
天山花楸种子特征及休眠原因初探
2022-12-22 09:17:38赵善超李克一古丽米热艾合买提潘存德肖中琪陈兵权
种子 2022年11期
陈 虹,赵善超,李克一,古丽米热·艾合买提,潘存德,肖中琪,陈兵权
(1.新疆农业大学林学与风景园林学院/新疆教育厅干旱区林业生态与产业技术重点实验室,乌鲁木齐 830052;2.新疆维吾尔自治区天然林保护中心,乌鲁木齐 830000)
我国花楸资源丰富,主要分布在东北、华北地区[1],作为北方珍贵的观叶、观花、观果、观形的优良树种,其具有很高的经济、生态、药用价值[2-4]。目前有关花楸属的研究主要集中在分布特征[5]、生殖生物学[6-8]、药用价值[9-10]、扦插育苗与嫁接技术[11]等方面,且种源试验研究结果表明,花楸表型遗传性状的变异表现为种间变异大于群体间遗传变异[12-13]。新疆地区的天山花楸(SorbustianschanicaRupr)广泛分布于天山山脉海拔1 400~2 000 m的山坡、林缘、林地内[14]。在调查中发现,该树种多为单株散生,在林地里常以天山云杉(Piceaschrenkianavar.tianschanica)伴生树种零星出现[15]。因处于从属地位或相对次要的地位,通常不具备种间竞争优势,导致其再生能力下降,同时种子难以萌发,造成该树种在自然状态下天然更新极困难,给新疆平原地区的引种和繁殖、天然更新带来许多不便。因此,本研究以天山花楸种子为材料,通过分析种子结构、内源萌发抑制物类型的生物学鉴定入手,深入了解天山花楸种子休眠类型。旨在为天山花楸种子学研究提供参考,也为天山花楸开发利用提供依据。
1 材料与方法
1.1 试验材料
天山花楸种子于2020年9月采自新疆农业大学实习林场(43°16′~43°26′N,86°46′~86°57′E)内的野生母株,采回后将种子洗出自然晾干,于4 ℃冰箱保存待用。
1.2 试验方法
1.2.1种子基本特性及结构观察
按照林木种子检验规程(GB/T 2772—1999)相关标准进行花楸种子基本特性检测[16],用游标卡尺测定种子大小、万分之一电子天平测定种子重量、红四唑(TTC)测定种子生活力;并用倒置解剖镜进行花楸种子基本解剖特性观察;每个处理重复3次。
1.2.2种皮结构的观察
以未处理种子种皮为对照,与45 d萌发种子的种皮置于75%无水乙醇中,经逐级脱水后,用HCP-2临界点干燥仪粘合—IB-v粒子溅射仪喷金—PHILIPS SEM-505扫描电子显微镜观察,并对种皮横断面结构进行记录和拍照,分析变化情况。
1.2.3种子透水性研究
取完整的种子和破皮种子各100粒,称其质量,浸入3倍种子体积的蒸馏水中,25 ℃恒温条件下进行种子透水性吸胀实验,前12 h每2 h取出种子后,用滤纸快速吸干表面水分并称重,随后每12 h称重1次,直至质量恒定。3次重复。
吸水率(%)=[(湿质量-干质量)/干质量]×100%。
1.2.4种子中抑制物的测试
参照尹德洁等[17]的方法进行种子抑制物测定,具体方法如下:取种子1 g,经充分研磨后,分别用甲醇、去离子水定容至10 mL,于4 ℃振荡培养箱中浸提36 h后4 ℃、4 000 r/min离心15 min取上清液。并用旋转蒸发仪(56 ℃)对甲醇上清液进行处理,待甲醇挥发后,用蒸馏水溶解剩余物质,定容至10 mL;去离子水上清液直接定容至10 mL。得到浓度为0.1 g/mL的甲醇和去离子水浸提液。以清水为对照,设计0.01、0.05、0.1 g/mL 共3个实验浓度,测定白菜种子发芽率、根长、苗高,并计算内源萌发抑制物活性。每个试验重复3次。
1.3 数据处理与分析
采用SPSS 18.0软件进行单因素方差分析和多重比较、使用Excel软件进行图表处理。
2 结果与分析
2.1 天山花楸种子基本特性
种子表观和解剖结构观察表明(图1),成熟的天山花楸种子为红褐色,性状呈扁卵形,先端尖,侧向压扁(腹面),微弯曲,背面隆起,有光泽,种皮较光滑。纵径在4.01~4.97 mm之间,横径在1.89~2.67 mm之间;种子平均千粒重为5.556 g。种子含水量约为6.47%,种子的生活力为33.3%。表明天山花楸种子属于小粒种子,且生活力不高。种子由种皮和种胚两部分构成。经测量其种皮厚度约为164 nm,由外种皮和内种皮组成,外种皮坚硬,两端较厚,木质化程度高,外种皮表面覆有角质层;内种皮膜质半透明,内侧呈白色,光滑有光泽(图1 b)。种胚包括胚芽、胚轴、胚根、子叶四部分,胚发育完全,为典型的无胚乳种子。胚芽、胚轴、胚根长度约为1.02 mm,子叶为两片,长度约为3.75 mm,均为乳白色。说明天山花楸具有完整的胚。
2.2 天山花楸种子透水性
通过测定完整种子及破皮种子的天山花楸种子的吸水率可知(图2),天山花楸种子的吸水过程分为快速吸水(0~2 h)、缓慢吸水(2~24 h)、近饱和吸水(24~36 h)3个阶段。且完整种子与破皮种子的吸水速率曲线变化过程的趋势基本一致,即快速吸水阶段,种子吸水速率急剧上升,随后种子的吸水速率下降,待近饱和阶段,两者吸水速率基本一致。经T检验结果表明,破皮种子与完整种子的吸水速率在快速吸水阶段存在极显著差异(p<0.01),破皮种子和完整种子的吸水率分别为(61.1±0.45)%和(35.8±0.21)%;而近饱和阶段,破皮种子与完整种子的吸水率之间差异不显著(p<0.05),破皮种子和完整种子的吸水率为(76.3±1.27)%和(83.5±2.47)%。与破皮种子相比,完整种子的吸水速率缓慢,说明种皮存在透性障碍,种子存在吸水障碍。

注:a为种子外观及大小;b为内、外种皮结构;c为角质层;d为胚结构。图1 天山花楸种子形态和解剖结构Fig.1 Seed morphology and anatomical structure of Sorbus tianschanica
2.3 天山花楸种皮结构的观察
运用扫描电镜技术,对比未经处理的干种子和萌发后种子的种皮结构(图3)可知,干种子种皮厚度约为164 nm,且表面覆有一层角质层,木栓质化严重,细胞呈不规则排列且排列紧密,部分地方有褶皱。内种皮紧贴外种皮,表面较光滑,其细胞呈纵向排列,排列致密。而萌发后的种皮厚度约为27.3 nm,约为干种子种皮厚度的1/6。种皮由光滑变得粗糙不平,内种皮与外种皮之间出现分离,且部分内种皮内卷。萌发后的种皮角质层基本破裂,部分已完全脱落,外种皮暴露在外,细胞间空隙增大,透性增大,机械阻力减小。说明种皮对天山花楸种子透水透气性影响很大,外种皮上的角质层和坚硬较厚的种皮会影响种子的透性,不利于种子进行水分交换,影响种子萌发。
2.4 萌发抑制物对白菜种子萌发特性的影响
甲醇和水分别浸提花楸种子后进行白菜种子萌发抑制物测定(表1)可知,花楸种子甲醇和水浸提液下的白菜种子发芽率、主根长、主苗高组间存在显著差异(p<0.05)。甲醇和水浸提液浓度为0.01、0.05、0.1 g/mL时,发芽率比对照分别降低10.4%、13.7%、15.0%和13.6%、15.9%、19.6%。对照白菜主根长和苗高分别为2.12 cm和2.93 cm,而不同浓度甲醇浸提液、水浸提液处理后的主根长为1.43~1.98 cm和1.35~1.84 cm,与对照相比,主根长分别减少了6.6%、11.8%、32.5%和13.2%、25.9%、36.3%;苗高分别为1.76~2.43 cm和1.64~1.88 cm,苗高与对照相比分别减少了17.0%、34.1%、39.9%和35.8%、40.9%、44.0%。综上表明,花楸种子中含有影响种子萌发的水溶性内源抑制物和醇溶性内源抑制物,且随着浸提液浓度的增加,内源抑制物的活性和抑制种子发芽及幼苗生长的作用增强,同时种子水浸提液下白菜种子的萌发率、主根长、苗高低于甲醇浸提液处理,这可能是由于花楸种子萌发抑制物的水中溶解性大于甲醇中的溶解性。
3 讨 论
种子的形态学特征是种子长期进化过程中对环境条件的一种适应机制,其大小、形状等对种子的传播及其幼苗生长策略具有重要影响[18]。本实验结果表明,天山花楸种子平均纵径约4.47 mm,平均横径约2.16 mm,平均千粒重为5.556 g,而水榆花楸种子的大小(平均纵径为6.12 mm,平均横径为3.48 mm),尤其是重量,为天山花楸的4倍以上[19],说明不同种的花楸种子在形态特征上有较大差异,且多种植物的研究中发现,小种子的发芽率低[20]。此外种子的生活力对于种子能否成活也起到至关重要的作用,一般情况下,高活力的种子在适宜的条件下发芽快,出苗整齐。天山花楸种子生活力为33.3%,低于水榆花楸[19](Sorbusalnifolia(Sieb. & Zucc.) K. Koch)和黄山花楸[21](Sorbusamabilis)种子生活力,表明天山花楸种子的活力不高,这是否与种源地的母树树势有关有待进一步研究。
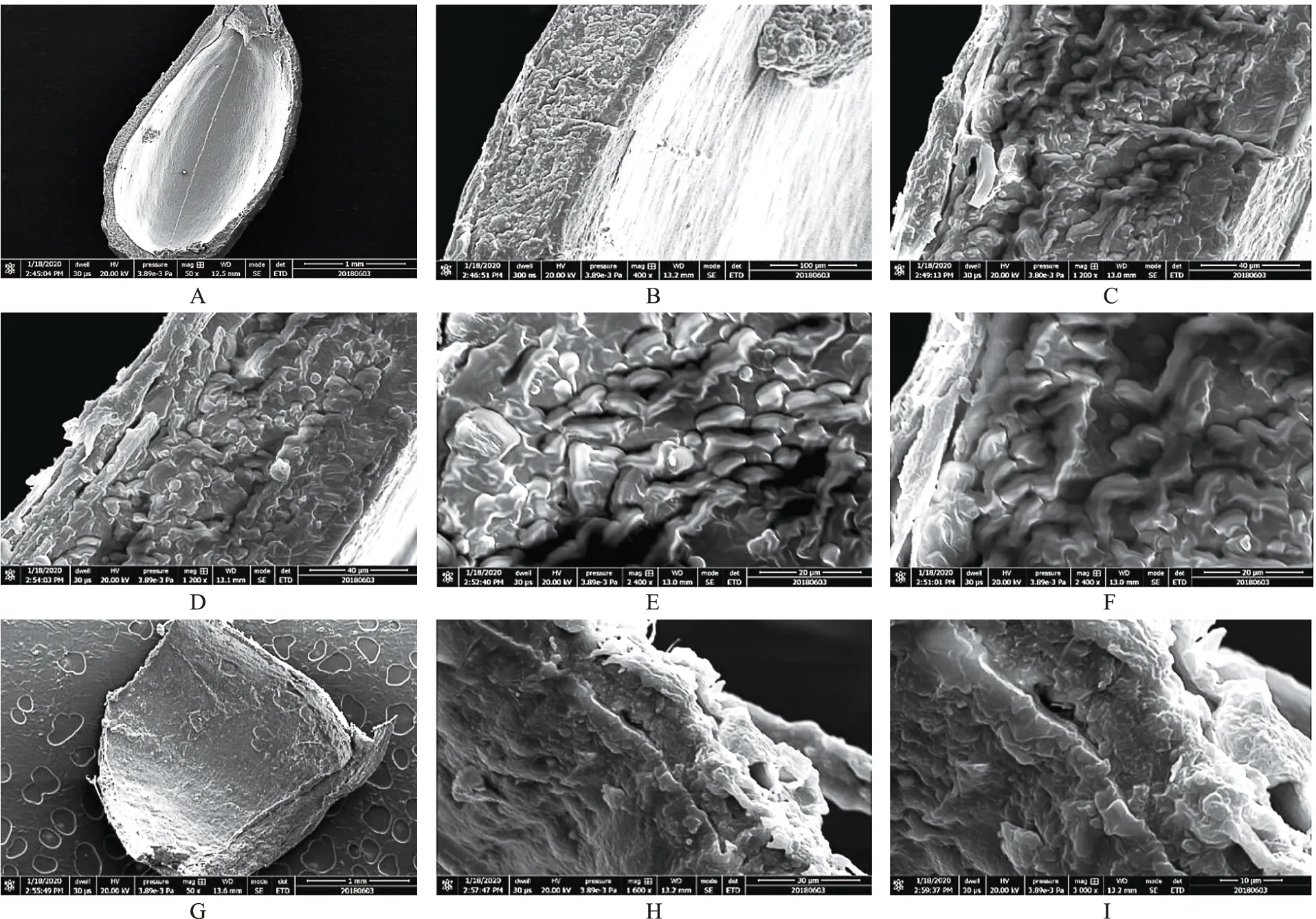
注:A为干种子种皮整体结构(×50);B为干种子种皮断面结构(×400);C为干种子种皮断面结构(×1 200);D为干种子种皮断面结构(×1 200);E为干种子种皮断面结构(×2 400);F为干种子种皮断面结构(×2 400);G为发芽后种子部分种皮结构(×50);H为发芽后种子种皮断面结构(×1 600);I为发芽后种子种皮断面结构(×3 000)。图3 扫描电镜下天山花楸种子种皮结构Fig.3 The seed coat structure of Sorbus tianschanica under SEM
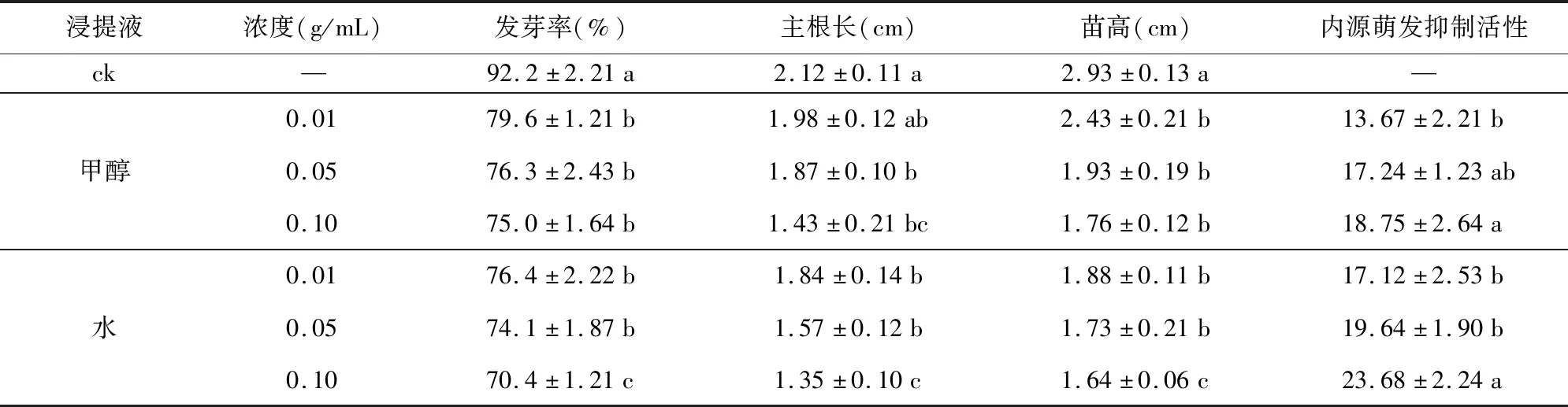
表1 种子浸提液对白菜种子萌发及幼苗生长影响Table 1 Effects of different concentrations of extracts from the seeds of Sorbus tianschanica on the germination rate and seedling growth of Chinese cabbage seeds
植物为适应环境保持自身繁殖发展而形成的种子休眠特性对于植物本身具有重要的生态学意义[22-23]。很多研究指出,引起植物种子休眠的原因主要与种皮结构[24]、种子抑制物的存在[25]、胚发育不完全[26]、种子代谢缓慢[27]有关。通过解剖观察发现,天山花楸种子的种胚包括胚芽、胚轴、胚根、子叶四部分,胚发育完全,表明天山花楸具有完整的胚,不存在生理后熟,这与水榆花楸、单叶蔓荆(VitextrifoliaLinn.var.simplicifoliaCham.)的研究结果一致[17]。种皮在具有机械束缚作用的同时还具有透性障碍,本研究中,通过扫描电镜观察对比无处理干种子和萌发后种子的种皮结构发现,干种子的种皮厚度约为164 nm,且种子外种皮表面覆有一层角质层,内部木栓质化程度严重,内种皮种子的结构致密,这些限制性条件阻碍了天山花楸种子种胚与外界的水分交换,影响种子的吸胀和萌发,进而影响天山花楸种子的萌发。在东北红豆杉(TaxuscuspidataSieb. et Zucc.)种子[28]、青钱柳(Cyclocaryapaliurus(Batal.) Iljinsk.)种子[29]的研究中,均指出外种皮角质化、中种皮有木栓质,内种皮木非常坚硬等种皮结构是阻碍胚与外界的水分和气体的交换的影响因素。虽然天山花楸与黄山花楸的种皮结构具有一致性,但天山花楸的种皮厚度(约164 nm)远大于黄山花楸(约33.75 nm)种皮厚度[21],进一步证实了不同种的花楸种皮特征具有差异性,可能这也是天山花楸较花楸属其他种更难萌发的原因。种子吸水性结果进一步指出,因致密的种皮结构造成的透性障碍是造成天山花楸种子难以萌发的原因。这在紫楠(Phoebesheareri(Hemsl.) Gamble)种子[30]、水榆花楸[19]、黄山花楸[21]等相关研究中也得到相似结论。同时天山花楸种子甲醇和水浸提液能明显抑制白菜种子萌发和生长,说明花楸种子中含有影响种子萌发的水溶性内源抑制物和醇溶性内源抑制物,且种子萌发抑制物的水中溶解性大于甲醇中的溶解性。因此,种子外种皮角质化和中种皮有木栓质和种子中的内源抑制物是造成天山花楸休眠的重要因素,需进一步明确种子中的休眠物质成分,为人工繁育提供技术解决方案。
4 结 论
天山花楸种子属于小粒种子,且生活力较低,种皮机械障碍造成的物理休眠和种胚内源抑制物是造成天山花楸种子休眠的主要因素。
猜你喜欢
粮油与饲料科技(2022年2期)2022-11-24 13:12:07
中国野生植物资源(2022年3期)2022-04-19 09:13:00
中国民间疗法(2021年7期)2021-07-22 06:43:50
杂文月刊(2020年3期)2020-05-21 10:44:47
贵州农业科学(2020年1期)2020-03-25 08:14:48
植物资源与环境学报(2019年3期)2019-09-12 06:17:32
安徽农业科学(2018年1期)2018-05-14 08:59:41
福建中医药(2017年6期)2018-01-09 06:49:54
西南农业学报(2016年5期)2016-05-17 05:42:27
西南农业学报(2016年4期)2016-05-17 05:42:14
