
澳门地区褐家鼠对溴鼠灵的抗性检测及其Vkorc1基因多态性分析
2022-12-19 12:48:14郭敏梁捷何向阳欧伟新彭定雄麦展昭黄海涛张礼标
兽类学报 2022年6期
郭敏梁捷何向阳欧伟新彭定雄麦展昭黄海涛,3*张礼标*
(1广东省科学院动物研究所,广东省动物保护与资源利用重点实验室,广东省野生动物保护与利用公共实验室,广州 510260)(2澳门市政署,澳门特别行政区 999078)(3广东科建生物技术有限公司,广州 510260)
预防鼠患是保障农业及城市健康发展的一项重要工作(石杲,2003;孟庆云,2003)。在鼠患预防中,抗凝血灭鼠剂在全球使用量最大,共历经了两代(陈心尧,1992;刘刚和曾虹,2002),溴鼠灵(brodifacoum)属于第二代抗凝血灭鼠剂。溴鼠灵于1975年开始投入使用,药效比第一代抗凝血灭鼠剂强约100倍,能迅速杀死对一代药和部分二代药(如溴敌隆和鼠得克)有抗性的褐家鼠(Rattus norvegicus)(Buckleet al.,2012),其毒性不仅限于凝血功能障碍(Bachmann and Sullivan,1983;Wareet al.,2015;Kalininet al.,2017),也包括细胞损伤(Bakeret al.,2002;Gulet al.,2016)。由于溴鼠灵药效强、田间试验效果好,该药物已在国内外推广使用了数十年。目前对溴鼠灵的研究主要集中在灭杀高效性(宋明亮等,2002;熊孟韬等,2003;Buckleet al.,2012;陈蔚恩等,2016)和生物安全性(Easonet al.,2002;Ayalaet al.,2007;Geduhnet al.,2014;Alomaret al.,2018;Waltheret al.,2021)等方面,但对其长期使用是否造成鼠类抗药性的进化,尤其是维生素K环氧化物还原酶复合体亚单位1(vitamin K-epoxide reductas complex 1,Vkorc1)基因核苷酸改变的相关研究还比较缺乏。
澳门地区已使用溴鼠灵近25年,为调查鼠类对溴鼠灵抗药性进化提供了良好模型。本研究拟结合生理抗性检测和Vkorc1基因突变检测来探查澳门地区褐家鼠对溴鼠灵的抗性及进化,试图通过该研究为鼠类对溴鼠灵抗药性的发生和发展提供理论依据,引导溴鼠灵的安全使用,更好地预防鼠患。生理抗性检测参考全国鼠类抗药性监测协作组的致死期食毒法(LFP)开展(鼠类抗药性监测协作组,1991;张涛和吴明寿,2007)。例如陈戊申等(2007)使用LFP法进行溴鼠灵的灭杀效果评价,发现毒饵消耗的高峰期在第3天,总消耗量超过12 mg/kg(毒饵/体重),但死亡高峰期在第6天。在有统计学意义的个体数量中,LFP法检测到的抗性鼠若占比超过15%,则认为该地区该种鼠群对所用灭鼠药已产生抗性(鼠类抗药性监测协作组,1991)。此外,结合Vkorc1基因突变检测,可对鼠类抗药机理及抗性关联进化进行深入挖掘。例如对黄胸鼠(Rattus flavipectus)和褐家鼠进行生理抗性测试实验(华法林)和Vkorc1基因突变检测,发现了抗性鼠以及与抗性有显著关联的Vkorc1基因突变位点(褚敏捷等,2018)。姚丹丹等(2019)亦通过生理抗性测试实验(杀鼠灵)和Vkorc1基因突变检测,发现了黄毛鼠(Rattus losea)抗性种群及12个Vkorc1基因突变位点,但这些突变位点与生理抗性测试结果无显著相关性。前人研究揭示,褐家鼠Vkorc1基因的外显子中至少有8个位点与抗药有关联,其中Y(Tyr)139C(Cys)的突变与两代灭鼠药的抗性关联最强(Rostet al.,2009;Buckleet al.,2012;Mooneyet al.,2018)。而且随着鼠种的不同,Vkorc1基因发生抗性突变的位点和方式也不同(Rostet al.,2009;王智泉等,2014;褚敏捷等,2018)。
1 研究方法
1.1 研究对象
研究对象为澳门地区捕捉到的61只褐家鼠,捕捉方法为笼捕法,对捕捉到的全部褐家鼠个体进行称重、性别鉴别以及编号。选择体重≥80 g的44只健康成年鼠(34只捕获于澳门红街市,10只捕获于澳门营地街市)进行生理抗性检测。剔除实验前死亡、不满足实验条件和实验期间产仔的个体,最后完成生理抗性检测的褐家鼠个体为35只,其中雌鼠11只,雄鼠24只。此外,对全部61只褐家鼠进行Vkorc1基因序列多态性检测。
1.2 褐家鼠生理抗性检测
于2019年12月2日至14日对捕获于澳门红街市(34只)和营地(10只)的褐家鼠,参照LFP法进行生理抗性检测实验。
具体步骤依次为:(1)适应期喂养。将实验鼠单笼分装,开展4 d的适应期喂养,即投喂无毒的番薯块和足量的纯净水。(2)毒饵制备及食毒期喂养。将澳门地区当前主要投放的第二代灭鼠药TALON(厂家:Syngenta;有效成分:brodifacoum)与番薯块按0.005%(质量比:w/w)混合拌匀来制备毒饵。每天供应充足的毒饵(≥40 g)和足量的纯净水,持续6 d,记录实验鼠个体当日毒饵消耗量(测算方法见下)。淘汰食毒期第一天毒饵消耗量不及前一天无毒饵料消耗量1/10的个体。再根据每日实验鼠食毒量和体重比值的均值,淘汰当天毒饵消耗量异常的个体。若食毒期个体死亡,记录死亡鼠的出血部位和死亡时间;对无出血表征的死亡鼠进行解剖,观察有无内出血。(3)观察期喂养。将食毒期结束后的余存个体转移至干净鼠笼进行无毒喂养观察,即喂食无毒番薯块和足量的纯净水,持续14 d。若观察期出现个体死亡,记录死亡鼠的出血部位和死亡时间;对无出血表征的死亡鼠进行解剖,观察有无内出血。此外,淘汰实验期间产仔的个体。
毒饵消耗量测算方法为:投放毒饵时在笼下铺放干净的纸,次日收集纸上的饵料(剔除粪便),与笼内剩饵一起称量,来计算个体每日毒饵消耗量。
1.3 褐家鼠Vkorc1基因的突变检测
2019年10—12月在澳门地区捕捉到61只褐家鼠(包括已完成生理抗性检测实验的44只褐家鼠),取其肌肉和肝脏组织,分别保存于无水酒精和RNAlocker(厂家:生工)中。然后,利用Ezup柱式动物基因组DNA抽提试剂盒(厂家:生工)提取样品的基因组DNA,再扩增Vkorc1基因片段,最后通过对扩增片段测序分析来检测Vkorc1基因序列的核苷酸多态性。
由于鼠类Vkorc1基因全长约为2 800 bp,难以通过一对引物来扩增出全长片段。因此,参照Goulois等(2016)用两对引物(两对引物的扩增区间能覆盖Vkorc1基因全部的3个外显子区)来分段扩增Vkorc1基因序列,以检测Vkorc1基因外显子区的核苷酸多态性。第一对引物目标扩增片段为Vkorc1基因的前1 105 bp(5′-3′方向):正向引物序列 为rVKOR-S1(5′-GGTTCTTCCCTCTTGTGTCTG-3′),反向引物序列为rVKOR-AS1(5′-GGGTCACCAAGACATGAGGTG-3′);第二对引物目标扩增片段为Vkorc1基因的后1 335 bp:正向引物序列为rVKORC1-S2(5′-ACTT-GGGCAAGGCTCATGTG-3′),反向引物序列为rVKORC1-AS2(5′-AAGAGTAGGGGACAAGGTG-GC-3′)。
PCR扩增条件:94℃3 min;94℃20 s,52℃20 s,68℃2 min(12个循环);94℃20 s,59℃20 s,68℃2 min(38个循环);68℃10 min;10℃30 min。PCR扩增产物电泳检测的凝胶浓度为1.5%,上样量为2 μL。最后使用ABI 3730XL测序仪对阳性PCR扩增产物进行双向测序。
1.4 数据分析
通过计算实验鼠个体的摄毒量与体重的比值,得到个体的单位体重摄毒量。然后分别按地点(红街市、营地)和性别(雌、雄)对实验鼠进行分组,对个体分组后的单位体重摄毒量数据,进行以下分析:(1)计算各组单位体重摄毒量,用平均值±标准误(mean±SE)来表示;(2)分析组间单位体重摄毒量差异的显著性。先通过R软件的“ks.test”函数(One-sample Kolmogorov-Smirnov test)检测各组数据是否呈正态分布,若P>0.05则是正态分布;再通过R软件的“var.test”函数(F-test)检测分组数据是否方差齐性,若P>0.05则方差齐性。对符合正态分布且方差齐性的分组数据,通过R软件的“t.test”函数进行t-test分析,检测组间差异的显著性;对不符合正态分布且方差齐性的分组数据,则通过R软件的“wilcox.test”函数分析来检测组间差异的显著性。
以NCBI数据库中挪威褐家鼠的Vkorc1基因(https://www.ncbi.nlm.nih.gov/gene/309004)为参考基因,进行澳门褐家鼠Vkorc1基因外显子核苷酸多态性分析。分析方法:通过SeqMan Pro将扩增得到的全部澳门褐家鼠Vkorc1基因序列与参考基因序列进行比对,比对后有差异的核苷酸Seq-Man Pro会自动以红色标示出来。再以参考基因3个外显子CDS的起止位置(https://www.ncbi.nlm.nih.gov/nuccore/NC_051336.1?report=genbank&from=182502491&to=182505012&strand=true)为基准,界定澳门褐家鼠Vkorc1基因的外显子区段。最后人工统计澳门褐家鼠Vkorc1基因与参考基因在外显子区段有差异的核苷酸的位置、种类和频次。
2 结果
2.1 褐家鼠生理抗性
生理抗性检测的35只实验鼠均在实验期死亡。31只在食毒期死亡,剩余4只在观察期首日死亡。雄鼠的平均致死时长为3.3 d,雌鼠为3.5 d。实验前,雄鼠个体体重均值为248 g(86~406 g),雌鼠个体为258 g(122~470 g),雄鼠和雌鼠的平均摄毒量均高于12 mg/kg:雄鼠摄毒量2.02~35.03 mg/kg,平均为(15.69±1.59)mg/kg,雌鼠摄毒量0.17~31.43 mg/kg,平均为(14.39±2.93)mg/kg,总体平均摄毒量为(15.28±1.40)mg/kg。
此外,分别按地点和性别对实验鼠分组后,组内数据均不符合正态分布(One-sample Kolmogorov-Smirnov test,P<0.05),故采用Wilcoxon方法检验组间差异显著性,结果显示组间溴鼠灵的平均摄毒量均无显著差异(地点间:P=0.254 7;性别间:P=0.687 3)(图1)。
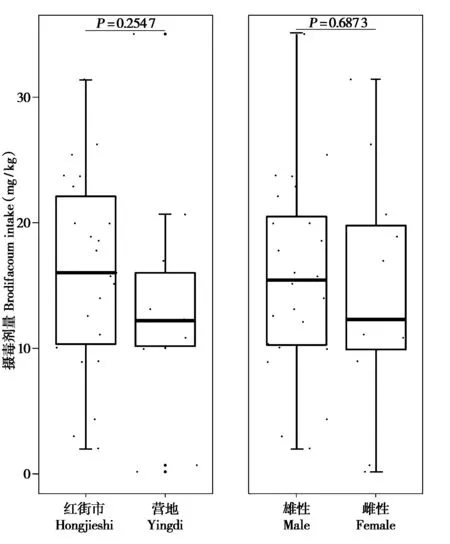
图1 澳门地区不同地点、性别褐家鼠的摄毒量.图中圆点代表实验鼠个体.纵坐标表示个体在食毒期内的单位体重摄毒剂量(mg/kg).通过Wilcoxon方法检验按地点或性别分组的褐家鼠组间平均摄毒量差异的显著性Fig.1 Variations in brodifacoum intake among R.norvegicus grouped by gender and living site from Macao.Solid dots denote rat individuals.The Y-axis shows individual brodifacoum intake per weight(mg/kg)after LFP test.Significance of the difference between genders or living sites was detected by Wilcoxon method
2.2 褐家鼠Vkorc1基因外显子核苷酸多态性
2.2.1 褐家鼠Vkorc1基因外显子序列扩增
用两对引物对全部61只褐家鼠Vkorc1基因的外显子区段进行扩增,结果显示,第一对引物(rVKOR-S1、rVKOR-AS1,扩 增Vkorc1基 因 前1 105 bp序列)的扩增效果明显比第二对引物(rVKORC1-S2、rVKORC1-AS2,扩增Vkorc1基因后1 335 bp序列)好:第一对引物成功扩增出57只(93%)褐家鼠Vkorc1基因的前1 105 bp序列,其中55只在正向和反向引物上均测序成功;第二对引物仅成功扩增出20只(33%)褐家鼠Vkorc1基因的后1 335 bp序列,其中13只在正向和反向引物上均测序成功。最终,58只(95%)褐家鼠的Vkorc1基因被扩增到阳性条带,但仅12只两对引物均能成功扩增和测序。3只褐家鼠的Vkorc1基因两对引物均无扩增条带(表1)。

表1 Vkorc1基因片段阳性扩增子测序成功的褐家鼠个体数Table 1 Number of R.norvegicus successfully sequenced after positive amplification of Vkorc1 gene fragments
2.2.2Vkorc1基因外显子核苷酸多态性
鼠类Vkorc1基因有3个外显子,共表达161个氨基酸(aa)。本研究中,褐家鼠Vkorc1基因的3个外显子区均能被成功扩增序列所覆盖,但由于两对引物的扩增成功率有较大差异,从而表现出有的外显子区段覆盖次数多,有的则少。通过与NCBI数据库中挪威褐家鼠Vkorc1基因的外显子编码序列(CDS)比对,澳门褐家鼠Vkorc1基因的外显子编码序列仅2个核苷酸位点有差异,且均为同义突变,分别为:第68位氨基酸H(CAC)-H(CAT)(共有55只褐家鼠扩增到该核苷酸位点,突变发生率为100%)和第82位氨基酸I(ATA)-I(ATT)(共有55只褐家鼠扩增到该核苷酸位点,突变发生率为32.72%)。本研究发现,澳门地区褐家鼠个体的Vkorc1外显子序列有很强的保守性,通过进一步整理,得到澳门褐家鼠Vkorc1基因3个外显子编码区(CDS)的保守(一致)核苷酸序列(161 aa):第 一 外 显 子 为ATGGGCACCAC CTGGAGGAGCCCTGGACGTTTGCGGCTTGCAC TATGCCTCGCTGGCCTAGCCCTCTCACTGTACG CACTGCACGTGAAGGCGGCGCGCGCCCGCAA TGAGGATTACCGCGCGCTCTGCGACGTGGGCA CGGCCATCAGCTGTTCCCGCGTCTTCTCCTCTC G;第二外显子为GTGGGGCCGGGGCTTTGGG CTGGTGGAGCATGTGTTAGGAGCTGACAGCAT CCTCAACCAATCCAACAGCATTTTTGGTTGCAT GTTCTACACCATACAGCTGTTGTTAG;第 三 外显子为GTTGCTTGAGGGGACGTTGGGCCTCT ATCCTACTGATCCTGAGTTCCCTGGTGTCTGTC GCTGGTTCTCTGTACCTGGCCTGGATCCTGTTC TTTGTCCTGTATGATTTCTGCATTGTTTGCATCA CCACCTATGCCATCAATGCGGGCCTGATGTTGC TTAGCTTCCAGAAGGTGCCAGAACACAAGGTC AAAAAGCCCTGA。
3 讨论
澳门是世界上人口密度最高的地区之一,1995年后开始使用第二代抗凝血灭鼠剂。基于澳门独特的城市及发展背景,对该地区鼠类抗药能力的调查能为鼠类对第二代抗凝血灭鼠剂抗药性的发生和发展提供非常有价值的参考。本研究以澳门地区褐家鼠对溴鼠灵的抗药性为切入点,通过生理抗性检测和Vkorc1基因分子检测相结合的方法,探索鼠群对溴鼠灵的抗药发展。结果表明,澳门地区投放溴鼠灵近25年,没有出现褐家鼠群体抗药性,揭示该地区可继续使用溴鼠灵灭鼠剂,但检测到2个同义核苷酸突变,表明仍需要持续监测抗药性的发生和发展。
本研究使用的抗凝药溴鼠灵问世于1975年,药效比第一代抗凝血灭鼠剂强,鼠类食用后主要富集于肝脏,并能维持高浓度超过96 h。溴鼠灵对人类及与人类生活关联密切的动物也有毒性(Breckenridgeet al.,1985;Booth,1989;Boermanset al.,1991;Easonet al.,2002;Ayalaet al.,2007;Geduhnet al.,2014;Booth and Mody,2016;Fitzgeraldet al.,2018),而且还可以通过食物链进行传播和累积(Easonet al.,2002;Ayalaet al.,2007;Geduhnet al.,2014;Alomaret al.,2018;Waltheret al.,2021)。这强调了全球化背景下对各地区溴鼠灵的使用监管以及调查鼠类对溴鼠灵抗药的必要性。
尽管溴鼠灵已在澳门地区投放近25年,本研究尚未发现抗溴鼠灵的褐家鼠,可能有以下3个主要原因。第一,溴鼠灵的强毒性。溴鼠灵对鼠类机体功能的破坏高效且致命(Buckleet al.,2012),以致于溴鼠灵自问世以来历经了约45年的使用,国内外仍没有发现溴鼠灵抗性鼠的报道。第二,溴鼠灵的致病复杂性。除了凝血功能障碍,溴鼠灵还能造成鼠类多种生理机能损伤(Bachmann and Sullivan,1983;Bakeret al.,2002;Wareet al.,2015;Gulet al.,2016;Kalininet al.,2017),关联基因多,鼠类很难通过短时间的进化来产生全面抗性。第三,溴鼠灵并非澳门地区唯一使用的灭鼠药,市区内其他灭鼠药的并行使用减缓了溴鼠灵对鼠群的选择压力。这点从Vkorc1基因的突变情况能有所反映:相比挪威褐家鼠,澳门褐家鼠的Vkorc1基因仅检测到2个同义突变,且均发生在氨基酸密码子的第三位,该位点本身就具有很强的摆动性。这两个同义突变是否蕴含其他的进化生物学意义还有待挖掘。
由于溴鼠灵的致病复杂性以及无可参考的溴鼠灵抗性鼠Vkorc1基因,已发现的对第一代和部分二代(溴敌隆和鼠得克)抗凝血灭鼠剂有抗性的Vkorc1基因突变位点对溴鼠灵未必有效,反映出开展生理抗性检测的重要性。本研究中,澳门地区褐家鼠虽然在食毒期及观察期内全部死亡,显示出溴鼠灵灭杀的高效性,但总体平均摄毒量高达(15.28±1.40)mg/kg(高于12 mg/kg)。出现这种情况可能的原因包括:其一,溴鼠灵良好的适口性以及慢性发毒过程。国内研究人员对褐家鼠和大白鼠进行0.005%溴鼠灵毒饵投喂实验发现,尽管实验鼠在4~7 d内全部死亡,但均有很高(摄毒量均高于12 mg/kg)的毒饵消耗量(宋明亮等,2002;熊孟韬等,2003;陈蔚恩等,2016),这也表明对食毒期内溴鼠灵毒理过程的阐明是一个今后可挖掘的方向。其二,除上述原因以及Vkorc1基因突变之外的其他原因导致对溴鼠灵耐受的增强。例如研究人员对南通市有抗药性的黄胸鼠、褐家鼠等家栖鼠Vkorc1基因的重要抗药突变位点检测,未发现氨基酸突变(王智泉等,2014)。类似结果也报道于广东省抗性鼠的Vkorc1基因片段检测中(姚丹丹等,2019)。这揭示了Vkorc1基因的其他位点、基因组其他基因(如细胞色素P450等),以及生物遗传差异等因素对鼠类抗药能力也有重要影响(Endepolset al.,2013)。
致谢:感谢广州市荔湾区海龙村梁锦松,以及广东省科学院动物研究所李秋剑副研究员,刘炳荣、胡少芳助理研究员等在野外工作中给予的协助。
猜你喜欢
电子科技大学学报(2022年5期)2022-10-29 01:57:52
林业科技(2021年2期)2021-07-08 05:02:50
中国生殖健康(2020年4期)2021-01-18 02:58:10
野生动物学报(2018年4期)2018-11-23 03:04:20
中国生殖健康(2018年4期)2018-11-06 07:12:16
植物保护(2018年1期)2018-05-14 12:17:22
河南农业(2017年1期)2017-02-01 16:25:21
吉林大学学报(医学版)(2015年1期)2015-12-17 07:47:24
河北农业(2015年11期)2015-04-11 07:36:36
湖北农业科学(2014年11期)2014-09-10 18:06:07
