
基于DNA条形码的江门近岸海域春季鱼卵的种类鉴定及其形态
2022-12-20 07:28:46黄德练王雪辉李亚芳宁加佳王亮根刘双双林昭进杜飞雁
南方水产科学 2022年6期
陈 静 ,黄德练,王雪辉,徐 磊,张 健,李亚芳,宁加佳,王亮根,刘双双,林昭进,杜飞雁
1. 上海海洋大学 海洋科学学院,上海 201306
2. 中国水产科学研究院南海水产研究所/广东省渔业生态环境重点实验室,广东 广州 510300
鱼卵是鱼类的早期发育阶段,掌握其丰度和分布对了解鱼类早期资源变动具有重要意义[1]。鱼卵的形态分类和鉴定是早期渔业资源种类调查和评估的重要依据[2-4]。南海作为我国三大陆缘海之一,具有独特的半封闭海域条件,是世界上海洋生物种质资源最丰富的区域之一,已记录鱼类3 400多种,然而有形态信息和分类研究的鱼卵种类却极其缺乏[5-6]。鱼卵的发育时间短、形态变化大、相近种类特征相似,在鱼类的产卵期与产卵场相互交叠的情况下难以准确对其进行分类研究[5],限制了对近海鱼类早期资源的准确评估和深入研究。随着GenBank (www.ncbi.nlm.gov) 和BOLD systems(v4.boldsystems.org) 等数据库的不断完善,DNA条形码技术在鱼卵的分类鉴定中应用广泛,为解决鱼卵形态学鉴定的研究瓶颈提供了新思路和手段。DNA条形码技术在进行鱼卵仔鱼鉴定时,不受限于种类形态、状态及发育时期,仅依靠少量的组织就能获取该种类的基因序列,经扩增、测序及序列比对即可获得较为准确的种类鉴定结果,使鱼卵形态特征和分类研究更加准确、快捷[2,7-8]。
珠江口地处亚热带咸淡水交汇海域,常年受珠江冲淡水、沿岸流及上升流的综合影响,是我国最富生产力的河口之一[9-11],孕育了丰富的渔业资源,也是近海鱼类重要的产卵场[10,12]。江门位于珠江口的西南部,是南海海域的典型河口区域之一,其沿岸的黄茅海河口、广海湾及其邻近海域 (中华白海豚保护区),受珠江口和南海海域的综合作用,孕育了丰富的渔业资源[13-14],成为鲾科、科、舌鳎科和鲱科等多种小型鱼类的产卵场和育幼场。本文以珠江口江门海域现场采获的鱼卵种类为样本,在DNA条形码准确鉴定种类的基础上,描述鱼卵的主要形态特征并探讨其形态分类,以期为今后珠江口渔业资源的保护和管理提供参考。
1 材料与方法
1.1 样品采集及处理
鱼卵样品于2021年4月在广东珠江口江门海域的7个采样点采集获得。每个站位以大型浮游生物网 (网口内径0.8 m、网长2.8 m,网口面积0.5 m2)在水体表层进行水平拖网采样,每个站位拖网1次,每次10 min,采样完成后收集网底管内样品充氧保存以保持活体状态。所有站位样品在采集后充分混合,取1/7作为抽样样本。体视显微镜(Nikon SMZ800N) 下随机挑出3 245 颗鱼卵,参考鱼卵形态分类指南[1],进行初步的形态分类。按比例共计选取了136 颗鱼卵[15],体视显微镜下对其进行拍照和形态记录。鱼卵样品记录形态后逐一放入1.5 mL无菌离心管中,以体积分数为95%的乙醇固定,−20 ℃保存以进行后续分子生物学实验。
1.2 DNA提取和PCR扩增
鱼卵样品DNA提取使用海洋动物组织基因组提取试剂盒 (天根)。使用引物FishBCL (5'-TCAACYAATCAYAAAGATATYGGCAC-3') 和FishBCH(5′-ACTTCYGGGTGRCCRAARAATCA-3')[16]对线粒体细胞色素C第I亚基 (COI) 目的片段进行PCR扩增。30 μL PCR反应体系组成为TaqDNA聚合酶(TaKaRa) 0.3 μL,dNTP 3 μL,10× Taqbuffer 3 μL,正反向引物各0.9 μL,DNA模板3 μL,ddH2O补足。反应条件为95 ℃预变性5 min;94 ℃变性45 s,52 ℃退火30 s,72 ℃ 延伸1 min,35个循环;72 ℃延伸10 min。PCR扩增产物经质量分数为1%琼脂糖凝胶电泳检测后送广州天一辉远基因科技公司进行纯化和序列双向测定。
1.3 序列分析和种类确认
利用DNAstar 7.1软件包 (DNASTAR, Inc.) 对测序获得的原始序列进行检查、拼接及校对,确定序列准确性,并将获得的序列与GenBank及BOLD systems数据库进行对比。依据遗传相似度>99%认定为种水平、92%~99%为属水平,以此为基准对鱼卵进行种类判定[17]。从NCBI数据库下载与样品序列同源性达到种水平的COI基因序列。使用MEGA 7.1软件对所有的COI基因序列进行多重序列比对,以斑节对虾 (Penaeus monodon)作为外系群。采用基于K2P (Kimura two parameter model) 遗传距离的邻接法 (Neighbor-Joining, NJ) 构建系统进化树[18]以最终确认鱼卵所属的种类或类群,Bootstrap置信值设置为重复抽样1 000次。校验后的序列上传至GenBank。
2 结果
2.1 种类组成
本研究共获得86个样品的测序结果,序列长度为628~684 bp。通过与基因数据库比对分析,发现其COI序列与两个数据库中参考序列比对均能达到99%~100%的种水平,且在种鉴定水平范围内出现相同的1~3个种类。初步判定13个鱼卵种类分属于3目10科10属;其中4种鱼卵能鉴定至种,分别为缘边银鲈 (Gerres limbatus)、多鳞(Sillago sihama)、眶棘双边鱼 (Ambassis gymnocephalus) 及斑头舌鳎 (Cynoglossus puncticepsc);8种鉴定至属,分别为沙丁鱼属、小公鱼属、棘鲷属、副叶鲹属、叫姑鱼属、属、舌鳎属-1和舌鳎属-2;1种鉴定至科,为鲾科 (表1)。构建的NJ系统进化树,所有物种均能成功进行种属的聚类 (图1)。
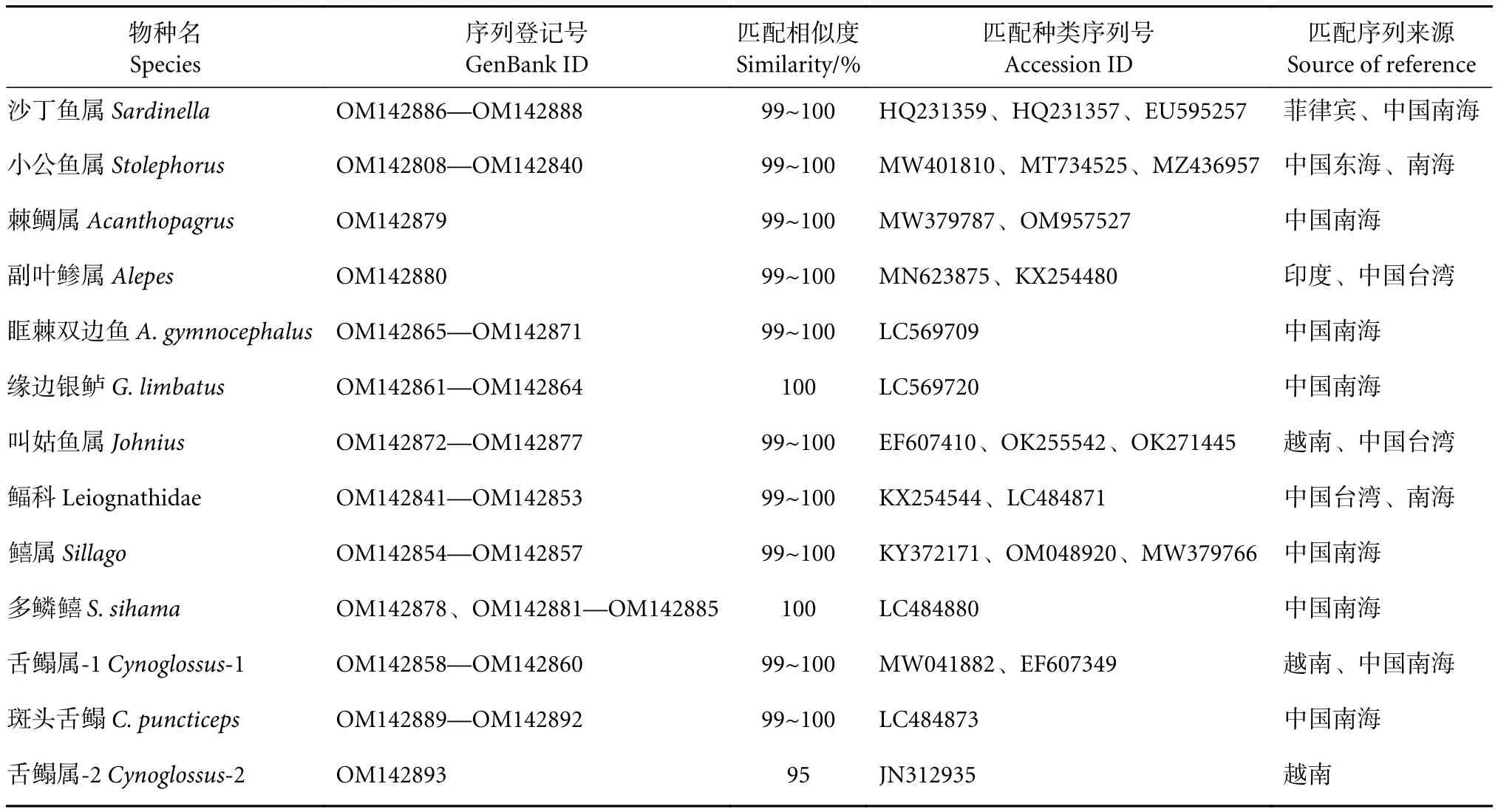
表1 研究海域鱼卵DNA序列对比及物种鉴定结果Table 1 Results of DNA sequence comparison and species identification of fish eggs in survey area
分子鉴定的种类组成结果中,以鲈形目最多,其次为鲽形目,鲱形目再次之。个体数量以小公鱼属最多,占总个体数的38.4%;鲾科次之 (15.1%);眶棘双边鱼再次之 (8.1%)。结合图像信息,发现相同种类鱼卵所处的发育阶段一致。其中沙丁鱼属鱼卵处于刚完成受精不久的受精卵时期,胚体尚未形成;舌鳎属和叫姑鱼属鱼卵处于卵裂期和囊胚期;其余鱼卵种类均处于胚体和器官形成时期。
2.2 形态特征
沙丁鱼属卵为圆球状 (图2-a),分离浮性;卵膜薄、光滑透明,无特殊构造;卵径介于1.563~1.615 mm,均值为 (1.588±0.024) mm;卵周隙介于0.337~0.458 mm,均值为(0.385±0.051) mm,约为卵径的47%~53%。卵黄近圆形,直径介于0.836~0.870 mm,均值为(0.850±0.012) mm,分两层,外层半透明,颜色较浅,内层呈乳白色兼有不规则的龟裂。单油球,呈浅黄色,位于卵黄囊中央,直径介于0.131~0.141 mm,均值为 (0.135±0.004)mm;油球表面光滑,未见明显色素胞分布。
小公鱼属卵为椭圆状 (图2-b),分离浮性;卵膜薄、光滑透明,无特殊构造,具有弹性;卵周隙较小;卵黄囊也呈椭圆形,有不规则龟裂;长径介于1.177~1.189 mm,均值为 (1.182±0.005) mm,短径介于0.684~0.692 mm,均值为 (0.689±0.003) mm;此时胚体形成,孵化出膜围绕卵黄囊3/5,胚体后端1/3形成尾部,已孵化出膜;橙色油球一个,靠近胚体尾部1/3处,直径介于0.095~0.101 mm,均值为 (0.098±0.002) mm,油球表面光滑,未见明显色素胞分布。
棘鲷属卵为圆球状 (图2-c),分离浮性;卵膜薄、光滑透明,无特殊构造;卵径介于0.772~0.780 mm,均值为 (0.776±0.003) mm,动、植物极卵周隙变宽;此时胚体形成,孵化出膜围绕卵黄囊3/5,视囊内晶体形成,听囊出现;胚体自头到尾沿身体两侧布满褐色色素胞,胚体末端形成尾芽,紧附卵黄囊,向内弯曲伸长;单油球,靠近胚体尾部1/3处,直径介于0.158~0.168 mm,均值为 (0.163±0.004) mm,油球表面零星出现点状黑色素胞。

图2 珠江口江门海域 13种鱼卵不同发育阶段的图像注:a. 小沙丁鱼属;b. 小公鱼属;c. 棘鲷属;d. 副叶鲹属;e. 眶棘双边鱼;f. 缘边银鲈;g. 叫姑鱼属;h. 鲾科;i. 属; j. 多鳞;k. 舌鳎属-1;l. 斑头舌鳎;m. 舌鳎属-2;比例尺均为 0.1 mm。Fig. 2 Photographs of different embryonic development stages of fish eggs of 13 species from Jiangmen coastal area of Pearl River EstuaryNote: a. Sardinella sp.; b. Stolephorus sp.; c. Acanthopagru sp.; d. Alepes sp.; e. A. gymnocephalus; f. G. limbatus; g. Johnius sp.; h. Leiognathidae;i. Sillago sp.; j. S. sihama; k. Cynoglossus sp.-1; l. C. puncticeps; m. Cynoglossus sp.-2. Scale bar is 0.1 mm.
副叶鲹属卵为圆球状 (图2-d),分离浮性;卵膜薄、光滑透明,无特殊构造;卵径为0.644~0.659 mm,均值为 (0.650±0.005) mm;未见明显卵周隙;此时观察到胚体已具雏形,一端逐渐膨大拱起,形成胚体前端,另一端沿着卵膜延伸逐渐形成胚体尾端,胚体两端靠近卵膜一侧零星分布有黑色素胞;单油球,直径介于0.136~0.148 mm,均值为 (0.142±0.004) mm,油球表面零星分布有黑色素胞。
眶棘双边鱼卵为圆球状 (图2-e),分离浮性;卵膜薄、光滑透明,无特殊构造;卵径介于0.716~0.724 mm,均值为 (0.720±0.003) mm;未见明显的卵周隙;卵黄颗粒细且均匀,略显现乳白色;此时胚体出现雏形紧密围绕着油球,胚体外侧规则排列有2~3列点状黑色素胞,胚体一端正在形成头部,视囊隐约可见;单油球,直径介于0.157~0.169 mm,均值为 (0.161±0.005) mm,油球表面可观察到5~6个点状黑色素。
缘边银鲈是短棘银鲈(G. lucidus)的同物异名(图2-f)。卵为圆球状,分离浮性;卵膜薄、光滑透明,无特殊构造;卵径介于0.716~0.732 mm,均值为 (0.725±0.006) mm;此时胚体已形成,环绕卵黄囊2/3,鱼卵原口已关闭;胚体前端形成头部,头部膨大形成具有分化的脑部;脑部视囊形成;未见明显晶体形成;视囊后方出现听囊;耳石未形成;胚体尾部尾芽雏形初现;胚体头部、躯干部及尾部均零星分布褐色色素胞,在胚体背部分布较密集且呈线形规则分布;单油球,直径介于0.155~0.164 mm,均值为 (0.160±0.003) mm,表面零星分布有色素胞。
叫姑鱼属卵为圆球状 (图2-g),分离浮性;卵膜薄、光滑透明,无特殊构造;卵径介于0.755~0.770 mm,均值为 (0.761±0.007) mm;卵周隙较窄;此时为刚受精不久的鱼卵,原生质向动物极集中,形成狭窄的卵周隙,形成白色帽状胚盘;单油球,呈乳白色;位于卵黄囊中央,直径为0.189~0.198 mm,均值为 (0.192±0.003) mm,油球表面光滑,未见明显色素胞分布。
鲾科卵为圆球状 (图2-h),分离浮性;卵膜薄、光滑透明,无特殊构造;卵径介于0.650~0.673 mm,均值为 (0.663±0.010) mm;卵周隙较窄;卵黄囊透明,表面无明显龟裂;胚胎环绕卵黄囊3/5时期,视囊内晶体形成,听囊出现;胚体出现星状黄色素胞沿中央两侧成行列式规则分布至尾端;胚体末端形成尾芽,已离开卵黄并向内弯曲伸长;单油球,靠近胚体尾部1/3处,直径介于0.156~0.160 mm,均值为 (0.155±0.002) mm,表面分布星状黄色色素胞且夹杂有黑色点状色素胞。
舌鳎属-1为受精不久的鱼卵 (图2-k)。卵为圆球状,分离浮性;卵膜薄、光滑透明,无特殊构造;卵径介于0.843~0.865 mm,均值为 (0.849±0.010) mm;未见明显的卵周隙;油球10个左右,大小各异,直径介于0.042~0.096 mm,聚集紧密,油球表面光滑,未见明显色素胞分布;与油球相对的动物极出现多层分裂球叠加排列形成细胞团,两侧微隆起呈小丘状,整个细胞团界限模糊。
斑头舌鳎为受精不久的鱼卵 (图2-l),受精孔清晰可见。卵为圆球状,分离浮性;卵膜薄、光滑透明,无特殊构造;卵径介于0.784~0.806 mm,均值为 (0.793±0.008) mm;未见明显的卵周隙;卵黄颗粒细匀;团状原生质集中于油球相对侧动物极,形成帽状胚,胚盘处的分裂球持续分裂聚集成囊胚状,油球16个左右,大小各异,直径范围为0.045~0.073 mm,聚集紧密,表面光滑,未见明显色素胞分布。
舌鳎属-2卵为圆球状 (图2-m),分离浮性;卵膜薄、光滑透明,无特殊构造;卵径介于0.733~0.739 mm,均值为 (0.734±0.003) mm;卵黄颗粒细匀,呈现半透明状态;此时胚体已完全形成,鱼卵原口已关闭,胚体纤长环绕卵黄囊1/2时期;神经管发育完善,沿胚体中轴向前延伸至吻端;胚体前端膨大形成分化的头部,紧紧依附在卵黄囊上,吻端钝圆;出现黑色素沉淀;视囊后方形成长椭圆形听囊,左右各1个;耳石未形成;视囊形成;油球16个左右,大小各异,直径介于0.045~0.075 mm,彼此分离分布在胚体中后部,表面光滑,未见明显色素胞分布。
2.3 形态分类
本文采用形态学方法和DNA条形码技术相结合的研究方法,对江门海域采获的13种鱼卵进行了种属水平的种类鉴定。采获的13种鱼卵均为浮性卵,卵膜单层、光滑透明,无特殊构造。形态特征的差异主要体现在卵膜形状,卵径大小,油球数量、分布及其色素分布,卵周隙宽窄,胚体上色素胞的颜色、数量及分布等。可分别依据以上特征大致将鱼卵种类进行形态分类。首先是卵径形状:小公鱼属鱼卵呈椭圆形,长径明显大于短径,其他种类为圆球形。其次是卵径大小:沙丁鱼属鱼卵卵径最大,为1.563~1.615 mm,明显大于其他鱼卵卵径 (0.644~0.873 mm);副叶鲹属卵径最小,为0.644~0.659 mm。种属间相近种类的卵径变化相对较小,难以进一步区分,但鱼卵油球数量、分布及其色素分布具有一定差异。舌鳎属鱼卵油球数量较多且大小不一,表面无明显色素分布,呈聚集或分散分布。其中舌鳎属-1油球10个左右,聚集紧密,直径介于0.042~0.096 mm;斑头舌鳎油球16个左右,聚集紧密,直径介于0.045~0.073 mm,聚集紧密;舌鳎属-2油球16个左右,直径介于0.045~0.075 mm,散布在胚体尾部;除舌鳎属外其他种类鱼卵均为单油球。此外,油球表面色素分布也可以进一步辅助形态区分,如沙丁鱼属、小公鱼属及叫姑鱼属油球均无明显色素分布,属油球表面黄色色素胞较为密集,其他种类油球均有不同程度的黑色或黄色色素胞分布。再者是卵周隙宽窄:沙丁鱼属、眶棘双边鱼鱼卵的卵周隙均较宽,其他种类鱼卵卵周隙均较窄。对于已有胚体形成的鱼卵,胚体上色素胞颜色、数量及分布也是种类鉴定的重要依据。棘鲷属胚体体干两侧及背部明显分布有黄色素胞;副叶鲹属胚体两端规则分布有行列式黑色素胞。
3 讨论
3.1 形态分类结合DNA条形码在鱼卵分类中的应用
形态学鉴定作为早期鱼类种类鉴定常规且普遍的研究方法,其鉴定过程复杂烦琐,受限于分类专家的主观判断和仪器设备的制约[19]。Ko等[17]借助DNA条形码技术对5个实验室仔鱼形态鉴定结果的准确性进行检查,摒弃未知种类,形态学方法鉴定至属水平的准确率为75.4%,而种水平仅为43.7%,认为形态学方法仅能将鱼类早期阶段鉴定到科属水平。DNA条形码技术可较准确地将早期鱼类鉴定到种水平,形态分类与DNA条形码结合显著提高了种类鉴定的准确性,已普遍应用于早期鱼类研究中。陈永久等[20]和周美玉等[21]研究发现DNA条形码技术与形态学鉴定相结合对于准确鉴定鱼卵、仔稚鱼非常有效,特别是对鱼卵的鉴定。Hou等[22]和Becker等[23]利用分子生物学方法对相关海域的鱼卵进行种类鉴定,发现DNA条形码能将形态学分类结果鉴定至更低阶元,证实了DNA条形码技术在鱼卵物种识别中的可行性。然而单一的DNA条形码分析方法具有局限性,在种类鉴定和物种归属等问题上仍有争议。Labrador等[24]和Sukumaran等[25]使用两个mtDNA标记高变控制区和COI基因序列对沙丁鱼属鱼类进行系统发育分析,发现沙丁鱼属鱼类为单倍体群体,遗传分化水平较高,COI基因序列无法识别该属中的隐种和种群结构,需要借助高度特化的核DNA标记进一步分析。本文仅借助COI基因序列可能无法对沙丁鱼属进行种水平的准确划分,为谨慎起见,仅保留鉴定至属的分类结果。近年来,陆续发表的一些基因片段如12S rRNA、16S rRNA和Cytb等均可用于鱼类种类鉴定研究[26-27]。因此,在形态分类结合DNA条形码的基础上,要实现对鱼卵准确的物种鉴定,还需要结合更多的序列片段分析,以提高种类鉴定及早期鱼类形态判别的准确性。此外,准确性高、完整性强及基因片段丰富的成鱼数据库是序列分析鉴定种类的前提。南海DNA条形码数据库的建立[28],使珠江口及近岸海域鱼卵种类鉴定的准确性得到了提高。Hou等[29]使用DNA条形码和形态学共鉴定出珠江口春季和秋季的鱼卵仔稚鱼75个分类群,初步构建了珠江口鱼卵仔稚鱼的形态数据库。本文同样依托公开数据库,探讨江门海域春季鱼卵的形态特征及分类。
3.2 江门海域春季鱼卵种类的形态及其初步分类探讨
鱼卵发育早期的种类鉴定主要以观察其内部形态为主,本文采获的鱼卵样品最明显的分类特征在于鱼卵形状、卵径、油球、卵周隙等。我国海域广泛分布有鲱科和鳀科鱼类,种类数量均超过20种,其中记录在册的鲱科、小公鱼属鱼类多达9种[30]。小公鱼属和鳀属同属于江门海域常见种类[18,31],鱼卵形状均为椭圆形,其早期鱼卵形态及胚胎发育变化极为相似,有无油球是两者最明显的鉴定特征[32]。对鱼卵样品进行形态学观测,卵径范围为0.658~1.615 mm,油球径范围为0.042~0.193 mm,13种鱼卵的发育时期存在差异,致使测量值有一定程度的重叠,不能作为鱼卵分类的准确依据。沙丁鱼属鱼卵最明显的分类特点是卵径较大、卵黄囊较小、卵周隙宽,本文获取的沙丁鱼属鱼卵卵径、卵黄囊及卵周隙大小均与赵孝威等[33]研究的青鳞沙丁鱼 (Sardinella zunas) 早期发育阶段的形态相契合,但油球尺寸与其差异较大,现有的形态学资料无法鉴定到种。舌鳎属鱼卵的结构特征具有一定的相似性[34],舌鳎属鱼卵的显著特征为多油球,根据万瑞景等[35]得出的鱼卵形态特征,可初步判定3种鱼卵皆为舌鳎属鱼卵。叫姑鱼属隶属石首鱼科,在《中国近海鱼卵与仔鱼》[36]中详细记录了叫姑鱼属受精卵和胚体环绕卵黄1/2时期鱼卵内部的形态变化特征,本文研究的叫姑鱼属卵径和油球径均在其记录的合理范围内。科早期生活史的形态学基础资料相较于其他鱼类鱼卵是最为完整的,万瑞景和张仁斋[5]通过人工授精和孵化详细记录了多鳞鱼卵11个发育时期的形态变化特征。相近种类鱼卵的结构特征具有一定的相似性[34],单纯依靠形态学分类难以将相近种类鉴定到种。鲾科鱼类个体较小、数量较大,在珠江口海域一直处于优势地位[37],然而其早期生长阶段的形态变化却未被完全掌握,沖山宗雄[38]以图简单示出颈斑项鲾 (Nuchequula nuchalis)鱼卵的形态,本文所获取鲾科鱼卵的形态特征可对现有资料进行有效补充。早期形态特征研究多关注具有重要经济价值的种类,如日本鳗鲡 (Anguilla japonica)[39]和马口鱼 (Opsariichthys bidens)[40]等,许多个体小、经济价值低但数量较大且具有重要生态作用的种类未被重视[15],因而珠江口江门海域常见小型鱼类早期发育阶段的形态鉴定资料较少。本文通过对种类鉴定结果与鲜活样品的形态记录进行分析,对江门海域13种鱼卵的形态特征进行了描述,可为今后江门海域鱼卵形态的研究提供基础资料。
3.3 不足及展望
本文依据从采获样品中获取的鱼卵形态特征,结合历史资料,尝试依据鱼卵形状和大小、卵周隙宽窄、油球数量和色素分布等特征进行总结,可大致将研究海域春季鱼卵种类进行初步分类,但研究仍存在许多不足。首先,鱼卵不同发育阶段的形态变化较大,本文仅可获取鱼卵单一发育阶段的形态信息,未掌握其他发育阶段的形态特征。种属间相近种类鱼卵内部形态特征相似,难以根据单一阶段形态进行区分,还需结合人工培养等获取鱼卵更多发育阶段可靠的形态学信息。珠江口江门海域作为鱼类重要的产卵场,本文通过单次采样获得的种类相对有限,构建完整的形态分类检索还需依托更多的采样调查。其次,本文采集的是鲜活鱼卵样品,种类丰富、个体数量多,短期内获得了多个种类的形态特征信息,但是鱼卵样品处理及形态记录需要一定时间,所处水体环境的骤然改变,导致某些种类鱼卵的保存状态变差。部分样品基因组降解导致种类鉴定失败;部分样品受到水霉菌等的污染而死亡,导致鱼卵内部模糊不清,也在某种程度上改变了样品的外在形态,影响了鱼卵种类鉴定和形态特征获取的效果。为尽可能获取更多发育阶段鱼卵的形态特征,野外采获后的鱼卵需保持水体洁净,并在减少人为干扰的情况下尽快观察和记录其形态。公共数据库中序列的丰富性、准确性和完整性可极大地影响DNA条形码的鉴定结果[17,29],此外,沙丁鱼属等种类属于单倍型群体,单一的COI片段不能满足其鉴定至种的要求,因而加快建立并完善相关海域多种类、多片段的成鱼基因数据库是应用DNA条形码准确鉴定鱼卵的必要前提条件。同时在进行鱼卵资源调查时,结合同期采集的成鱼样品进行种类分析或可提高鱼卵鉴定及其形态特征获取的准确性[8,41]。
4 结论
本文根据2021年春季珠江口江门海域的浮游生物调查,阐述了江门海域13种常见鱼卵的形态学特征。研究发现大部分鱼卵可通过卵膜形状,卵径大小,油球数量、分布及其色素分布,卵周隙宽窄,胚体上色素胞颜色、数量及分布等形态特征进行分类。然而相近种类难以通过单一发育阶段的形态特征进行准确区分,文中借助DNA条形码技术对形态学分类结果进行验证,发现两者的鉴定结果在科水平上较为一致,且DNA条形码技术能成功对江门海域鱼卵进行更低阶元的划分。受限于公共数据库中鱼类数据的完整性和准确性,借助DNA条形码进行种类分析时,为谨慎起见,相关鱼卵种类未能鉴定至种。本文首次针对珠江口江门海域春季鱼卵种类进行形态特征研究,未涉及其他季节,鱼卵种类相较于同期捕获的成鱼种类较少,今后为更全面地掌握珠江口江门海域鱼卵的形态特征和检索资料,还需要进行重复采样和深入研究。
猜你喜欢
天津农学院学报(2024年1期)2024-04-07 11:20:22
水产学报(2024年3期)2024-03-25 07:43:22
南方医科大学学报(2022年6期)2022-06-30 08:36:06
食品工业(2020年6期)2020-07-18 04:07:20
创新作文(3-4年级)(2019年8期)2019-12-17 07:59:21
天工(2019年7期)2019-01-20 10:02:10
创新作文(小学版)(2019年23期)2019-01-11 10:37:45
浙江农业学报(2016年3期)2016-10-31 09:20:59
动物营养学报(2015年10期)2015-12-01 02:26:34
畜牧兽医学报(2015年8期)2015-03-22 07:36:32
