
宿半夏凝集素基因的克隆与分析
2022-12-19 09:10朱艳芳梁志鹏
淮北师范大学学报(自然科学版) 2022年4期
朱艳芳,朱 斌,梁志鹏,董 庆,薛 涛,盛 玮
(1.淮北师范大学 生命科学学院,安徽 淮北 235000;2.安徽省特色资源植物利用工程实验室,安徽 淮北 235000)
0 引言
半夏Pinellia ternata(Thunb.)Breit为天南星科半夏属多年生草本植物,其块茎为主要的药用部位,是一种常用的中草药. 半夏在中药处方中的使用率位居前列,有着很高的药用价值[1]. 半夏始载于《神农本草经》,中国药典记载其功效为燥湿化痰、降逆止呕、消痞散结等,可用于治疗湿痰寒痰、咳喘痰多、痰饮眩悸、风痰眩晕、痰厥头痛、呕吐反胃、胸脘痞闷、梅核气等,外治痈肿痰核. 现代临床应用中,证明半夏具有治疗恶性肿瘤的重要作用[2]. 目前,普遍认为半夏提取物以及半夏化学成分中的半夏蛋白、半夏总生物碱、谷甾醇和半夏多糖等都具有抗肿瘤作用[3].
半夏蛋白是半夏的主要药效成分之一,主要由半夏胰蛋白酶抑制剂和半夏凝集素组成[4]. 半夏凝集素属于单子叶甘露糖凝集素,能够识别并特异性的结合甘露糖,具有抑菌、杀虫、凝血和抗肿瘤等多种生理功能[5-10]. 由于半夏凝集素的生理功能多样,对半夏凝集素的研究一直是个热点,其研究多集中于对该蛋白的提取纯化[4,11]、功能研究[3-4,12]、基因克隆[13]、异源转化[8,14]等方面. 宿半夏是分布于皖北的道地药材,尚未见针对宿半夏凝集素的相关报道. 因此,本研究以宿半夏的块茎为试验材料,对其凝集素基因进行克隆和生物信息学分析,为进一步研究宿半夏凝集素及深入开发宿半夏资源利用奠定基础.
1 材料与方法
1.1 试验材料
本试验所用半夏为安徽道地药材宿半夏P.ternata的新鲜块茎. 宿半夏块茎盆栽于淮北师范大学生命科学学院生物化学与分子生物学实验室,以盆栽宿半夏叶柄为外植体,经消毒后接种于MS基本培养基,获得宿半夏无菌苗,培养温度为(25±2)℃,光暗周期为12 h/12 h,光照强度为2 000 lx.
1.2 试验方法
1.2.1 总RNA的提取、检测及反转录
以无菌组培的半夏新鲜块茎为材料,上海生工Total RNA Extractor(Trizol)试剂提取新鲜块茎总RNA,利用微量分光光度计检测总RNA 的质量和浓度. 高质量的RNA 用莫纳生物反转录试剂盒合成cDNA第一链,-70 ℃保存备用.
1.2.2 引物设计与合成
根据半夏块茎发育转录组测序数据结合Genbank数据库登录半夏属凝集素基因序列,设计半夏凝集素基因(PtPTA)上下游引物,用于半夏凝集素基因的克隆,引物序列为:
PTA-F:5’-ATGGCCTCCAAGCTCCTCCTC-3’
PTA-R:5’-TTAATTCACCTTCTCCGTCACCATG-3’
1.2.3PtPTA克隆、胶回收及测序
以cDNA为模板,PTA-F、PTA-R为引物进行PCR扩增反应. PCR产物经质量分数为1%琼脂糖凝胶电泳,回收纯化目标基因,并与克隆载体pMD18-T连接,转化至大肠杆菌DH5α感受态细胞,挑取单菌落,37 ℃在含Amp的LB液体培养基中震荡培养. 以载体通用引物做菌落PCR检测,将阳性转化子送通用生物系统(安徽)有限公司测序.
1.2.4 生物信息学分析
将测序获得的PtPTA序列进行在线分析和软件分析:用NCBI ORF Finder(http://www.ncbi.nlm.nih.gov/orffinder)在线查找序列的最大开放阅读框;用在线分析工具ProtParam(http://www.expasy.ch/tools/protparam/html)分析蛋白质的基本性质;在NCBI 网站的Conserved Domain Database 数据库(http://www.ncbi.nlm.nih.gov/cdd)分析蛋白质保守结构域;用DNAMAN 软件将天南星科不同来源的数种凝集素蛋白质序列进行多序列比对(参数均为默认值);用MEGA7.0 构建系统进化树(Bootstrap 为1 000);用WoLF PSORT(http://wolfpsort.hgc.jp)对该蛋白进行亚细胞定位预测;用TMHMM Serer V2.0对蛋白进行跨膜结构预测;用SOMPA(https://npsa-prabi.ibcp.fr/cgi-bin/secpred_sopma.pl)在线预测蛋白质的二级结构;用SWISS-MODEL(https://www.expasy.org/resources/swiss-model)在线预测蛋白质的3D结构.
2 结果与分析
2.1 半夏块茎总RNA质量检测
电泳检测提取半夏块茎RNA 质量,结果如图1 所示. 分别得到3 条清晰的RNA 条带,即28S、18S 和5S RNA条带,其中28S RNA 条带的亮度是18S RNA条带亮度的2倍,说明提取的总RNA完整性好,几乎无降解. 超微量分光光度计检测样品RNA的A260/A280为1.913,A260/A230为0.998,结果显示样品的RNA纯度高,污染少,可用于后续的基因克隆实验.
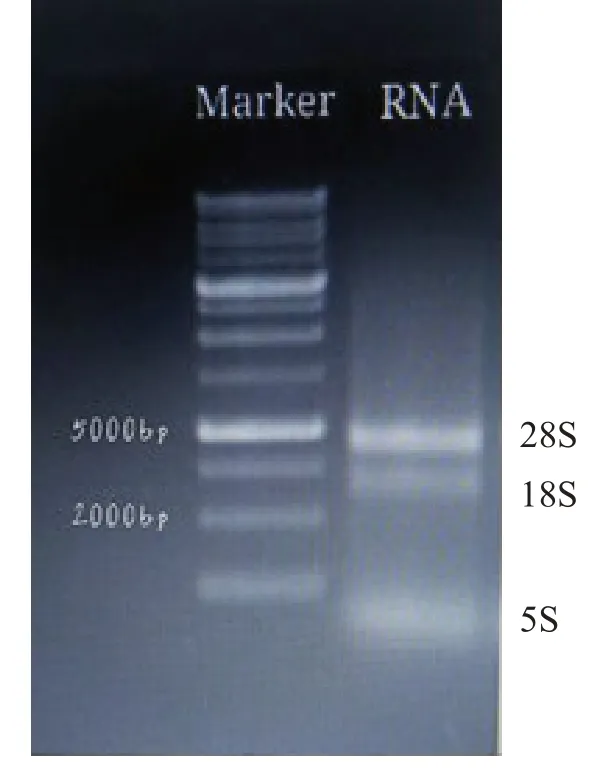
图1 RNA凝胶电泳图
2.2 半夏凝集素基因(PtPTA)的克隆及测序
以cDNA为模板,以PTA-F,PTA-R为引物,PCR 扩增PtPTA. 将PCR产物凝胶电泳,其电泳结果如图2所示. 与Marker相比,在750 bp和1 000 bp之间有一条明亮的带,得到约800 bp特异片段.
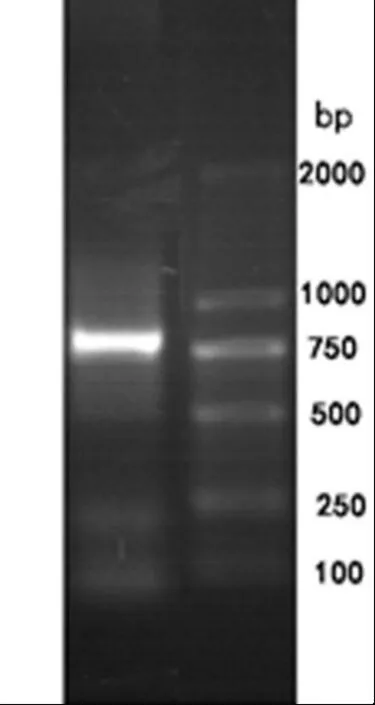
图2 PCR产物电泳结果图
对PCR扩增所得条带胶回收并测序,获得一条开放阅读框全长为810 bp的序列,编码269个氨基酸(图3). 将目的基因编码的蛋白质与NCBI中登录的其它6种半夏植物凝集素蛋白进行序列比对,结果显示,几种半夏植物凝集素的同源性比较高,一致性达到97.82%(图4).

图3 PtPTA全长cDNA序列及其推导氨基酸序列
2.3 PtPTA编码蛋白(PTA)特性分析
2.3.1 PTA理化性质
理化性质分析结果显示,PTA 蛋白的分子式为C1305H2028N354O391S11,由269 个氨基酸残基组成,总分子量为29 285.25,理论等电点(pI)为6.58;带负电氨基酸残基(Asp+Glu)总数为23,带正电氨基酸残基(Arg+Lys)总数为22. 不稳定系数为25.30,脂肪系数为87.70,亲水性系数为-0.161,属于稳定的亲水性蛋白.
2.3.2 PTA的跨膜区域预测
用TMHMM预测PTA的N-端第5~27 个氨基酸残基位于跨膜区域(图5),说明PTA属于跨膜蛋白,推测PTA 可能定位于细胞膜上,属于分泌蛋白.用WoLF PSORT 预测该蛋白亚细胞定位于细胞外,与TMHMM预测相一致.SignalP预测该蛋白质有信号肽的概率为99.700%,信号肽类型为SP(Sec/SPI),切割位点位于第24位丙氨酸A和第25为缬氨酸V残基之间,信号肽序列为:MASKLLLFLLPAILGLVIPPAATA.
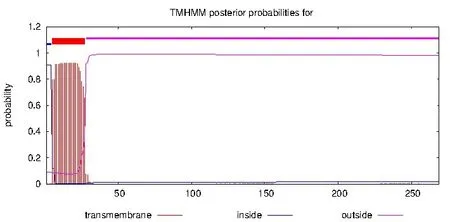
图5 PTA跨膜区结构分析
2.3.3 PTA疏水性分析
通过Protscale 对PTA 的疏水性进行预测表明,多肽链第6 位的亮氨酸(Leu,L)具有最高分值2.98,第182位的赖氨酸(Lys,K)具有最低的分值-2.19. 已知氨基酸分值越低亲水性越强,反之分值越高疏水性越强,可以得出在第182位的Lys亲水性最强,第6位的Leu疏水性最强. 从图6中可以看出,该蛋白质N-端部分氨基酸残基和中间区域的部分氨基酸残基是由一些疏水性氨基酸组成,其余区域主要由亲水性氨基酸组成. 从整体看,其疏水性氨基酸少于亲水性氨基酸,所以推测其为亲水性蛋白.
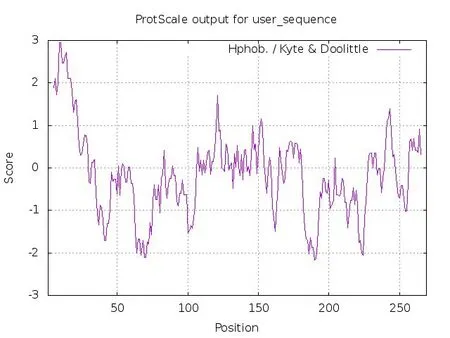
图6 PTA疏水性分析
2.3.4 PTA结构域分析
NCBI在线分析PTA结构域,如图7所示,该蛋白质包括2个B-lectin结构域,属于B-lectin超家族,且包含3个甘露糖结合位点,见图4中黑色下划线标示序列. 由图4可知,3个甘露糖结合位点的保守性不同,第1和第2个甘露糖结合位点保守性高,而第3个甘露糖结合位点氨基酸组成差异大,保守性低,说明不同半夏植株的凝集素仍存在一定差异.

图7 PTA结构域
2.3.5 PTA二级结构及三维结构预测
用SOPMA在线预测PTA二级结构见图8,PTA二级结构主要是无规则卷曲,占据45.72%的氨基酸残基;其次是β-折叠,占据33.46%的氨基酸残基;α-螺旋占据11.15%的氨基酸残基;最少的是β-转角,占据9.67%的氨基酸残基. α-螺旋主要位于蛋白质的两端,β-折叠、β-转角和无规则卷曲散布在蛋白质中间.

图8 PTA二级结构预测
通过SWISS-MODEL 在线比对,发现PTA 有2 个结构域,分别比对到氨基酸序列相似度都在80%以上的高度相似的模板,模板分别是Structure of Lectin fromColocasia esculenta(L.)Schott(SMTL ID:5j76.1)和Structure ofColocasia esculentaagglutinin(SMTL ID:5d5g.2),分别是芋的贮藏蛋白植物凝集素和芋块茎凝集素. 以2个已知蛋白模型为基础,预测PTA两个结构域的3D结构见图9,A是1~109个氨基酸形成结构域的3D结构,B是117~227个氨基酸形成结构域的3D结构.

图9 PTA三维结构预测
2.3.6 PTA系统进化
从NCBI 数据库中选取12 条凝集素蛋白与PTA 进行相似性比较,构建系统进化树(图10). 结果表明:本次克隆的宿半夏凝集素基因编码的蛋白PTA 与其他6 种半夏凝集素AAR27794.1、ABX47148.1、AAU29612.1、AFY06641.1、QNL35375.1和AAP20876.1遗传关系近,聚为一支,与海芋凝集素ABC69036.1的关系最远. 说明半夏凝集素的进化关系与物种进化关系基本一致. 半夏凝集素与掌叶半夏凝集素AAR27793.1聚为一大支,与石蜘蛛凝集素AOA49622.1未聚在一起,还有一种半夏凝集素AEZ35184.1未与多种半夏凝集素聚合在一起.

图10 利用NJ法构建植物凝集素间的进化树
3 讨论和结论
植物凝集素是一类分布广泛的非免疫来源的糖结合蛋白,由于其具有显著的生物活性而得到广泛的研究和应用. 植物凝集素可以通过识别并结合动物、昆虫肠胃及微生物表面特异性糖基,从而发挥促进有丝分裂、抗氧化、抗肿瘤、降血糖、抗菌、杀虫、抗病毒等活性[15];也可以提高植物对环境的适应能力,如耐旱、耐盐、耐热性[16];植物凝集素广泛存在于植物的各种组织器官中,在根、茎、叶、花、果实中都能被检测到,以种子和营养贮存器官(如块茎、鳞茎、块根)中最为丰富,参与植物的生长发育过程[17].
半夏凝集素(Pinellia ternataagglutinin,PTA)是一种典型的具有甘露糖结合位点的植物凝集素[4],属于雪花莲家族(GNA)的凝集素[18]. 半夏蛋白具有很多生理功能,其中半夏凝集素是半夏蛋白发挥其生理功能的重要成分之一. 尽管对不同产地的半夏凝集素开展一定的研究[4,13,18-19],但是由于半夏的高度遗传多样性,不同半夏之间遗传物质有一定的差异[20]. 宿半夏的凝集素至今未见报道,本研究克隆宿半夏的凝集素基因,为宿半夏凝集素的应用及优良品种选育提供参考.
本试验成功克隆半夏块茎中的凝集素基因,其开放阅读框长度为810 bp,编码269个氨基酸,分子量为29 285.25,等电点为6.58,具有典型的跨膜结构域,属于跨膜亲水蛋白,预测定位于细胞膜,与赵欢等[18]研究结果PTA亚细胞定位于细胞膜上相一致. 多序列比对结果显示该蛋白与其他6种半夏植物凝集素氨基酸序列具有高度的相似性,相似度达97.82%,说明不同半夏植株的凝集素序列具有高度的保守性,该结果与赵欢[18]的研究结果一致. 不同产地、不同生境下,不同半夏植株的半夏凝集素也存在一定差异,尤其第3个甘露糖结合位点存在较大差别,该结果与张正英[13]研究结果一致,同样也说明半夏的遗传多样性.
通过比对,本次克隆的宿半夏PTA与赵欢等[18]克隆的半夏凝集素蛋白序列(AGV40777.1)完全一致,赵欢所用材料是川半夏的主产区四川省南充市的野生半夏叶片DNA,本研究用的材料是安徽省淮北市道地药材宿半夏块茎RNA,两半夏材料生境差别大,川半夏基因组的半夏凝集素和宿半夏块茎中表达的半夏凝集素的氨基酸序列完全相同,那么2种半夏块茎中凝集素的含量是否相同,其他药效成分含量是否相近,2种半夏能否互相替代,还有待于进一步深入研究. 系统进化树分析表明,大部分半夏凝集素聚合在一起,唯有半夏凝集素AEZ35184.1游离在外,分析发现半夏凝集素AEZ35184.1是由257个氨基酸组成,与半夏PTA 的一致性为91.88%. 而赵欢等[21]认为半夏凝集素PTA 的氨基酸序列长度为268-269aa,半夏凝集素AEZ35184.1还有待于进一步分析.
猜你喜欢
生物化学与生物物理进展(2022年6期)2022-07-21
今日农业(2021年15期)2021-11-26
中学生数理化(高中版.高考理化)(2021年2期)2021-03-19
现代园艺(2017年19期)2018-01-19
中华老年多器官疾病杂志(2016年7期)2016-04-28
池州学院学报(2015年3期)2016-01-05
天津科技大学学报(2015年2期)2015-08-09
云南中医学院学报(2015年2期)2015-07-31
医学研究杂志(2015年5期)2015-06-10
国际心血管病杂志(2015年5期)2015-02-27
