调查及鱼类群落特征等方面。
本研究利用线粒体CO I和Cytb序列分析了丹江口水库鳙增殖放流亲本、苗种及库区自然群体的遗传多样性,研究结果不仅是科学的渔业管理、制定保护策略依据,也将为鳙种质资源开发利用和增殖放流遗传管理提供基础支撑。
1 材料与方法
1.1 实验材料
2019-2020年于丹江口增殖放流站随机采集鳙亲本(Yong-QB)和子代(Yong-ZD),收集丹江口库区自然水体鳙样品(Yong-KQ),共计收集鳙样品92尾,具体信息见表1。每个样品经过形态鉴定后,剪取适量尾鳍置于装有无水乙醇的EP管中,带回实验室冻存于-20℃冰箱保存备用。
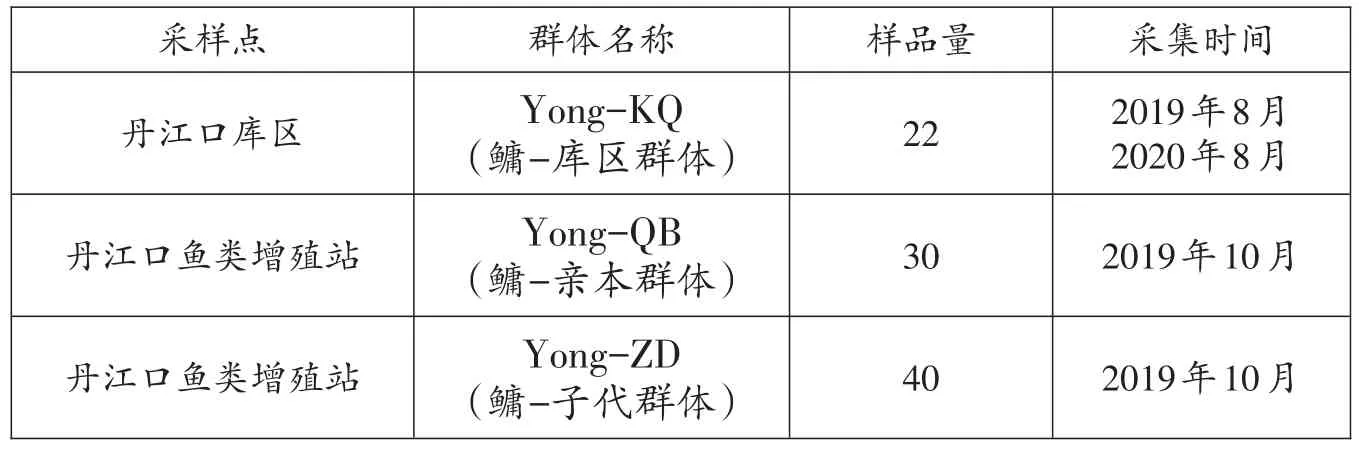
表1 鳙样本信息
1.2 基因组总DNA提取及PCR扩增
鳙基因组总DNA采用酚/氯仿法提取,溶解于200 μL TE缓冲液中,检查DNA样品浓度后,保存于-20℃冰箱备用。DNA工作液稀释为 50 ng/μL,PCR 反应体系为40 μL,其中Mix 20 μL、正反向引物各 2 μL、双蒸水 14 μL、DNA模板2 μL。PCR扩增条件如下:94℃预变性 5 min,94℃变性 35 s,58℃退火 35 s,72℃延伸1 min,35个循环;72℃终延伸10 min。PCR产物送天一辉远生物科技有限公司测序。鳙线粒体PCR扩增引物见表2。
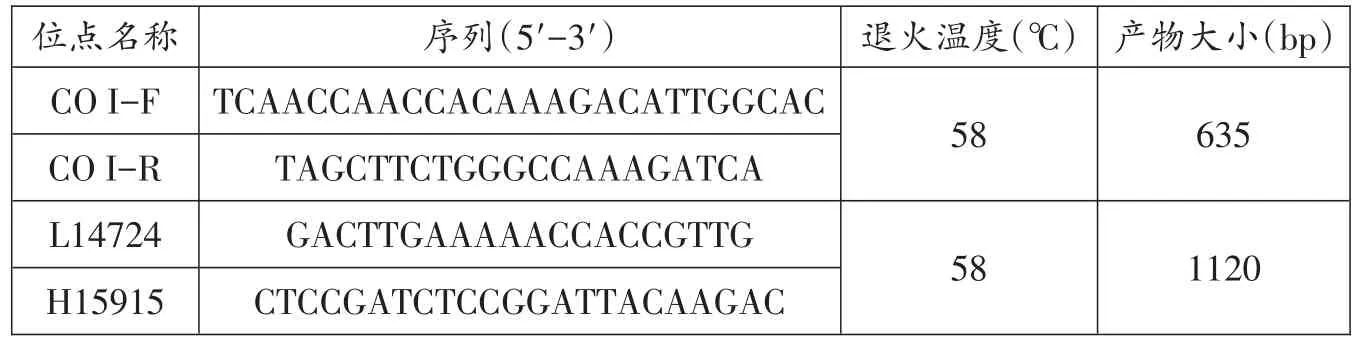
表2 鳙CO I和Cytb序列扩增引物信息
1.3 数据处理
利用软件FinchTV对测序得到的CO I和Cytb序列进行拼接,辅以人工校对,并与NCBI数据库中已发表的鳙mtDNA序列进行对比,确保扩增CO I和Cytb序列的准确性。使用DnaSP 5.0软件计算单倍型数目(N)、变异位点(V)、单倍型多样性(Hd)、核苷酸多样性(Pi)、平均核苷酸差异数(K)、基因流(Nm),以及单倍型类型等参数。使用Arlequin 3.01软件,根据pairwise difference模型计算种群间的分化指数值(Fst)和分子方差分析(AMOVA)。使用MEGA V6.0计算碱基含量和转换/颠换比率,根据Kimura双参数模型计算单倍型间遗传距离并进行NJ聚类。
2 结果
2.1 碱基组成
测序后经序列比对,共获得3个鳙群体92条有效的线粒体CO I和92条有效的Cytb序列,长度分别为653 bp和1120 bp。基于CO I基因序列分析结果表明,碱基的平均含量分别为A=25.5%,T=29.3%,C=27.3%,G=17.9%;其中A+T的平均含量为54.8%,G+C为45.2%。92条鳙线粒体CO I序列中检测到多态位点3个,其中简约信息位点3个,单一变异位点0个,定义了4个单倍型,其中1个单倍型为3个群体共有单倍型,2个单倍型为2个群体共有,1个单倍型仅在亲本群体中检测到(表3)。
基于Cytb基因序列分析结果表明,碱基的平均含量分别为A=29.6%,T=27.2%,C=29.0%,G=14.2%,转换颠换比为6.4;其中A+T的平均含量为56.8%,G+C为43.2%。92条鳙线粒体Cytb序列检测到多态位点9个,其中简约信息位点6个,单一变异位点3个,定义了8个单倍型,其中1个单倍型为3个群体共有单倍型,3个单倍型为2个群体共有,4个单倍型仅在单一群体中检测到(表3)。
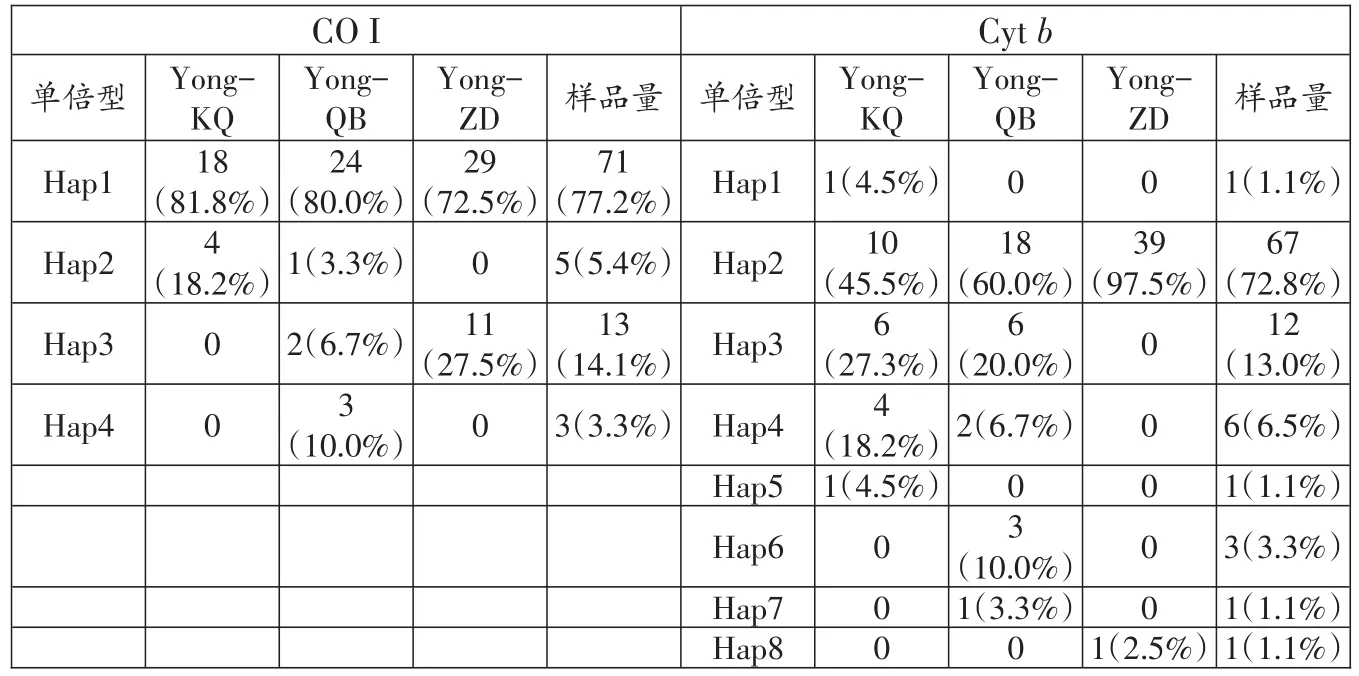
表3 鳙3个群体单倍型数目、类型、频率及分布
2.2 群体遗传多样性
线粒体CO I序列分析结果显示,3个鳙群体共定义了4个单倍型,其中Yong-KQ群体共22尾,定义了2个单倍型,其单倍型多样性最低,仅为(0.312±0.106);Yong-QB群体共30尾,定义了4个单倍型,其单倍型多样性为(0.356±0.106);Yong-ZD群体共40尾,定义了2个单倍型,其单倍型多样性最高,为(0.409±0.065)。核苷酸多样性和平均核苷酸差异分析结果显示,Yong-ZD群体最高,分别为(0.00063±0.00011)、(0.40897);Yong-QB 群体次之,分别为(0.00059±0.00019)、(0.38161);Yong-KQ 群体最低,分别为(0.00048±0.00016)、(0.31169)。
线粒体Cytb序列分析结果显示,3个鳙群体共定义了8个单倍型,其中Yong-KQ群体共22尾,定义了5个单倍型,其单倍型多样性最高,为(0.714±0.065);Yong-QB群体共30尾,定义了5个单倍型,其单倍型多样性为(0.605±0.016);Yong-ZD群体共40尾,定义了2个单倍型,其单倍型多样性最低,仅有(0.050±0.047),此群体中有1个特有单倍型,可能是在丹江口增殖站运行初期,引进了部分苗种所导致。核苷酸多样性和平均核苷酸差异分析结果显示,Yong-ZD群体最低,分别为(0.00004±0.00004)、(0.05000);Yong-QB群体次之,分别 为(0.00158± 0.00027)、(1.76782);Yong-KQ群体最高,分别为(0.00159± 0.00022)、(1.78355)(表4)。
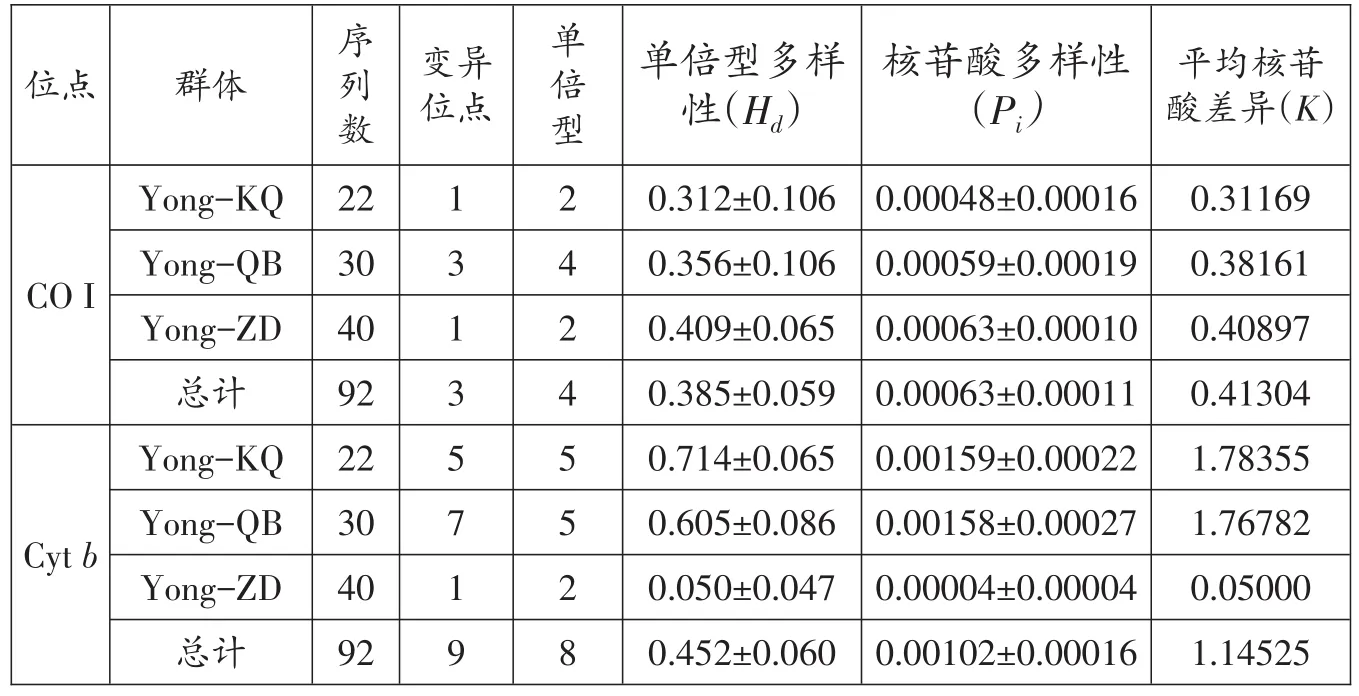
表4 鳙群体遗传多样性参数
2.3 群体遗传差异
根据线粒体CO I和Cytb序列,采用Kimura双参数(K2-P)模型分析鳙群体内和群体间遗传距离,结果显示群体内和群体间的遗传距离差异不明显(0.00048-0.00070,CO I;0.00004-0.00160,Cytb),群体内和群体间遗传距离小(表5)。遗传分化指数Fst和基因流Nm结果显示,鳙群体间的Fst在 0.06785~0.20484,Yong-ZD和Yong-QB群体间P值小于0.05,Yong-KQ和Yong-ZD群体间P值小于 0.001,Nm在 4.29~30.26(基于CO I序列);Cytb分析结果显示,Fst在 -0.00851~0.31930,Yong-ZD 和 Yong-KQ、Yong-QB群体间P值小于0.001,Nm在 0.89~186.66(表 6)。CO I分子方差分析(AMOVA)结果显示,群体间的变异贡献率占整个变异的12.60%,而群体内的变异贡献率占87.40%;Cytb分析结果显示,群体间的变异贡献率占整个变异的15.94%,而群体内的变异贡献率占84.06%(表7)。
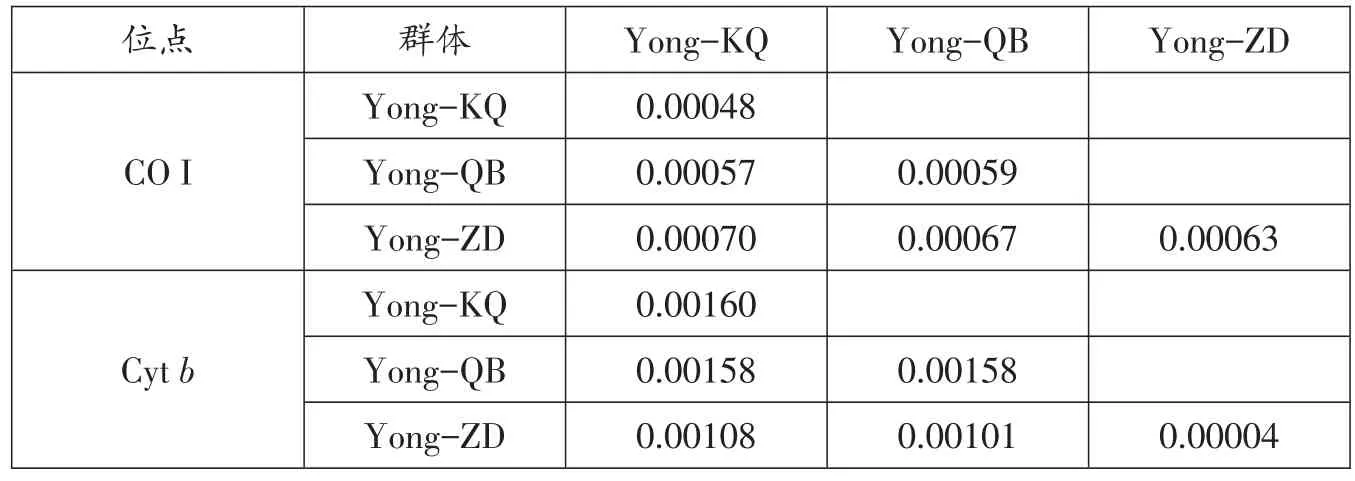
表5 鳙群体内和群体间的遗传距离
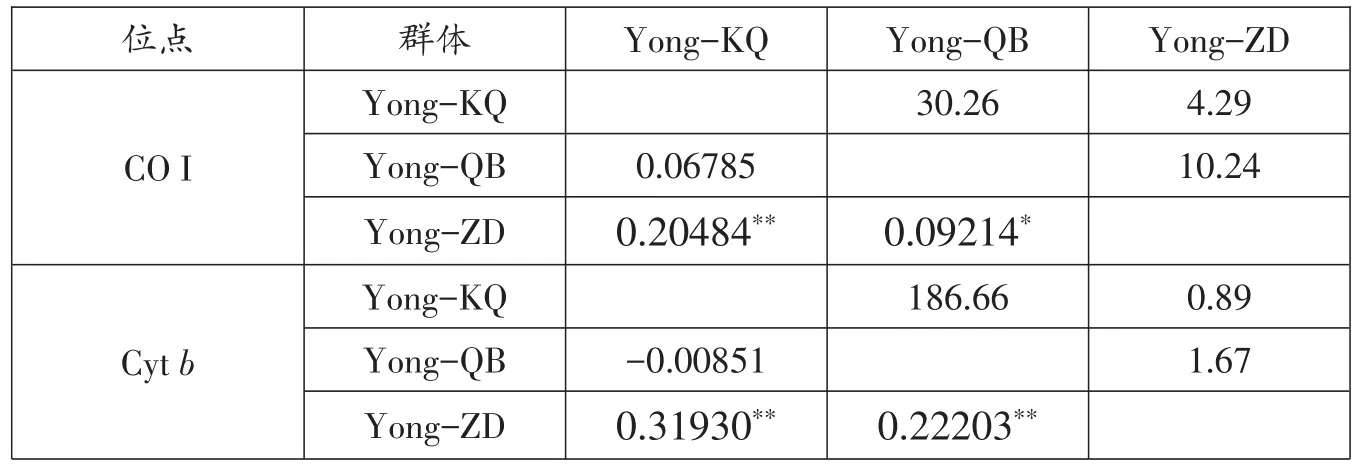
表6 鳙群体间的遗传分化系数和基因流

表7 鳙群体的分子方差分析
2.4 系统进化关系
基于 Kimura(K2-P)双参数模型构建鳙群体CO I和Cytb单倍型序列系统发生NJ树(图1),系统发育树各分支的置信度(bootstrap)均进行1000次重复检验。从CO I单倍型序列NJ树可以看出,CO I-Yong-Hap-4单倍型是Yong-KQ群体特有单倍型。4个鳙单倍型谱系分为2支,Yong-KQ群体特有CO I-Yong-Hap-4单独为一支,其余3个单倍型聚为另一支。
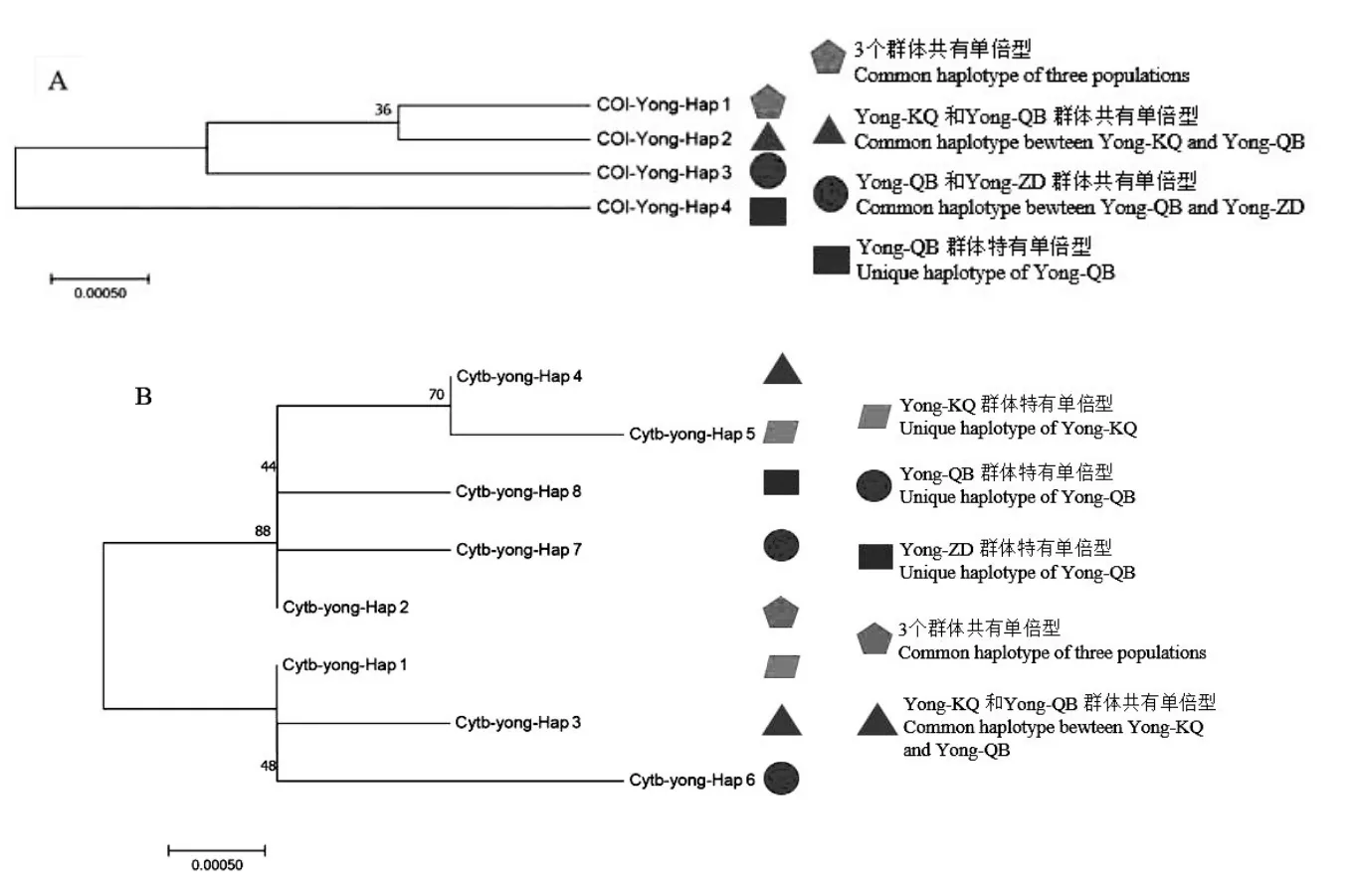
图1 基于CO I(A)和Cytb(B)序列单倍型系统发育树
Cytb单倍型序列NJ树结果显示,3个鳙群体同样存在2个明显的单倍型谱系分支,单倍型Cytb-Yong-Hap-2、Cytb-Yong-Hap-4、Cytb-Yong-Hap-5、Cytb-Yong-Hap-7和Cytb-Yong-Hap-8五个单倍型聚为一个分支,其余单倍型聚为另外一个分支,单倍型聚类关系中各群体单倍型散乱分布,3个鳙群体间并未形成明显的地理谱系。
3 讨论
3.1 群体遗传多样性
遗传多样性是生物多样性的基础和重要组成部分,遗传多样性每丢失10%,就会对生物繁育能力、存活率、生长等重要性状产生很大的负面影响。单倍型多样性(Hd)、核苷酸多样性(Pi)和平均核苷酸差异数(K)是衡量种群遗传变异的重要参数。本研究利用线粒体Cytb和CO I基因序列对丹江口增殖放流站鳙亲本群体、子代群体和丹江口库区自然群体进行了遗传多样性比较分析。基于线粒体Cytb基因序列研究结果显示3个鳙群体的平均单倍型多样性分别为0.452±0.060,平均核苷酸多样性分别为0.00102±0.00016,平均核苷酸差异分别为1.14525,低于其他流域鳙群体遗传多样性。吴伟军等利用Cytb基因序列研究红水河鳙3个野生群体发现单倍型多样性、核苷酸多样性和平均核苷酸变异差异分别介于0.667-0.8929、0.0014-0.0027和1.5556-2.9286。基于线粒体CO I基因序列研究结果显示3个鳙群体的平均单倍型多样性分别为0.385±0.059,平均核苷酸多样性分别为0.00063±0.00011,平均核苷酸差异分别为0.41304,低于其他增殖放流站鳙群体遗传多样性。刘慧芬等分析了河南省11个养殖场鳙群体的线粒体CO I基因序列,165个样品共检测出14个单倍型,平均单倍型多样性和核苷酸多样性分别为 0.86237、0.00223;Li等等通过线粒体D-Loop区和16S rRNA对比分析了长江(样品采集于汉江和长江石首段)、珠江和黑龙江鳙群体的遗传多样性,发现长江鳙群体(Hd=0.787,Pi=0.0020)低于珠江群体和黑龙江群体;不同流域和增殖放流站鳙群体显示出高单倍型多样性和低核苷酸多样性。沙航等利用微卫星标记分析了长江中游石首、监利和长沙3个鳙群体遗传多样性,结果显示3个鳙群体具有较高的遗传多样性(PIC>0.5,He>0.6);冯晓婷等基于微卫星标记分析长江下游7个原良种场鳙亲本和后备亲本遗传多样性,发现鳙亲本具有较高遗传多样性水平(PIC>0.789,He=0.889)。鱼类养殖群体的遗传多样性低于野生群体,在鲢、鲤、草鱼等养殖鱼类中均有报道,在鳙养殖群体也有遗传多样性下降的报道。丹江口水库3个鳙群体的线粒体DNA数据显示了相对偏低单倍型多样性和核苷酸多样性,可能是以下因素有关。首先,养殖场亲本在封闭的养殖条件下经过人工选择和淘汰,这可能导致亲本遗传多样性降低。其次,在采样过程中,特别是鳙子代样品,可能来自少数几个亲本的后代,表现出偏低的遗传多样性水平。再次,丹江口水库建成之后,近20多年进行了多次人工放流,放流的苗种来源多样,质量不一;如:有些繁殖场存在近亲繁殖苗种的不科学操作,或往往几条雌鱼的产量就足以满足当年鱼苗的需求量,或在进行人工繁殖时所用的亲本组合数较少,这就导致了后代群体的遗传多样性降低甚至出现生长性状退化、抗逆性降低等不利于水产养殖业发展的现象。
3.2 群体遗传结构
遗传距离、遗传分化指数(Fst)和基因流(Nm)是研究遗传结构的重要参数。本研究中根据线粒体序列数据计算出的鳙种群内和种群间遗传距离均<0.005,群体内和群体间遗传距离差异不显著,表明群体之间亲缘关系很近。AMOVA分析显示,有12.60%(CO I)、15.94%(Cytb)的变异来自于群体间,群体内的变异依然是主要来源;综合线粒体序列数据分析发现群体间差异显著(Fst=0.12604,Fst=0.15942,P<0.001)。一般认为,Fst值小于0.05时,表示群体间遗传分化极小;当Fst值在0.05-0.15之间时,说明群体间有遗传分化;当Fst值在0.15-0.25之间时,说明群体间遗传分化较大;当Fst值大于0.25时,表明群体间有极大的遗传分化。通过群体间的逐对比较进一步检测鳙群体间可能的遗传分化,Yong-ZD与Yong-KQ、Yong-QB之间Fst分别为 0.20484(P<0.001)、0.09214(P<0.05)(CO I)和 0.31930、0.22203(P<0.001)(Cytb),表明Yong-ZD与Yong-KQ群体之间有极大遗传分化,与Yong-QB群体之间有显著的遗传分化;而Yong-KQ和Yong-QB群体之间遗传分化不显著。根据对鳙群体间基因流(Nm)的检测结果,Yong-ZD与Yong-KQ群体之间有一定基因交流,而与Yong-QB群体之间有频繁的基因交流(Nm>30),这也反应出亲本与子代之间密切关系。从鳙群体间单倍型聚类关系分析可知,3个群体间均有共享单倍型,没有聚集成不同群体,另外其他2个群体相比,Yong-ZD群体只有1个特有的单倍型,这也反应了Yong-ZD群体与其他群体间的遗传分化。
3.3 鳙资源保护和利用
增殖放流是目前恢复渔业资源的主要手段之一,而放流后会不会导致鱼类遗传结构发生变化也是人们重点关注问题。本研究的结果表明,丹江口水库3个鳙群体遗传多样性处于较低水平,子代群体与丹江口水库自然群体有显著的遗传分化;亲本群体与丹江口水库自然群体之间有频繁的基因交流,遗传分化不显著。汉江是长江主要支流,在丹江口水库建成之前,分布于长江、汉江两个水系的鳙能够进行交流,丹江口水库建成之后,阻断与长江鳙交流通道。丹江口水库大坝加高蓄水后,回水区上延、水流变缓,对分布于水库上游至安康大坝江段的鳙及其他产漂流性卵的鱼类有较大影响。近年来,为库区鱼类资源的保护和可持续发展,采取了包括禁渔期、增殖放流、打击偷捕滥捕等措施。增殖放流活动在增加鳙资源量的同时,也可能改变了丹江口水库鳙群体遗传多样性水平,因此,为了改善丹江口水库天然种群的遗传结构,建议加强增殖放流遗传管理,选取来源于较高遗传多样性水平的长江鳙进行放流。
参考文献略。
猜你喜欢
世界科学技术-中医药现代化(2022年3期)2022-08-22
肝博士(2022年3期)2022-06-30
中国糖料(2021年3期)2021-07-13
中国饲料(2019年19期)2019-03-25
热带农业科技(2019年1期)2019-01-14
中国果业信息(2019年11期)2019-01-05
科学与财富(2018年8期)2018-05-09
湖北农业科学(2017年16期)2017-09-14
绿色科技(2017年10期)2017-07-05
