
烟草种子附生细菌群落结构与多样性分析
2022-12-20 05:58蔡刘体谢红炼刘文锋汪汉成宋光龙史彩华曹亚凡刘亭亭孟建玉
烟草科技 2022年11期
蔡刘体,谢红炼,2,刘文锋,汪汉成*,宋光龙,史彩华,曹亚凡,刘亭亭,2,孟建玉
1. 贵州省烟草科学研究院,贵阳市观山湖区龙滩坝路29 号550081
2. 长江大学农学院,湖北省荆州市荆秘路266 号 434025
3. 湖北中烟工业有限责任公司技术中心,武汉市东西湖区金山大道1355 号 430040
4. 贵州省烟草公司铜仁市公司,贵州省铜仁市碧江区锦江北路41 号 564100
5. 上海烟草集团有限责任公司,上海市杨浦区长阳路717 号西区 200082
烟草种子是烟草的重要器官,对烟草后代繁育具有重要作用。种子在发育与采收过程中,常携带包括病原菌在内的多种附生微生物。这些微生物通过附着在种子表面,直接或间接混杂于种子中[1],使带病种子成为烟草病害的初侵染源和传播载体。烟草种传病害包括真菌性、细菌性及病毒类病害,已报道的真菌性病害有烟草立枯病、烟草炭疽病等;病毒类病害有普通花叶病毒病、黄瓜花叶病毒病等[2-3];相比而言,烟草种传细菌性病害种类较少,但其危害严重,常见有烟草野火病和烟草细菌性角斑病等[4]。种传病害已成为种子生产加工、种衣剂研发的重点关注对象。
目前,烟草种子中有关细菌的相关报道较多,主要集中在内生细菌的研究方面。颜谨等[5]采用传统分离法研究发现K326 种子的内生细菌有假单胞菌属(Pseudomonas)、芽胞杆菌属(Bacillus)、贪铜菌属(Cupriavidus)等5 个属。Chen 等[6]利用高通量测序技术发现4个烟草品种(毕纳1、K326、PVH1452和云烟87)种子内生细菌的共有优势菌门为变形菌门和厚壁菌门。谢红炼等[7]试验发现云烟87、云烟85和K326 种子的内生优势细菌包括假单胞菌、埃希氏-志贺氏菌(Escherichia-shigella)等菌属。分离培养与高通量测序技术是种子微生物研究的常用手段,作者前期对种子附生和内生真菌[7-9]进行了初步研究。而针对烟草种子附生细菌的深入研究却鲜见报道[10]。为此,采用分离培养法对6个品种烟草种子附生细菌进行分离鉴定,并采用Illumina高通量测序技术对其中4个品种烟草种子的附生细菌群落结构与多样性进行分析,旨在进一步了解烟草种子附生细菌的种群结构,为烟草种子附生细菌的研究利用及种子病害防治提供依据。
1 材料与方法
1.1 材料
采用烟草裸种,品种为烤烟K326、云烟85、云烟87 和红花大金元,雪茄烟301,白肋烟L8,均由贵州省烟草科学研究院提供,并置于-80 ℃冰箱中保存。
NA培养基:称取蛋白胨10.0 g、牛肉浸粉3.0 g、NaCl 5.0 g、琼脂15.0 g,加入1 000.0 mL纯水中加热溶解,调节pH 为7.3 ± 0.1,置于121 ℃条件下高温灭菌20 min,备用。
1.2 可培养附生细菌分离与鉴定
1.2.1 可培养附生细菌分离
称取各品种的裸种50 g,分别置于盛有250 mL无菌水的三角瓶中,170 r/min摇床振荡培养2 h后过滤获得种子洗涤液,备用。吸取各品种的种子洗涤液1 mL,进行10 倍梯度稀释,将1×10-5、1×10-6和1×10-7倍的稀释液涂布于NA平板上,并置于28 ℃条件下黑暗培养2 d。用无菌接种环蘸取NA平板上不同颜色、形态和生长速率的单菌落[11],置于新的NA 平板上划线纯化,并将纯化后的细菌置于20%的甘油溶液中于-20 ℃保存。
1.2.2 可培养附生细菌鉴定
利用细菌鉴定通用引物27F(5'-AGAGTTTGAT CCTGGCTCA-3')和1492R(5'-GGTTACCTTGTTA CGACTT-3')对分离的细菌进行菌落PCR 扩增[12]。反应条件:94 ℃预变性3 min;94 ℃变性45 s,55 ℃退火45 s,72 ℃延伸90 s,30 个循环;72 ℃延伸7 min。2%琼脂糖凝胶检测PCR产物。扩增产物由赛默飞世尔科技有限公司测序,测序结果在NCBI上进行序列比对分析。
1.3 种子附生细菌群落结构与多样性测定
1.3.1 种子附生微生物DNA提取
选取1.2.1 节中K326、云烟85、L8 和301 的种子洗涤液,10 000 r/min离心10 min,弃上清液,获得种子附生微生物样品,每品种3 次重复。K326、云烟85、L8 和301 种子附生微生物样品编号分别为K326-1、K326-2、K326-3、Y85-1、Y85-2、Y85-3、L8-1、L8-2、L8-3和301-1、301-2和301-3。取各种子附生微生物样品0.50 g,采用E.Z.N.A.®Soil DNA Kit(美国Omega Biotek 公司)提取样品总DNA,用NanoDrop 2000[赛默飞世尔科技(中国)有限公司]检测其浓度和纯度。
1.3.2 16S rRNA文库构建及高通量测序
参照谢红炼等的方法[7],以各种子附生微生物样品基因组DNA 为模板,利用细菌通用引物779F(5'-AACMGGATTAGATACCCKG-3')和1193R(5'-ACGTCATCCCCACCTTCC-3')扩增各样品细菌16S rRNA基因的V5~V7区。反应体系(20 μL):5×FastPfu 缓冲液4 μL,dNTPs(2.5 mmol/L)2 μL,BSA 0.2 μL,上、下游引物(5 μmol/L)各0.8 μL,FastPfu聚合酶0.4 μL和DNA模板10 ng、ddH2O补足至20 μL。将合格扩增产物送至上海美吉生物医药科技有限公司进行16S rRNA 文库构建与扩增子测序(Illumina MiSeq PE300)。
1.3.3 数据处理
将测序数据进行质控分析,将生成的OTU 比对Silva 数据库(SilvaRelease132 https://www.arb-silva.de/documentation/release-132),计算多样性指数,并进行PCoA分析和聚类热图分析[13-14]。
2 结果与分析
2.1 种子附生可培养细菌分离与鉴定
从6 个烟草品种种子上共分离鉴定出可培养细菌35 株(表1),其中假单胞菌属(Pseudomonas)9 株(25.71%),芽胞杆菌属(Bacillus)7株(20%),肠杆菌属(Enterobacter)6 株(17.14%),泛菌属(Pantoea)4株(11.43%),鞘氨醇单胞菌属(Sphingomonas)2 株(5.71%),农杆菌属(Agrobacterium)2 株(5.71%),新鞘酯杆菌属(Novosphingobium)、寡养单胞菌属(Stenotrophomonas)、欧文氏菌属(Erwinia)、短小杆菌属(Curtobacterium)和金黄杆菌属(Chryseobacterium)各1株(2.86%)。
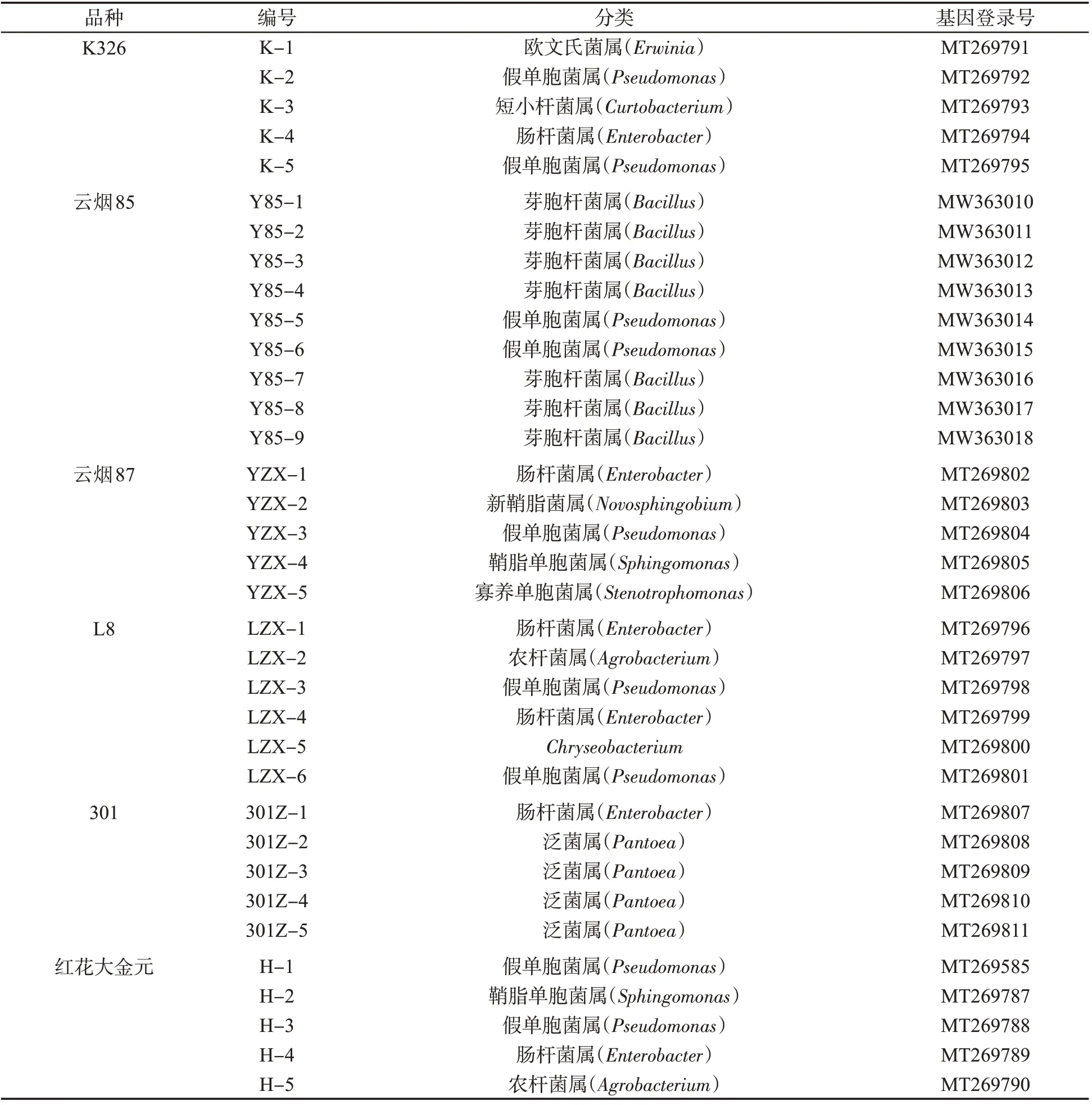
表1 6个烟草品种种子附生细菌的分子鉴定结果Tab.1 Molecular identification results of adnascent bacteria on seeds of six tobacco cultivars
K326种子上的附生细菌有假单胞菌属、欧文氏菌属、短小杆菌属和肠杆菌属;云烟85种子上的附生细菌有假单胞菌属和芽胞杆菌属;云烟87上的有假单胞菌属、肠杆菌属、新鞘脂杆菌属、鞘酯杆菌属和寡养假单胞菌属;L8种子上的有假单胞菌属、肠杆菌属、农杆菌属和金黄杆菌属;301 种子上的有泛菌属和肠杆菌属;红花大金元种子上的有假单胞菌属、鞘酯杆菌属、肠杆菌属和农杆菌属。
2.2 附生细菌群落结构与多样性分析
2.2.1 16S rRNA序列测序深度
4个品种的12个样品测序结果(图1)表明,随着测序深度的增加,OTU 水平下的香农指数稀释曲线趋于平缓,在测序序列数达到5 000 时,稀释曲线已饱和。12个样品共得到447 244条序列和200 011 454个碱基,所有序列的长度在266~559 bp 之间,平均长度为447 bp。
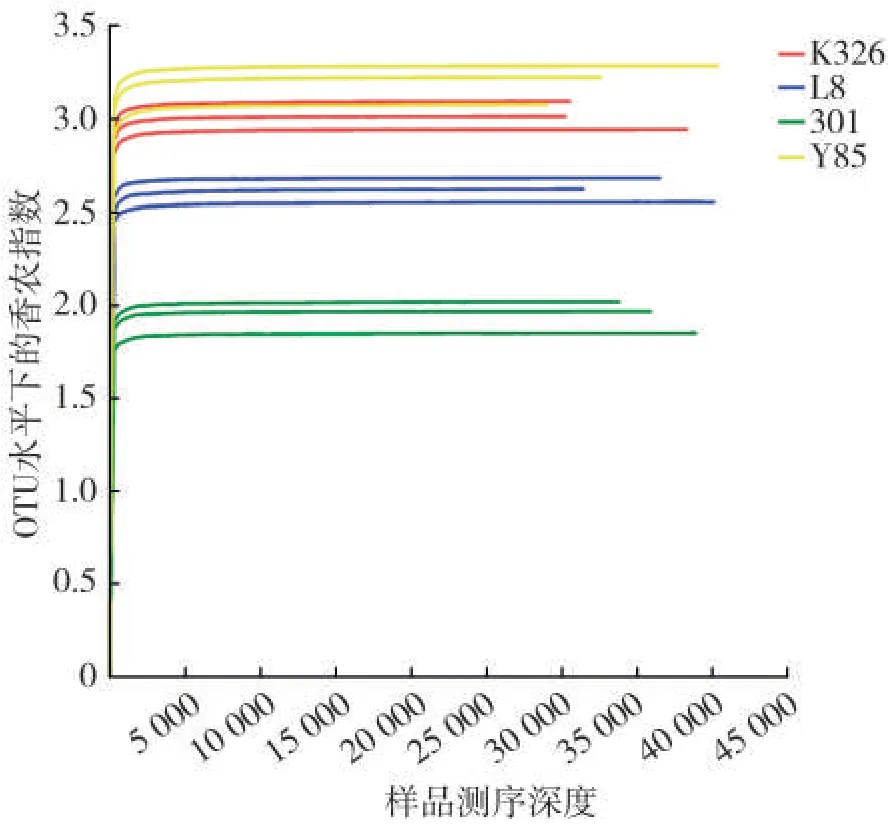
图1 稀释曲线(OTU水平Shannon指数)Fig.1 Rarefaction curve(Shannon index at OTU level)
2.2.2 烟草种子附生细菌群落多样性
4个烟草品种种子附生细菌共含有337个OTU,分别来自16 个菌门、26 个细菌纲、57 个细菌目、103个细菌科、182 个细菌属,共有263 个种。从OTU 分类水平来看,云烟85 种子上的附生细菌种类最多,为241 个;其次为L8(171 个);K326(165 个)和301(164 个)相对较少,见表2。4 个品种烟草种子附生细菌共有的细菌属和OTU 数量分别为48 和75 个,见图2。
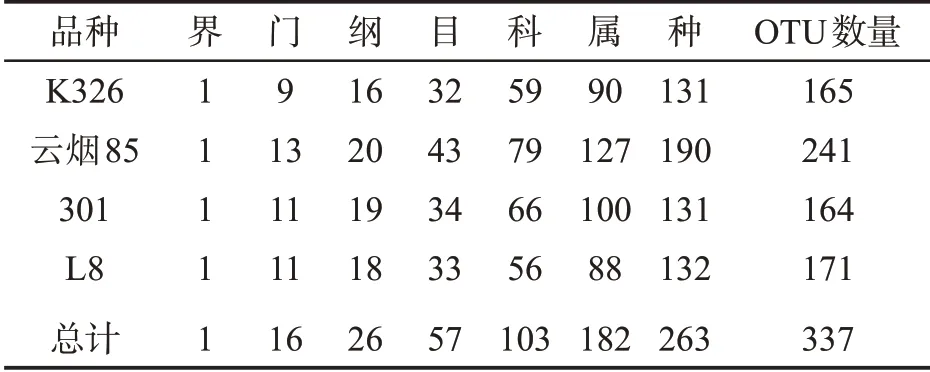
表2 4个品种烟草种子附生细菌不同分类水平数量Tab.2 Total amount of adnascent bacteria on seeds of four cultivars at different taxonomic levels (个)

图2 4个品种烟草种子附生细菌群落Venn图Fig.2 Venn diagram of adnascent bacterial communities on seeds of four tobacco cultivars
4个烟草品种种子附生细菌在丰富度指数、均匀度指数及多样性指数上均存在共性与差异(表3)。多样性大小排序依次为云烟85>K326>L8>301,且各品种间存在显著差异。丰富度指数(Sobs指数、Ace 指数、Chao 指数)表明4 个品种烟草种子附生细菌丰富度最大的为云烟85,最小的为301;K326 和L8种子附生细菌群落的丰富度差异不显著,云烟85和301 与其他品种种子附生细菌群落丰富度存在显著差异。4 个品种的均匀度和多样性指数也存在差异,相比而言,K326 种子附生细菌均匀度较高,而301的较低;云烟85的多样性指数相对较高。4个品种在覆盖度指数上差异不显著。
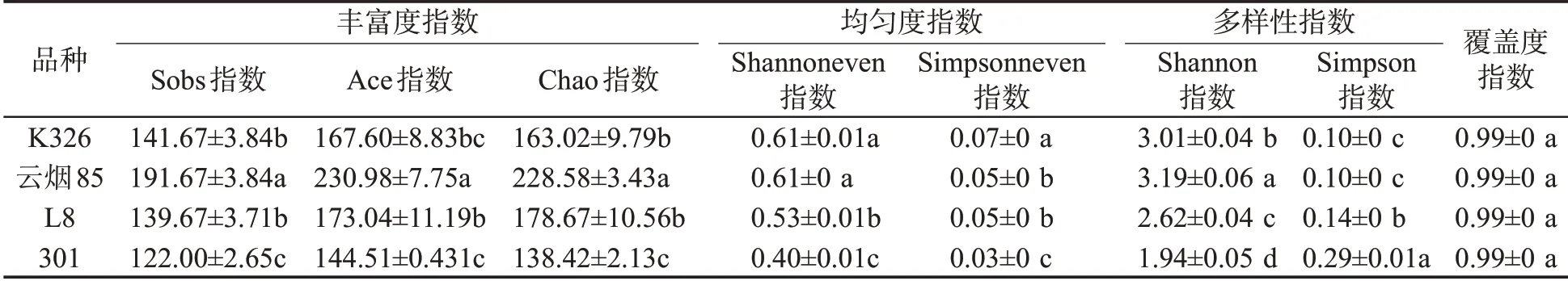
表3 4个品种烟草种子附生细菌Alpha多样性指数(OTU水平)①Tab.3 Alpha diversity indexes of adnascent bacteria on seeds of four tobacco cultivars at OTU level
2.2.3 烟草种子附生细菌群落结构
所有OTU序列比对Silva数据库显示,烟草种子附生细菌主要分布于变形菌门(Proteobacteria,74.12%~94.61%)、拟杆菌门(Bacteroidetes,0.43%~16.62%)、厚壁菌门(Firmicutes,0.98%~7.05%)、放线菌门(Actinbacteria,1.16%~5.39%)和蓝藻菌门(Cynobacteria,0.36%~3.17%),见图3。
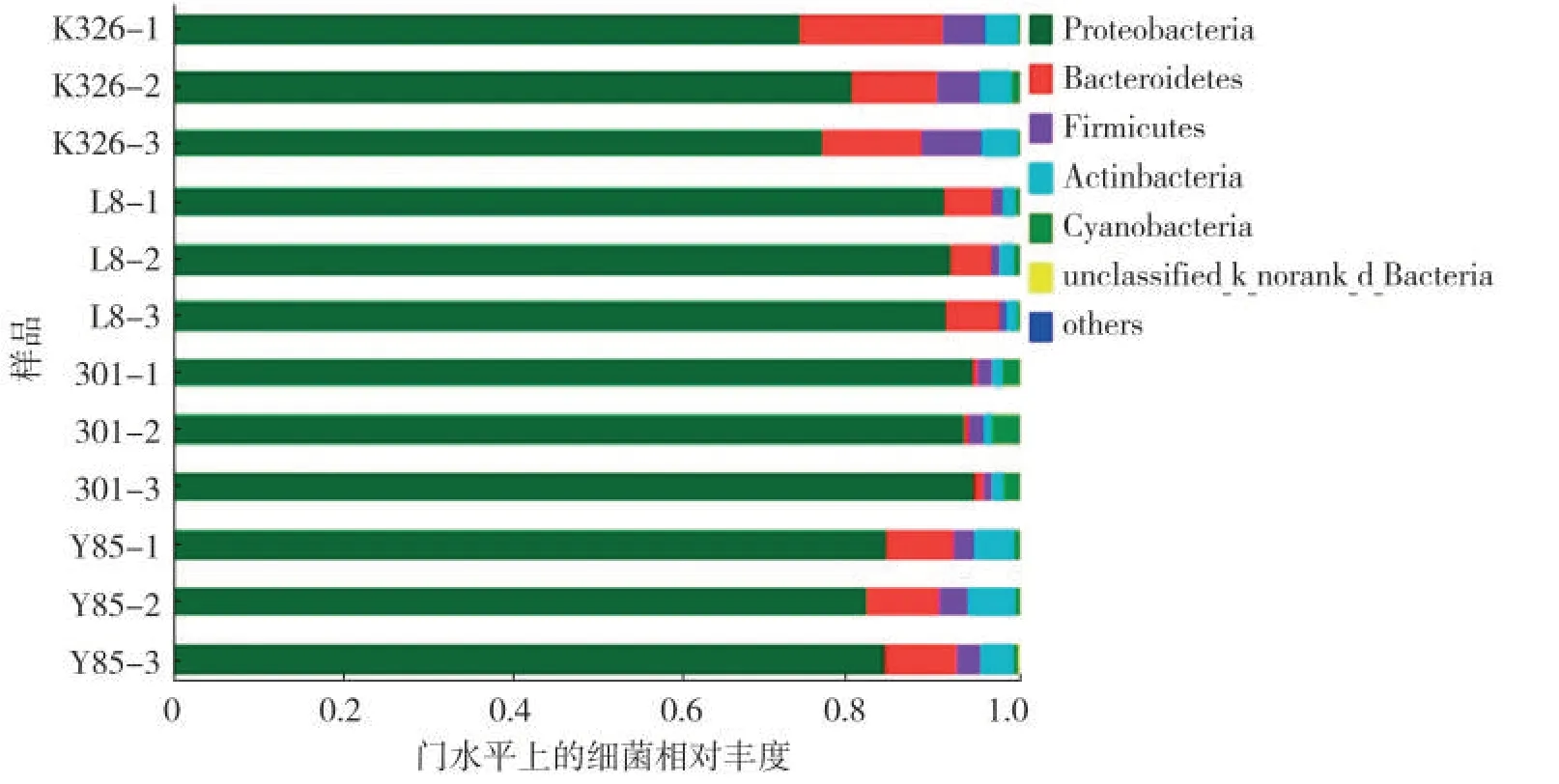
图3 4个烟草品种种子附生细菌在门水平上的相对丰度Fig.3 Relative abundances of adnascent bacteria on seeds of four tobacco cultivars at phylum level
4 个烟草品种种子附生细菌在属的组成上存在一定的相似性,其中K326、L8和云烟85的附生细菌的组成和丰度更为相似,301的附生细菌在丰度上与其他品种存在差异性(图4)。K326种子附生细菌群落中,假单胞属占32.22%,泛菌属占13.40%,鞘氨醇杆菌属(Sphingobacterium)占12.50%,肠杆菌属占9.87%,糖芽胞杆菌属(Saccharibacillus)占3.67%,寡养单胞菌属占3.62%,根瘤菌属占2.78%,苍白杆菌属(Ochrobactrum)占2.21% ,类芽胞杆菌属(Paenibacillus)占1.31%,代尔夫特菌属(Delftia)占1.27% ,Falsirhodobacter占 1.19% ,微杆菌 属(Microbacterium)占1.04%。L8 种子附生细菌群落中,假单胞菌属占27.60%,泛菌属占12.87%,寡养单胞菌属占8.76%,肠杆菌属占7.90%,根瘤菌属占3.55%,鞘氨醇杆菌属占2.84%,金黄杆菌属(Chryseobacterium)占2.33%。301种子附生细菌群落中,泛菌属占52.58%,假单胞菌属占24.25%,肠杆菌属占2.88%,寡养单胞菌属占1.05%。云烟85假单胞菌属占30.59%,泛菌属占13.57%,肠杆菌属占11.40%,鞘氨醇杆菌属占7.22%,根瘤菌属占3.93%,寡养单胞菌属占2.85%,微杆菌属占1.43%,苍白杆菌属占1.40%,无色杆菌属(Achromobacter)占3.01%,藤黄单胞菌属(Luteimnas)占1.51%,Falsirhodobacter占1.50%,葡萄球菌属Staphylococcus占1.33%,甲基杆菌属(Methylobacterium)占1.87%,新鞘氨醇杆菌属(Novosphingobium)占1.07%。此外,在4 个烟草品种种子样品中还检测到一些低丰度的细菌,如马赛菌(Massilia)、Ralstonia和Cellvibrio等。
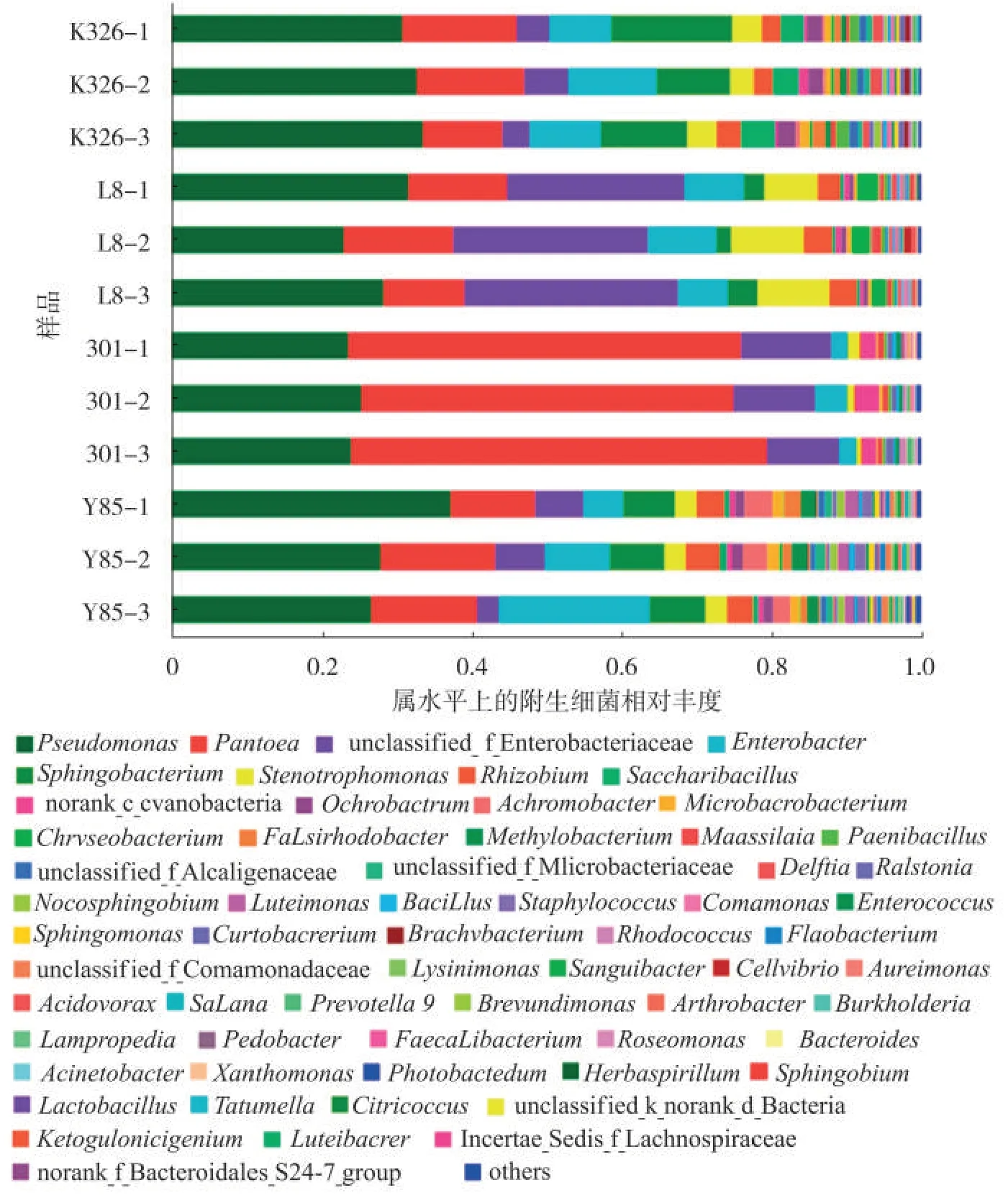
图4 4个品种烟草种子附生细菌在属水平上的相对丰度Fig.4 Relative abundances of adnsacent bacteria on seeds of four tobacco cultivars at genus level
2.2.4 种子附生细菌比较
基于Bray-Curtis距离算法对烟草种子附生细菌群落进行主坐标(PCoA)分析,结果(图5)表明,在OTU水平,品种K326和云烟85的样品聚在一起,而品种L8和301种子样品离其较远,且品种K326和云烟85种子附生细菌群落结构最为相似。PC1和PC2的贡献率分别为57.04%和22.53%,PC1 的贡献率明显大于PC2。表明PC1是影响烟草种子附生细菌群落结构差异的主要因素,PC1 的影响与K326、云烟85和L8种子附生细菌数量成正比,与301成反比,在PC1 的作用下,301 种子样品斑点远离其他种子,形成差异;PC2的影响与K326、云烟85和301种子附生细菌数量成正比,与L8成反比,在PC2的作用下,L8种子附生细菌样品斑点远离其他种子,也形成差异。
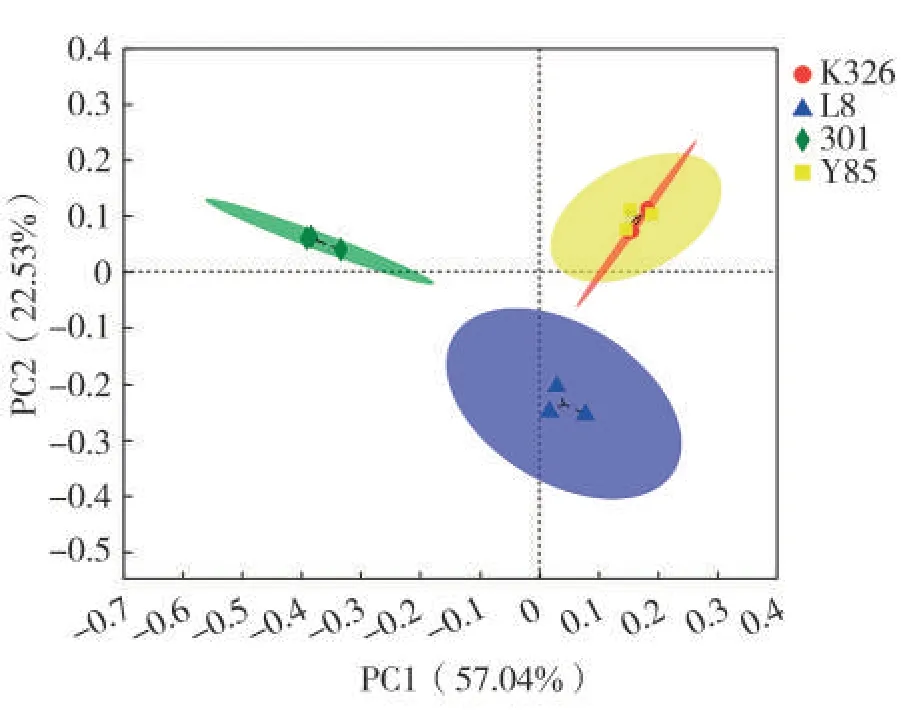
图5 OTU水平烟草种子附生细菌主成分分析Fig.5 Principal component analysis of adnsacent bacteria on tobacco seeds at OTU level
在属水平,对4个品种种子样品附生细菌按丰度进行排序,选取排名前30 的细菌绘制丰度热图(图6)。聚类热图结果表明,每个品种种子的3个样品单独聚为一支,说明样品重复性好,品种K326 与云烟85种子样品聚为一个分支,表明品种K326和云烟85种子的附生细菌群落最为相似,与主坐标分析结果一致。聚类树分析表明,假单胞属、泛菌属和在属水平未分类的肠杆菌科细菌聚为一支,肠杆菌属、鞘酯假单胞菌属、寡养单胞菌属和根瘤菌属聚为一支。各品种种子上,假单胞属、泛菌属的细菌最多,其次为肠杆菌属、鞘酯假单胞菌属、寡养单胞菌属和根瘤菌属细菌。
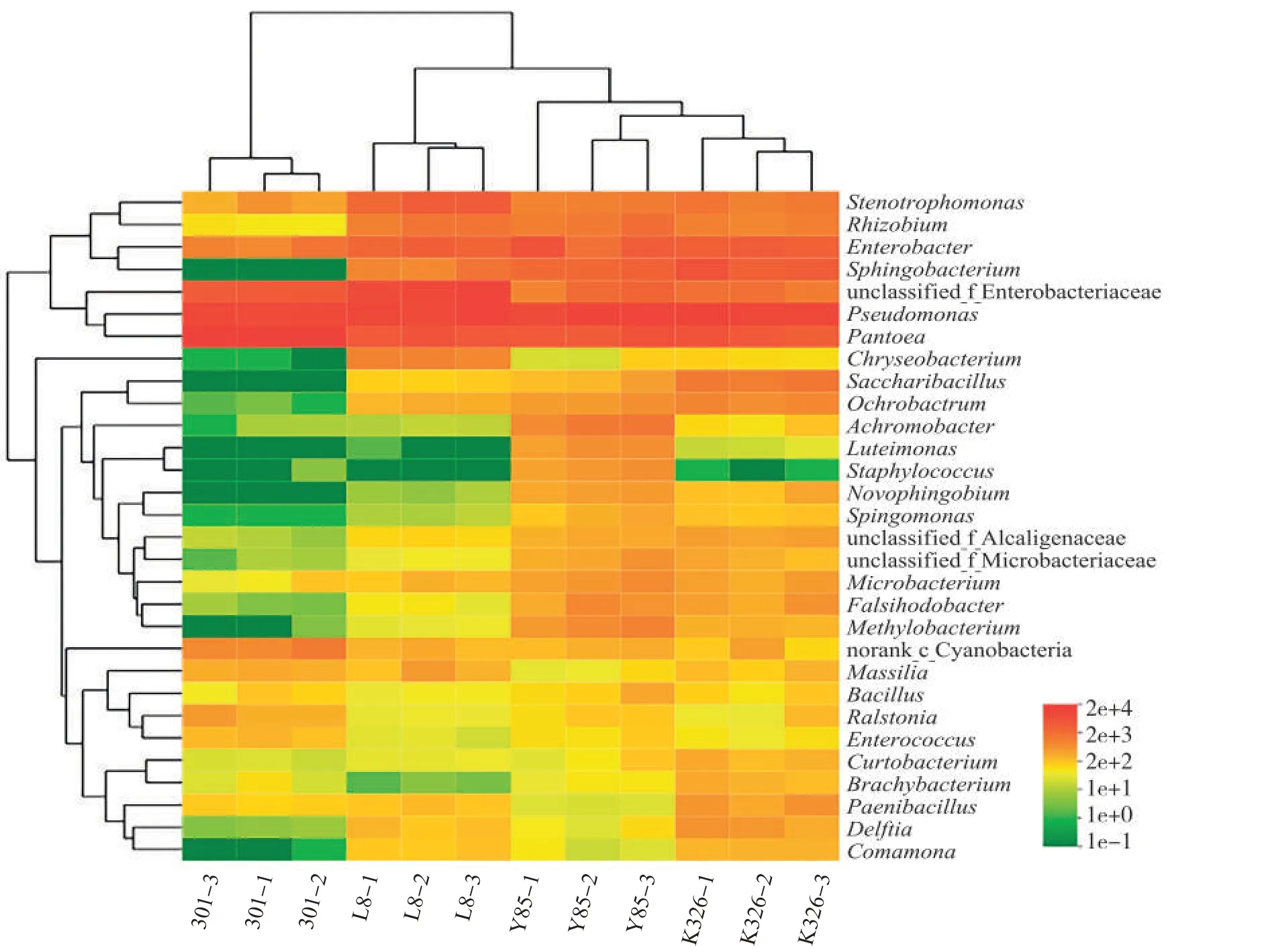
图6 属水平下4个品种烟草种子附生细菌的相对丰度Fig.6 Relative abundances of adnsacent bacteria on seeds of four tobacco cultivars at genus level
3 讨论
本试验中高通量测序发现,12 个样品的覆盖度指数均达到99.9%,且在测序序列数量内,稀释曲线完全达到饱和,表明测序结果足以反映所研究的烟草种子附生细菌群落生物信息。同时,相对丰度热图上方的聚类树和PCoA 的结果均发现每个品种的3个样品间差异较小,表明本试验中的取样排除了较大的偶然性误差。
高通量测序结果显示4 个烟草品种种子附生细菌均分布于变形菌门、拟杆菌门、厚壁菌门,其中含量最高的均为变形菌门。与3个烟草品种(K326、云烟85、云烟87)种子内生细菌在门水平上的优势种群结果一致[7]。在属水平上,4个品种丰度排名前10的附生细菌均分别是假单胞属、泛菌属、鞘氨醇杆菌属、肠杆菌属、糖芽胞杆菌属、寡养单胞菌属、根瘤菌属、苍白杆菌属、无色杆菌属、微杆菌属和金黄杆菌属。相比而言,烟草种子内生细菌的优势菌属为假单胞菌属、大肠杆菌—志贺菌属、细杆菌属、Lelliottia、鞘脂杆菌属、糖芽胞杆菌属、白色杆菌属、寡养单胞菌属等[7]。烟草种子附生细菌与内生细菌在群落结构上存在较大差异,但也存在共有优势的菌属假单胞属、糖芽胞杆菌属、寡养单胞菌属等细菌,表明这些细菌为烟草种子内、外共生细菌;同时也发现种子优势细菌泛菌属为非种子内生菌。此外,本研究中还发现在种子附生细菌多样性方面,烤烟云烟85和K326均高于雪茄烟301和白肋烟L8,这可能与烟草种子的种植环境、来源、采收季节不同等有关,同时也可能是由于烟草品种不同引起的差异,均有待通过同地、同季、同环境的比较试验进一步验证。
本试验中发现烟草种子可培养附生细菌有假单胞菌属、肠杆菌属、鞘酯假单胞菌属、寡养假单胞菌属、泛菌属、新鞘酯假单胞菌属、欧文氏菌属、短小杆菌属、芽胞杆菌属、金黄杆菌属和农杆菌属,其丰度较高细菌种类与高通量测序结果基本一致,说明传统分离法也能反映种子优势种群的群落结构和相对丰度。假单胞菌属为烟草种子附生和内生细菌中种群丰度均最高的细菌类群,已有的研究发现,由丁香假单胞烟草致病变种(Pseudomonas syringaepv.tabaci)引起的烟草野火病[15]和丁香假单胞杆菌角斑专化型(Pseudomonas syringaepv.angula)引起的烟草细菌性角斑病[16]为烟草上两种主要的细菌性病害,本试验中检测到的假单胞菌属细菌中是否含有这两种致病菌,仍有待进一步通过致病力检验来明确种子携带烟草致病菌的风险。此外,对于烟草种子的其他附生细菌,有研究发现泛菌属中的菠萝泛菌(Pantoea ananatis)和分散泛菌(Pantoea dispersa)均会引起水稻叶枯病[17],成 团泛菌(Pantoea agglomerans)会引起红枣坏死[18];肠杆菌属中的阴沟杆菌(Enterobacter cloacae)会引起洋葱球和生姜根茎的软腐病[19-20],桑肠杆菌(Enterobacter mori)会引起桃类果实的采前软腐病[21];类芽胞杆菌属中的多黏类芽胞杆菌(Paenibacillus polymyxa)会引起火龙果的细菌性茎腐病[22]。本试验中检测到的烟草种子附生细菌中是否也存在这些菌属的烟草致病菌还有待通过致病力试验进行验证。
4 结论
①6 个烟草品种种子上共培养分离鉴定出细菌35株,其中假单胞菌属9株,芽胞杆菌属7株,肠杆菌属6株,泛菌属4株,鞘氨醇单胞菌属2株,农杆菌属2 株,新鞘酯杆菌、寡养单胞菌、欧文氏菌、短小杆菌和金黄杆菌属各1株。②K326、白肋烟L8和云烟85种子的附生细菌组成和丰度相似,多样性排序依次为云烟85>K326>L8>301,雪茄烟301种子附生细菌在丰度上低于其他品种;4个烟草品种种子附生细菌共有的优势菌属有假单胞属、泛菌属、鞘氨醇杆菌属、肠杆菌属、糖芽胞杆菌属、寡养单胞菌属和根瘤菌属等。
猜你喜欢
河南医学研究(2022年19期)2022-10-19
农业技术与装备(2020年10期)2020-11-03
生态学报(2019年11期)2019-07-08
中成药(2017年9期)2017-12-19
中华老年口腔医学杂志(2016年2期)2017-01-15
河北书画研究(2016年6期)2016-02-03
诗歌月刊(2014年3期)2015-04-14
传奇故事(破茧成蝶)(2015年8期)2015-02-28
现代检验医学杂志(2015年1期)2015-02-06
食品工业科技(2014年23期)2014-03-11
