
马尾松非结构性碳库大小及分配的纬向变化*
2022-12-21 11:04倪妍妍简尊吉曾立雄阮宏华肖文发李迈和
林业科学 2022年8期
倪妍妍 简尊吉 徐 瑾 曾立雄, 阮宏华 雷 蕾 肖文发, 李迈和
(1.中国林业科学研究院森林生态环境与自然保护研究所 国家林业和草原局森林生态环境重点实验室 北京 100091; 2.南京林业大学南方现代林业协同创新中心 南京 210037; 3.瑞士联邦森林、雪与景观研究所 苏黎世 8903; 4.东北师范大学地理科学学院 长白山地理过程与生态安全教育部重点实验室 长春 130024)
非结构性碳水化合物(non-structural carbohydrates, NSC)是植物生存和生长的物质基础(Chapinetal., 1990; Martínez-Vilaltaetal., 2016)。NSC库由可溶性糖(蔗糖为主)和淀粉组成。树木光合同化形成的碳优先供给呼吸和生长后,将剩余的碳贮存形成稳定的NSC库(Barbarouxetal., 2002; Goughetal., 2009; Rochaetal., 2009)。NSC的动态变化可作为评估碳“源-汇”关系的指标(Körner, 2003),NSC库大小与物候节律、树木个体大小及所处环境等密切相关(倪妍妍等, 2017; Schoonmakeretal., 2021)。此外,Richardson等(2013)通过美国通量网Howland定位站长期观测发现,NSC的长期逐渐增加量与木质生物量的增加比例基本一致,表明NSC库不仅影响树木个体生长,还在生态系统碳循环中发挥重要作用。然而,迄今关于树木体内NSC的储存与分配特征的研究相对较少。
树木器官NSC库的大小可表征林分地上和地下生产力的分配情况(Lietal., 2018)。树木不同器官的NSC含量差异显著(Boldingh, 2000; Lietal., 2001; Smithetal., 2017),因此,利用单个器官NSC含量无法准确估计整个植株的NSC存储量。此外,树木不同器官对植株生长和代谢过程的作用与贡献不同(Lietal., 2001; 2002; Hochetal., 2003; Palacioetal., 2014)。例如,常绿树种叶片既是植株碳水化合物生产的源(Kozlowski, 1992; Hartmannetal., 2016),也是储存碳水化合物的库(Lietal., 2001; 2002); 树干除了作为碳水化合物的高消耗器官外,同时承担了输导功能和存储功能(Barbarouxetal., 2003; Hochetal., 2003); 根系含有较大比例的射线和轴向薄壁组织细胞,具备相对较高的存储能力,且这些储存主要被用于次年新枝生长(Lensetal., 2000; Zhuetal., 2012)。但Furze等(2019)研究发现,温带5个树种的枝条全年储存了足够多的淀粉,导致根系并不是主要储存器官,而且根系的储存并没有用于支持春季生长。因此,探究不同器官NSC库的动态变化,有利于深入了解树木的生长动态及其对环境变化的响应。
NSC的储存是对不同时空尺度下碳水化合物供应与需求非同步性的一种缓冲,具有明显的时空效应(Hochetal., 2012)。树木NSC含量高低和储存大小除与物种生物学特性和自身生长机制密切相关外(Congetal., 2018),也与生长环境密切相关(Wangetal., 2018)。树木NSC含量随纬度、海拔的变化特征有诸多研究(Martínez-Vilaltaetal., 2016; Liuetal., 2018; Liuetal., 2019; 杜建会等, 2020)。如Martínez-Vilalta等(2016)发现从寒温带针叶林到热带雨林,NSC及其组分在树干和根系中的含量随纬度无显著变化,而在叶片中的含量随纬度增加而增加。目前,在大空间尺度上探究某单一树种不同器官NSC储存量变化的研究较少(Liuetal., 2019)。
马尾松(Pinusmassoniana)是我国重要的乡土用材树种,在亚热带湿润区域广泛栽植,是探究跨地域尺度分布物种生存策略的理想材料。因此,本研究以马尾松成熟林为研究对象,在其分布区内沿纬向梯度在不同纬向梯度选择9个典型人工林,研究马尾松NSC存储及分配的纬向变化规律,并探讨生长环境对其影响作用,为准确理解和模拟气候变化背景下树木生长和碳存储提供依据。
1 材料与方法
1.1 研究区概况
根据马尾松在我国的分布和生长情况,在不同纬度(23.0°—33.5°N)选择9个马尾松人工林纯林,其中马尾松蓄积量不低于林分总蓄积量的65%。采样点从南到北依次为广东肇庆、广西贺州、广西桂林、湖南会同、湖南永顺、湖北宣恩、重庆忠县、四川万源和陕西汉中。样地年均温度和年均降水量从南到北递减,地带性土壤类型主要为红壤、黄壤和黄棕壤。依据国家标准《森林资源连续清查技术规程GB/T38590-2020》(国家林业和草原局, 2020)中马尾松人工林的龄组划分,优先选择林龄30~35年段的成熟林,其次选择25~30年段的近熟林。各地气候特征和林分结构存在差异,具体信息详见表1。
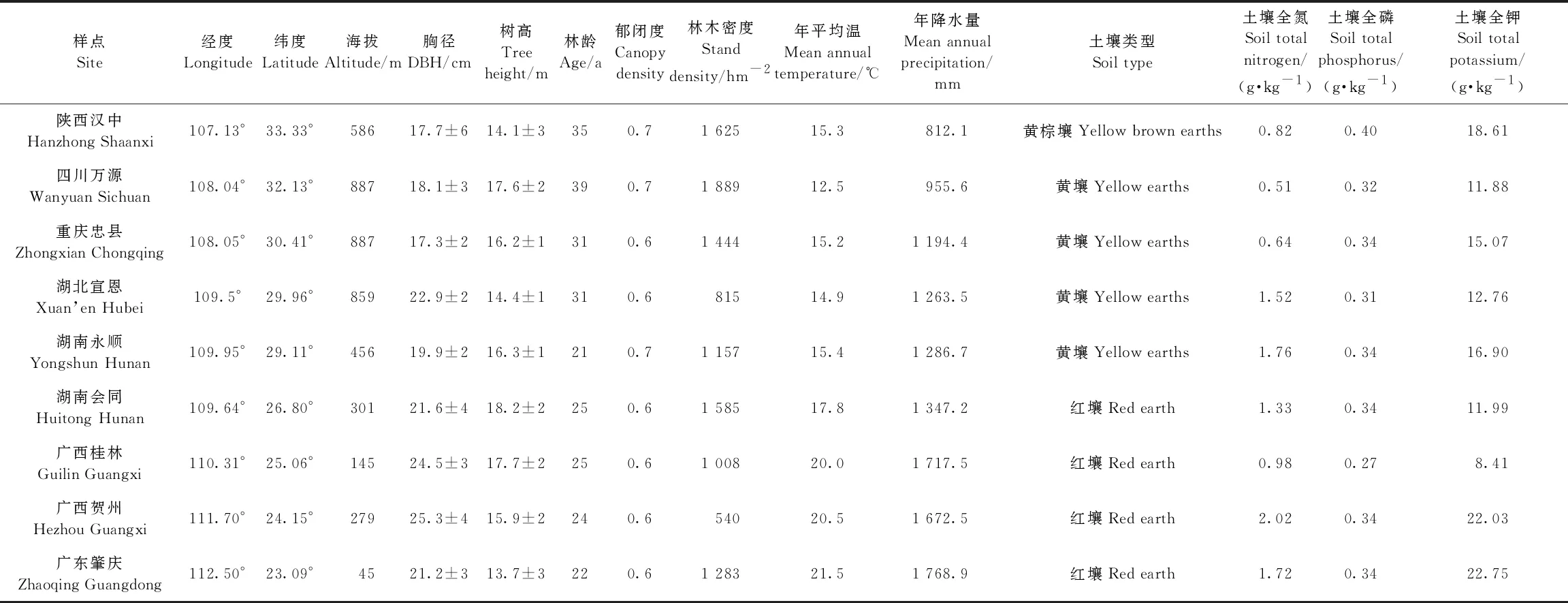
表1 马尾松人工林样地的地理位置、气候和林分特征
1.2 野外调查及样品处理
2018年10—12月,从北到南依次采集样品。每个样点选择3~4块样地(20 m × 30 m), 同一样点每2块样地间的间距不小于1 km。在每块样地内选择10~15个点,用土钻收集表层土(0~20cm),混装成1个样品,用于测定土壤养分。每木检尺记录样地内所有胸径大于4cm的马尾松的胸径和树高,确定5~7株标准木用于收集根、茎、枝、叶等器官样品。对每一株标准木,在树高2/3~3/4处选择2个阳生枝收集针叶和枝条; 利用生长锥在树干1.3 m胸径处钻取树干样品用来测定树木年龄并测定NSC含量; 利用军工铲收集细根。植物器官样品按照样地进行混装。在进入实验室预处理前,样品放置在保鲜箱。
在实验室内,将枝、茎、根去皮。所有植株样品用水清洗干净后用蒸馏水冲洗2次,于105℃下杀青15min,在65℃下烘干至恒质量,粉碎、过筛(0.15mm孔径)后储存在磨口瓶中待用。土壤样品置于阴凉处风干,过筛(0.07mm孔径)后储存待用。
1.3 NSC及土壤养分含量测定
NSC含量为同一样品中可溶性糖和淀粉之和。采用蒽酮比色法测定样品可溶性糖和淀粉含量(Lietal., 2018)。
土壤样品(1.2~1.5 mg)用专用锡箔纸包裹后置于元素分析仪(Euro EA, Hekatech Gabh, 德国)上测定全碳、全氮元素含量。样品先用酸消解(HNO3和HClO4)后用离子发射光谱仪(IRIS Intrepid II XSP, Thermo, 美国)测定全磷和全钾含量。
1.4 植物器官生物量估算和NSC及其组分储存量计算
用异速生长方程模型估算样地马尾松各器官的生物量:
M=aDbHc。
式中:M为各器官生物量;D为胸径;H为树高;a,b,c为参数,具体数值参考林业行业标准《立木生物量模型及碳计量参数——马尾松LY-T 2263-2014》(国家林业和草原局, 2014)。根据不同区域的异速生长方程模型求算样地马尾松的各器官生物量。
与分布区北界相比,马尾松在南界生长快、生长季长而轮伐周期短(表1)。为了标准化地表达NSC储存,采用树木各器官生物量及整株总生物量的年平均值。马尾松各器官的NSC库是利用器官NSC含量乘以器官年均生物量计算获得。树木NSC库为各器官库总和。
1.5 气象因子
从中国气象数据网(http:∥www.data.cma.an/)下载马尾松分布区内气象站点逐年数据。本研究选择1979—2018年间气候平均值数据。用Arcgis v10.0 进行克里金空间插值, 生成分辨率1 km的气象栅格图层,最后根据地理坐标提取各样地的气象数据。气象变量包括年平均温度、年均降雨总量、年均最冷月最低温、年均最热月最高温和帕默尔干旱强度指数。
1.6 数据分析
数据分析和作图均由R4.0.5软件(http:∥www.r-project.org/)完成。所有数据通过正态分布(Shapiro-Wilk检验)和方差齐性检验(Bartlett检验),满足数据进一步分析条件。利用线性混合效应模型,将样地和树龄作为随机因子,分析器官、纬度及其交互作用对马尾松生物量和NSC及其组分的含量、库大小以及库分配比例的影响。利用简单线性回归模型分析植株整体和各器官的NSC及其组分的浓度含量、库大小以及库分配比例随纬度变化的线性趋势。通过皮尔逊相关分析,分析马尾松整株NSC及其组分库大小与环境因子的关系,再按气象因子和土壤因子将显著的因子分类进行冗余分析,获得各因子的解释率。
2 结果与分析
2.1 器官生物量与非结构性碳水化合物含量的纬度变化
如表2所示,器官、纬度以及两者间的交互作用对马尾松年均生物量、可溶性糖和NSC含量影响显著(P<0.01),纬度对淀粉含量作用显著(P<0.001)。马尾松各器官(根、干、枝和叶)的年均生物量均随纬度升高而降低(图1),干中NSC、可溶性糖和淀粉含量也随纬度增加而降低,但在其他器官中无显著趋势(图1)。
2.2 非结构性碳水化合物存储的纬度变化
马尾松整株NSC库、可溶性糖库和淀粉库的大小均受纬度影响显著(P<0.001, 表3),且均随纬度增加而降低(图2)。器官、纬度及其两者间的交互作用对NSC库、可溶性糖库和淀粉库的大小影响显著(P<0.001, 表3)。各纬度的枝、干和根的淀粉库显著高于可溶性糖库(表4),且枝和根中的可溶性糖库、所有器官中的淀粉库和枝、干、根中的NSC库均随纬度增加而降低(图3)。

表2 器官、纬度及其交互作用对马尾松的生物量和NSC含量的影响(F值)①
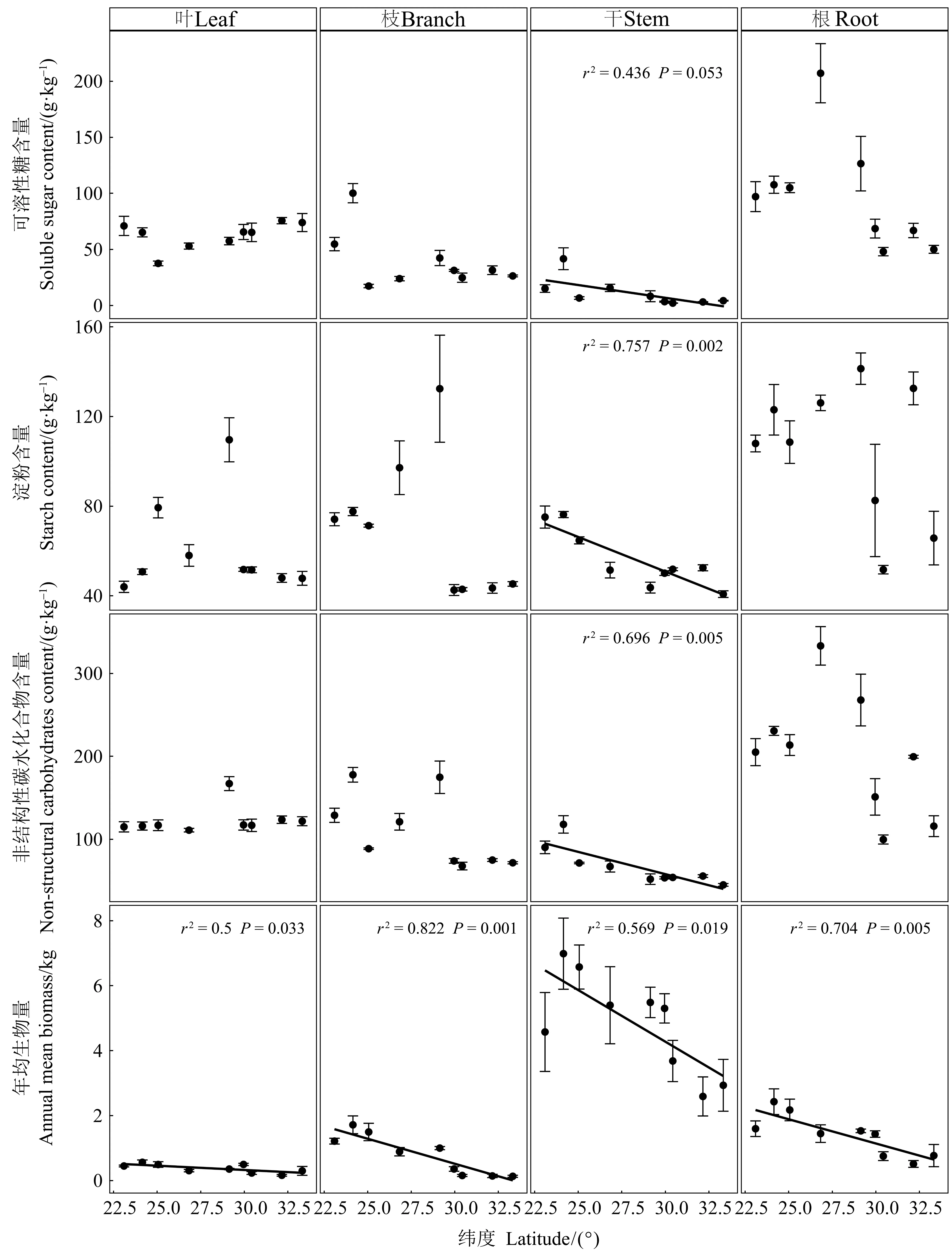
图1 马尾松不同器官年均生物量和非结构性碳水化合物含量的纬向变化
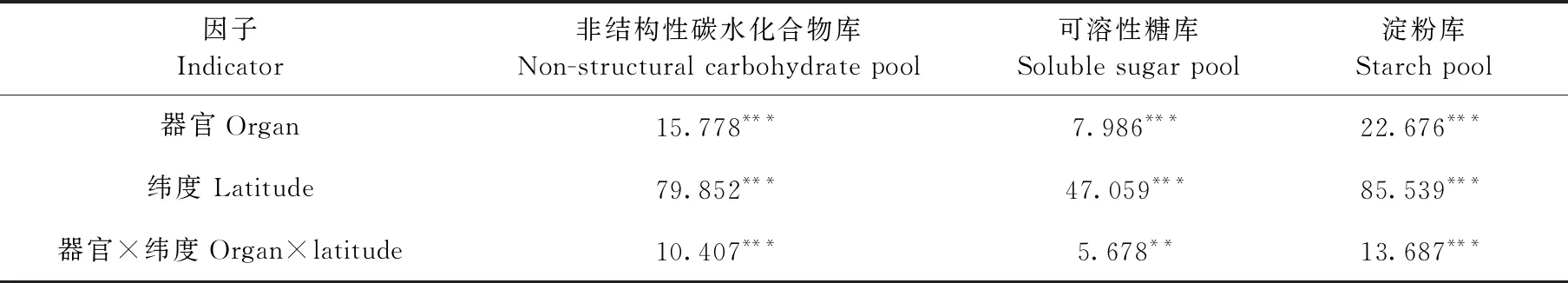
表3 器官、纬度及其交互作用对马尾松的NSC库大小的影响(F值)
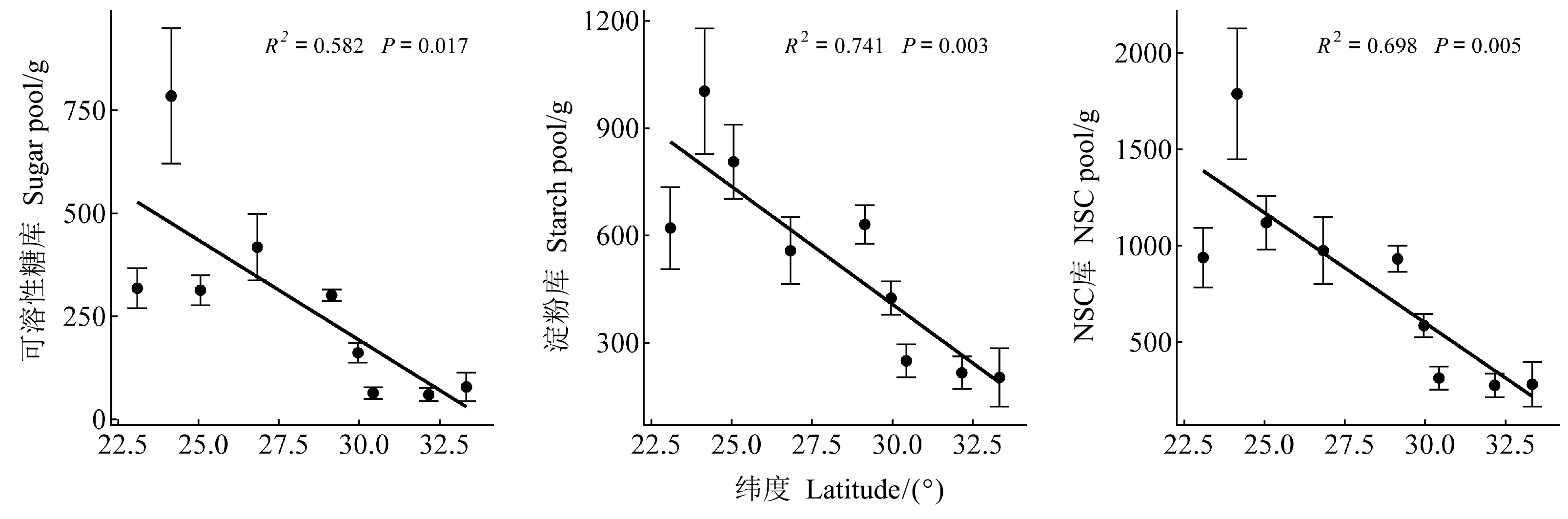
图2 马尾松整株NSC及组分库的纬向变化

表4 马尾松不同器官NSC及组分库的大小(g)和分配比例(%)
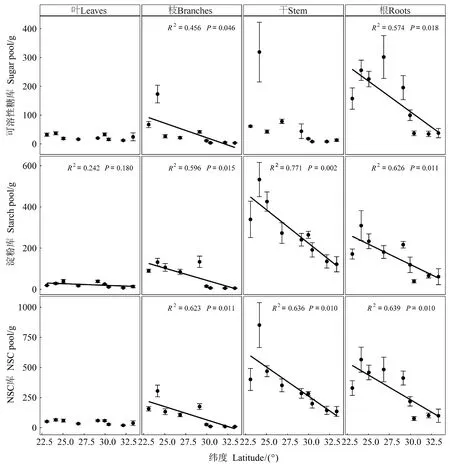
图3 马尾松不同器官NSC及其组分库大小的纬向变化
2.3 非结构性碳水化合物库在各器官中的分配比例
可溶性糖库和淀粉库平均大小分别占全树NSC库的31.1%和68.9%,表明马尾松NSC库组成整体以淀粉为主。马尾松整株的可溶性糖库占NSC库的比例随纬度增加而降低,淀粉库所占NSC的比例随纬度增加而增加(图4)。由表4和图5可知,NSC库、可溶性糖库和淀粉库在根、干、枝和叶中的分配比例差异均显著,但均主要储存在干和根中。随纬度增加,马尾松整株的NSC库、可溶性糖库和淀粉库在枝中的分配比例降低,可溶性糖库和NSC库在叶中的分配比例增加(图5)。
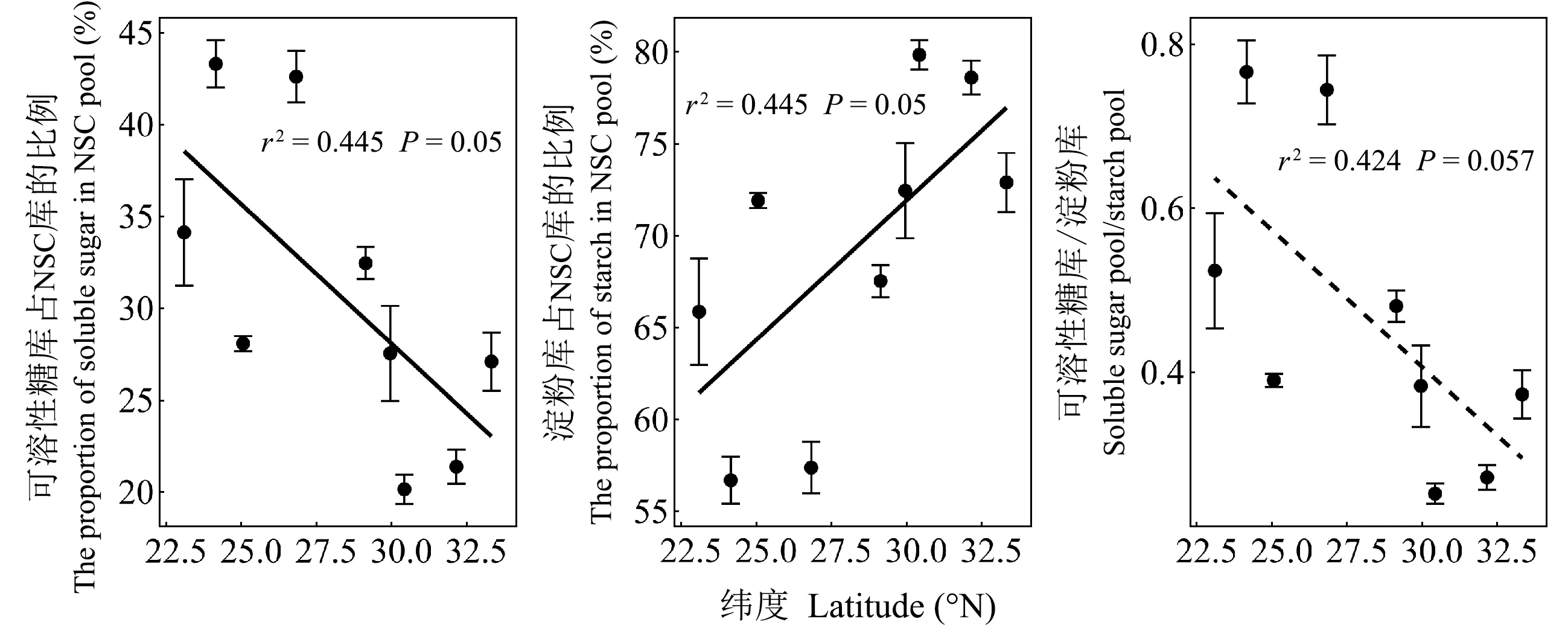
图4 马尾松整株可溶性糖库和淀粉库占NSC库的比例以及可溶性糖库和淀粉库比值的纬向趋势
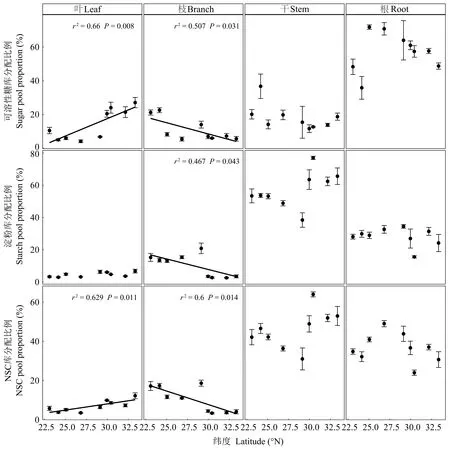
图5 马尾松整株的NSC及其组分库在根、干、枝和叶中分配比例的纬向趋势
2.4 非结构性碳水化合物库与环境因子的关系
由表5可知,马尾松整株的NSC库、可溶性糖库和淀粉库的大小与气象因子年平均温度、年均降雨量、年均最冷月最低温、年均最热月最高温、土壤全氮含量和土壤氮磷比显著正相关,但与帕默尔干旱强度指数、土壤全磷含量、土壤氮钾比和土壤磷钾比不相关,可溶性糖库的大小与土壤全钾含量正相关(表5)。将显著的影响因子按气候(年平均温度、年均降雨量、年均最冷月最低温和年均最热月最高温)和土壤(全氮、全钾和氮磷比)分类进行冗余分析方差分解,发现所有因子对NSC库、可溶性糖库和淀粉库的大小变化解释率高达70.5%,其中气象因子和土壤因子解释率分别为26.2%和7.6%,共同作用解释率为36.7%(图6)。
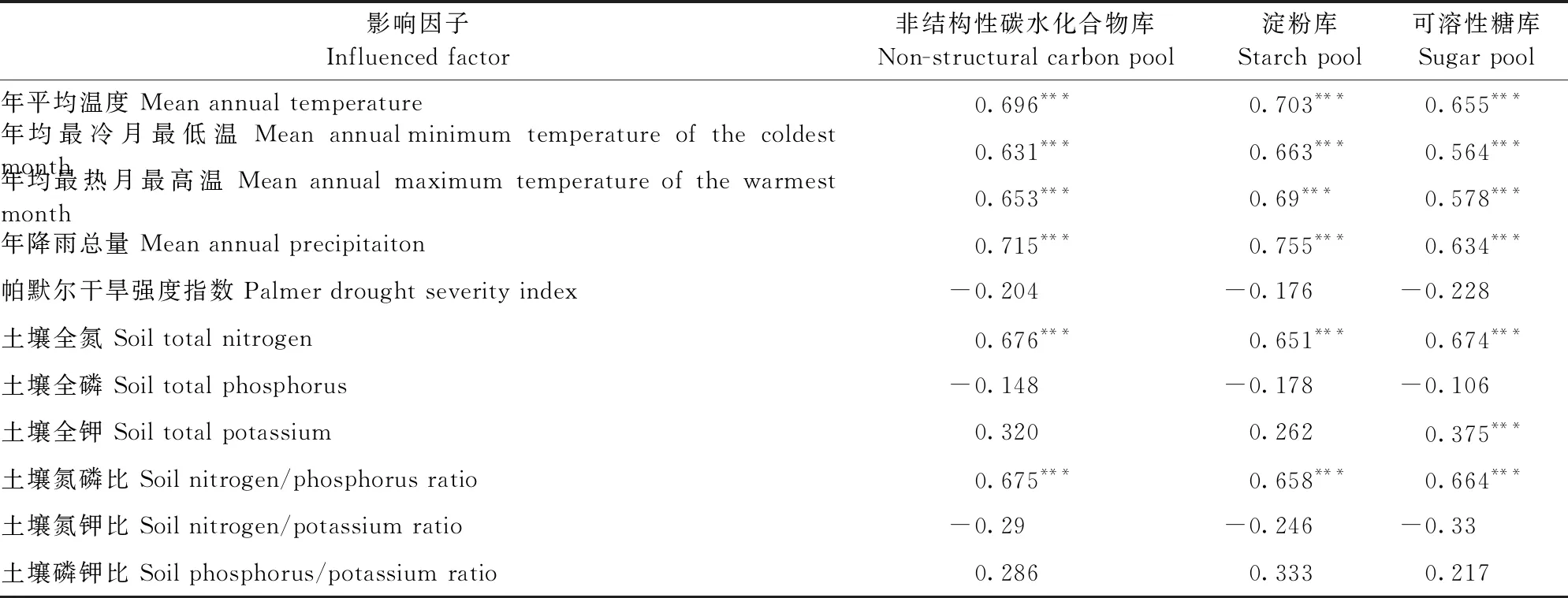
表5 马尾松NSC及其组分库与环境因子间的相关性
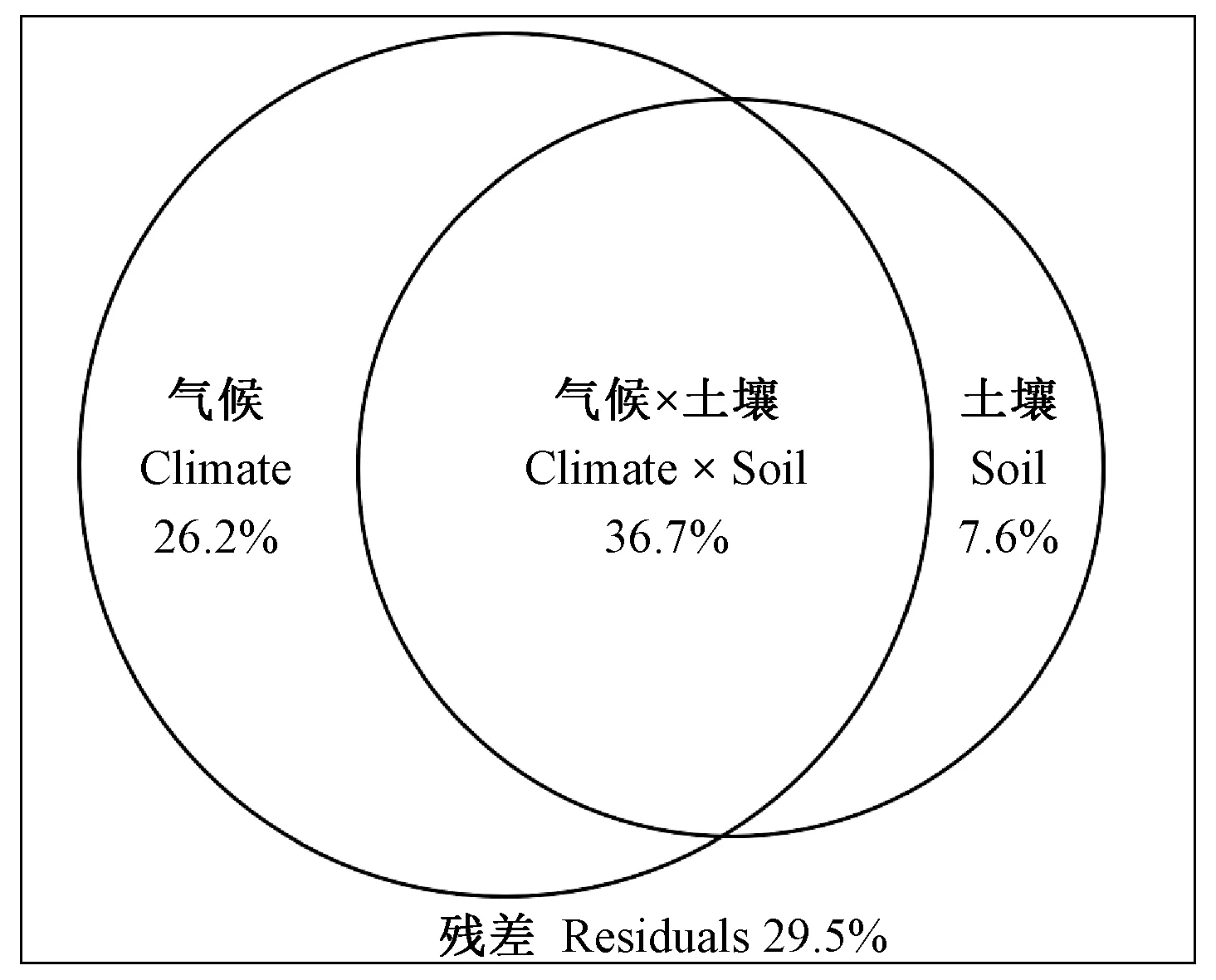
图6 气候与土壤对NSC及其组分库影响的方差分解结果
3 讨论
3.1 马尾松非结构性碳水化合物库的大小及分配差异
马尾松淀粉库的大小平均约占NSC库的68.9%,显著高于可溶性糖库(图2),表明马尾松植株整体NSC库的组成以淀粉库为主。本研究探讨的是生长季节末的NSC储存情况。在休眠季,NSC以淀粉储存为主,这与Richardson等(2015)研究中红栎(Quercusrubra)整株储存结果一致。目前,大多数研究侧重于器官含量而不是树木个体NSC库,发现尤其在枝边材(Hochetal., 2003)和干边材(Hochetal., 2003; Richardsonetal., 2013)淀粉含量高于可溶性糖含量。
植物体内NSC的储存大小和地上地下生物量的积累紧密相关(Schönbecketal., 2018)。本研究发现马尾松整株的NSC、可溶性糖和淀粉都主要储存在干和根中(图3, 5),表明树干和根是马尾松最重要的NSC储存器官,也反映出在非生长季植株多余的碳通过枝往树干和根中进行存储。树木器官根据自身各功能需求,体内NSC可以短期或者长期被分配存储在不同器官中(Richardsonetal., 2013),从而支持自身在呼吸大于光合的情况下(如春季发芽)或在不可预测的压力(干旱、极端气候等)下能够生存(Hochetal., 2003; Kleinetal., 2014)。另外,处于主林层的树木受光胁迫的概率较小,树体较大,多年积累致使NSC总量较多(Kobe, 1997; Myersetal., 2007),因此树干具有最高水平的NSC储存。
此外,马尾松NSC库在地上和地下器官的存储比例存在差异(表4, 图5),树干主要储存淀粉,而树根主要储存可溶性糖。这一差别可能因为生长季末地上部分已停止生长,地上部分的大多数可溶性糖转化为淀粉,致使树干淀粉储存最多。另外,因土壤温度高于大气温度,地下根系生长仍未停止,根系仍需大量可溶性糖用于呼吸消耗。与树干和根相比,马尾松叶和枝的储存比例相对较低(表4, 图5),这主要和它们的生物量相对偏低有关。此外,枝存储的NSC主要用来抵御冬季严寒和供给来年新叶生长(Kleinetal., 2014)。而常绿树种的叶则具有碳水化合物生产的“源”和“库”的双重功能,即在生长季节因其光合作用是源,在非生长季节则因其储存大量资源成为库(Lietal., 2002)。
3.2 马尾松非结构性碳水化合物库的纬向特征
马尾松整株NSC库、可溶性糖库和淀粉库的大小随纬度增加而降低(图2),在器官(枝、干、根)的大小也呈现相同趋势(图3),与我国不同区域森林碳储备空间格局的研究结果一致(周玉荣等, 2000; Lietal., 2004; Wenetal., 2016)。本研究表明随着纬度增加,马尾松碳供应不足,限制树木生长。叶片作为植物碳水化合物的合成器官,高纬度的植株通过增加叶片光合物质积累能力来应对相对较短的生长季,进而减弱其NSC存储的纬向趋势。然而,可利用性碳库的大小与树木生长之间的消长关系一直存在争议。如倪妍妍等(2017)发现,栓皮栎幼苗根NSC库的大小随纬度增加而增加,并认为树木通过主动调节体内NSC含量并维持在一定含量范围,而不影响植株生长(Lietal, 2018)。笔者认为这种差异主要由物种间碳的积累与分配方式(Liuetal., 2018)、物种年龄阶段(Guoetal., 2014)以及统计调查空间尺度的大小不同(Wenetal., 2016)造成。此外,本研究还发现,马尾松整株可溶性糖库占整个NSC库的百分比随纬度增加而降低,导致可溶性糖库与淀粉库的比值随纬度增加而降低(图5)。这可能是因为相对低温的环境致使植物活性降低,较高的淀粉储存比例利于生长在分布北界的树木将自身的淀粉水解成可溶性糖来抵御冬季严寒(Dangetal., 2015)。
3.3 环境对非结构性碳水化合物存储的影响
温度和降水对NSC的含量和储存具有显著影响(Körner, 2003; Hochetal., 2012)。在本研究中,马尾松整株NSC库、可溶性糖库和淀粉库的大小与年平均温度、年均最冷月最低温和年均最热月最高温正相关(表5)。NSC的储存大小随温度增加而增加,表明低温直接限制碳活性并影响结构组织的生长,进而限制碳水化合物的积累。树木储存的可溶性碳水化合物会用来弥补和缓冲由低温导致较低的光合产物无法维持生长的不足(Lietal., 2002)。NSC存储与年均降雨量正相关,但与帕默尔干旱强度指数无显著相关性(表5)。这表明,干旱胁迫降低森林碳储备,但植物通过主动调节体内NSC含量并维持在一定的含量范围来保证植物存活与生长(Lietal., 2016)。如Würth等(2005)发现,热带森林维持较高水平的碳供应确保树木在干旱季节生长不受碳限制。在大地理尺度上,植物往往由磷限制生长逐渐过渡到氮限制状态(Reichetal., 2004)。本研究发现,NSC库的大小随土壤的全氮含量和氮磷比增加而增加,而与全磷含量无关(表5)。这可能是因为马尾松生长环境土壤养分相对贫瘠,对额外资源采取更积极的利用方式,支持更高的生长率(Zadwornyetal., 2016)。除此之外,本研究发现气候因子对NSC库大小的影响高于土壤因子(图6)。相比于土壤,气候是树木生长空间变化的最强驱动力,并认为气候变化可能对森林生产力和固碳产生重大影响(Toledoetal., 2011),这支持本研究结果。本研究还发现,土壤对马尾松碳储存的影响主要通过与气候共同作用影响(图6)。在不同空间尺度上,土壤养分与气候条件的变化可能对树木生长产生复杂的相互作用(Gorissenetal., 2004)。如Baker等(2003)发现,在水分可利用性较高的情况下,半落叶性树木生长高于常绿森林,但这种差异仅在足够的降雨量情况下才明显。除气候和土壤外,林分结构、物种组成和植物多样性等因子均影响树木NSC的储存与分配(Tanakaetal., 2016; Chlumskaetal., 2022),但在本文中并未进行探讨,有待后续探索。
4 结论
通过在南北样带(23.0°—33.5°N)上对9个马尾松人工林的树木NSC及其组分库的大小估算认为,马尾松植株NSC库以淀粉储存为主,NSC库、可溶性糖库和淀粉库分别在干和根中存储最多; 整株和不同器官中的NSC库、可溶性糖库和淀粉库均呈现出显著的纬向变化格局; 气候对马尾松NSC存储及分配的影响大于土壤因子。
猜你喜欢
今日农业(2020年24期)2020-12-15
绿色科技(2019年5期)2019-11-29
测绘学报(2019年4期)2019-06-10
中国公共安全(2017年5期)2017-09-04
齐鲁周刊(2017年29期)2017-08-08
广西林业科学(2016年1期)2016-03-20
广西林业科学(2016年1期)2016-03-20
广西林业科学(2016年1期)2016-03-20
中国现代医学杂志(2015年26期)2015-12-23
