
野化培训大熊猫幼仔的食性转换
2022-12-19 12:47:58周世强何胜山屈元元罗永吴代福黄炎李德生张和民
兽类学报 2022年6期
周世强何胜山屈元元罗永吴代福黄炎李德生张和民
(中国大熊猫保护研究中心,大熊猫国家公园珍稀动物保护生物学国家林业和草原局重点实验室,都江堰 611830)
食物、水和隐蔽物是构成野生动物栖息地结构的重要组成因子,因食物的营养成分及含量、消化率、数量和质量等存在时空差异,从而驱使野生动物形成最佳觅食策略、饮食选择特性和时空活动格局,如季节性食物变化(Seasonal dietary shift)、饮食生态位转换(Dietary niche switching)和海拔迁移模式(Altitude migration model)等(Goldizenet al.,1988;Fryxell,1991;Rubenstein and Wikelski,2003;Lambert and Rothman,2015;Houet al.,2018;Heckeret al.,2021)。同时,不同生命周期阶段的动物,通过获取不同养分来应对营养供给与需求的变化,以便获得充足的营养和能量,促进自身的健康成长,形成发育性食性转换(Developmental dietary transition)规律(Raubenheimeret al.,2012)。
大熊猫(Ailuropoda melanoleuca)为我国国宝,世界生物多样性保护的标志性物种,大熊猫国家公园的旗舰动物(薛冰洁等,2017)。大熊猫属食肉目(Carnivora)熊科(Ursidae)大熊猫属(Ailuropoda)(Sarich,1973;Swaisgoodet al.,2020),具有食肉动物特有的简单胃和短肠道,经过长期的演化,已特化成以亚高山竹类为主要食物的草食性动物,竹子占据其食物组分的99%以上(Dierenfeldet al.,1982;Schalleret al.,1985;胡锦矗,2001;潘文石等,2001;Nieet al.,2015;Weiet al.,2015)。多年来,科研人员针对不同山系野生大熊猫的主食竹种类,竹笋、竹茎和竹叶的营养成分和次生化合物,大熊猫生理机能与代谢途径,大熊猫不同季节的觅食策略和迁移习性等方面开展了深入研究,取得了大量数据,基本摸清了大熊猫的季节性食性转换规律(Schalleret al.,1985;胡锦矗等,1985;潘文石等,2001;Garshelis,2004;Nieet al.,2015;Weiet al.,2015;李亚军等,2016;王丹林等,2017;董冰楠等,2017;Huanget al.,2018;Nieet al.,2019;Wanget al.,2020;国家林业和草原局,2021;周世强,2021)。相对而言,大熊猫发育性食性转换过程的研究,因受限于野外数据获取的难度和圈养目的的影响,至今没有较为完整的证据资料,仅有片段化的描述性记录,如对邛崃山系(卧龙自然保护区)和秦岭山系(佛坪、长青自然保护区)野生大熊猫幼仔不同年龄阶段取食母乳和竹子的研究(Schalleret al.,1985;胡锦矗等,1985;Luet al.,1994;胡锦矗,2001;Zhuet al.,2001)。在人工圈养条件下,为了使母兽尽快进入下一个发情期,参加第二年的繁殖,幼仔一般在4~6月龄人工断奶,饲喂配方奶与精饲料,以及富含大量纤维素和水分的竹笋、嫩竹和竹叶等食物,此饲养模式无法获得大熊猫发育性食性转换的真实数据(Penget al.,2001;张和民等,2003;张志和和魏辅文,2006)。因此,开展大熊猫幼仔食性转换的研究,不仅可以洞悉大熊猫幼仔的觅食行为策略和生长发育规律,而且还能够为大熊猫种群、栖息地和食物资源的保护提供理论支撑。
为此,本文基于中国大熊猫保护研究中心(以下简称:熊猫中心)2010—2020年的圈养大熊猫野化培训数据,分析大熊猫行为构成和粪便组分,探讨野化培训大熊猫幼仔的发育性食性转换模式和季节性食性转换规律。目的在于:(1)利用母兽带仔野化培训期间幼仔与母兽共同生活时间较长的契机,获得大熊猫幼仔食性转换过程中较为完整的数据,清晰了解其食性转换模式;(2)比较野化培训大熊猫与野生大熊猫的发育性和季节性食性转换是否具有相似的规律,为今后提高圈养大熊猫幼仔的生存力提供参考,为建立大熊猫野化放归的技术流程与评价体系奠定基础。
1 研究方法
1.1 研究对象
2010—2020年参与野化培训的大熊猫幼仔,经专家评估后达到放归野外标准的个体,共计15只(雄性3只,雌性12只)(表1)。每只个体参与野化培训的平均时长为821 d(最短713 d,最长1 422 d);与母兽共同生活的平均时长为746 d(最短197 d,最长965 d)。4只大熊猫幼仔(“淘淘”和“华娇”,“张想”和“张梦”)的母兽分别是野外救护的大熊猫“草草”和“张卡”,其余幼仔的母兽均来自人工圈养繁殖的个体。

表1 2010—2020年母兽带仔野化培训的大熊猫幼仔Table 1 Information of pandas undergoing pre-release training while reared by mothers from 2010 to 2020
1.2 方法
在总结圈养繁殖大熊猫亚成体“祥祥”野化放归失败的基础上,熊猫中心于2008年初提出了“母兽带仔野化培训”的理念与方法(李德生等,2011;周世强等,2012;张和民等,2013)。所谓“母兽带仔野化培训”就是参与野化培训的母兽在发情配种后,首先转运至第一阶段野化培训圈(半野化圈)生活,并在该野化圈中妊娠、分娩和育幼;在幼仔生长到6月龄或1岁多之后,再将母兽和幼仔一同转移到第二阶段野化培训圈(野生大熊猫栖息地内所建野化圈),幼仔跟随母兽学习野外生存技能(空间定位、食物选择、生境利用、水源寻找、警戒逃离等);在幼仔1.5~2.5岁,经专家评估后,合格的野化培训幼仔适时放归野外(野生大熊猫小种群区域)。在野化培训期间,对大熊猫幼仔既不进行强制断奶,也不提供任何人工食物,且尽量减少人为活动对幼仔培训的影响(近距离观察或接触大熊猫幼仔时,工作人员需穿戴熊猫伪装服、涂抹大熊猫尿液或粪便),仅在秋冬季为母兽提供辅助性食物,如精饲料、竹笋等。
1.3 实验环境
1.3.1 第一阶段野化培训圈
第一阶段野化培训圈位于大熊猫国家公园卧龙片区的核桃坪区域,为大熊猫中心卧龙核桃坪基地所在地,海拔1 820 m,年平均气温8.0℃,最冷月(1月)均温-2.6℃、最热月(7月)均温16.7℃。该野化圈为半人工环境,共建有3个由室外活动场(面积2 300~3 200 m2)和内室(20 m2)所组成的野化培训圈,四周安装红外视频监控系统(Surveillance Camera Digital Monitoring System,SCDMS),用于监测大熊猫活动,圈内设有自动供水系统与流动型饮水池(图1)。
第一阶段野化培训圈内主要包括人工针叶林、落叶阔叶林与灌草丛等生境斑块。乔木层郁闭度0.1~0.6,平均胸径17~23 cm,平均树高7.5~12.0 m。灌草植物盖度25%~100%,平均高度40~70 cm。
1.3.2 第二阶段野化培训圈
第二阶段野化培训圈建于大熊猫国家公园卧龙片区的核桃坪后山和天台山两个区域,都属于大熊猫的自然栖息地并常年有野生大熊猫生活其间(图1)。
核桃坪后山区域的野化培训圈由2个适应圈构成,海拔2 050~2 500 m,平均坡度20°~30°,年平均最高气温15℃,年平均最低气温4.7℃,空气相对湿度达95%。该野化培训圈先后培训了大熊猫“淘淘”、“张想”、“新媛”、“雪雪”、“冉冉”和“潘旺”。1号培训圈面积4 hm2,四周采用铁丝网和钢板进行隔离。圈内主要分布有拐棍竹(Fargesia robusta)和短锥玉山竹(Yushania brevipaniculata)两种大熊猫主食竹种,拐棍竹林占竹林面积的95%以上。其植被类型为落叶阔叶林,是卧龙野生大熊猫的最佳栖息地(落叶阔叶林、针阔混交林和亚高山暗针叶林)之一(Schalleret al.,1985;胡锦矗等,1985;卧龙自然保护区管理局等,1987)。2号培训圈面积约24 hm2,位于1号圈的上部,气温与1号圈相似。使用电网将培训圈分成三部分,从北到南依次是A区、B区和C区。四周围栏采用高压脉冲电网和铁丝网进行隔离,圈内安装170个枪机或球机式远红外视频监控系统。圈内的大熊猫主食竹和植被类型与1号圈基本相似,圈内有2~3处常年流水的小溪并建有专门的供水系统,以备野化培训大熊猫母幼饮水之用。
天台山区域的野化培训圈被划分成面积大小各异的5个适应圈,隔断及四周采用高压脉冲电网和铁丝网分隔,并建有多个隔离笼,以便对大熊猫母兽和幼仔进行体况检查、体重监测和回捕监管。培训圈内分布有多个常年流水的溪流,能够满足大熊猫母兽和幼仔的饮水需求。野化培训圈海拔2 400~3 010 m,年平均气温4.6℃,极端最低气温-10.2℃,极端最高气温25.5℃;最大面积47.89×104m2,最小面积23.97×104m2,总面积约1 km2;坡度30°~45°。大熊猫“张梦”、“华妍”、“华娇”、“八喜”、“映雪”、“琴心”、“小核桃”、“冉冉”、“潘旺”、“钢镚儿”和“田田”先后培训于此(图1)。森林植被类型为以岷江冷杉(Abies faxoniana)、铁杉(Tsuga chinensis)、桦木(Betulasp.)、槭树(Acersp.)为优势树种构成的针阔混交林和亚高山暗针叶林,仅1号野化培训圈内有小面积的灌草丛。1、2号野化圈内分布有大量的拐棍竹林(72.17%)和冷箭竹林(Bashania faberi)(20.44%),还生长着小面积(ca.54 400 m2)的短锥玉山竹林;其余野化培训圈仅有拐棍竹作为培训大熊猫的主食竹种,占据空间面积的95%以上。
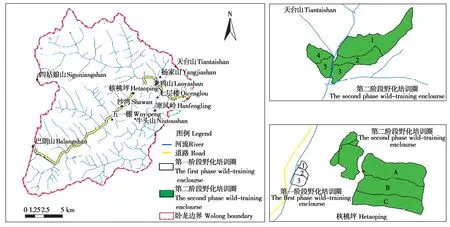
图1 母兽带仔野化培训大熊猫第一阶段和第二阶段培训圈位置示意图.图中数字和大写字母分别为面积不同的野化培训区Fig.1 Enclosure site map of mother-reared pandas undergoing pre-release training in the firth and second phase.The numbers and capital letters in the figure show the rewilding training areas of different sizes
1.4 数据采集
1.4.1 行为发育数据的采集
为了减少人类活动对培训大熊猫幼仔的干扰,在野化培训的第一阶段,研究人员以红外视频监控系统(SCDMS)对幼仔的行为进行观察。该系统可以远距离观察大熊猫的行为表现和生长发育,了解大熊猫的生存状况。SCDMS由带球机(或枪机)的高清摄像机、拾音器、通信光缆和操作平台(计算机、屏幕和软件等)组成。
在野化培训第一阶段,每天08:00—23:00对培训幼仔进行行为观察,收集从出生至6月龄(仅“淘淘”)或15月龄的行为发育数据。每天由2人查看视频或录像,以30 min(08:00—17:00)或60 min(18:00—23:00)的间隔记录大熊猫幼仔的行为信息,包括食乳、饮水、爬行、母幼交流、走动、爬树、咬玩物品(竹子和树木)、采食竹子和休息(醒着不活动或睡着)等,以及幼仔所处基质(母兽身体、地面、草丛、竹堆、木架、树干等)。每只幼仔视频收集数据平均时长达472 d(220~722 d)。
1.4.2 食性数据的采集
1.4.2.1 发育性食性转换
在野化培训第一阶段,大熊猫幼仔的觅食行为数据主要根据SCDMS进行扫描和记录。第二阶段,大熊猫幼仔的行为由项圈式声学记录装置(Acoustic Recording Unit,ARU),即音频项圈进行监测。ARU由录音机(索尼ICD-PX33M专业数字录音机)、铝盖、铝盒、皮带、防水胶带、纱布、外部麦克风和管道胶带条组成,总重量小于500 g;ARU的记录容量约为120 h,所有记录都以MP3文件类型存储,以便最大限度地提高存储容量和电池寿命(Yanet al.,2019)。为了减少大熊猫颈部摩擦和卸载过程中对记录数据的影响,每月仅选取中段72 h的数据进行行为解译,对每只大熊猫幼仔声音按每1 min监听分类,并将音频数据转换为行为数据(Yanet al.,2019)。音频数据共监听解译6 912 h,每只幼仔平均约461 h。
为了训练行为解译人员,我们选择了520 h的视频同步录音数据,采用全部事件和关键事件抽样方法进行测试。通过分析源自视频记录的行为与源自音频记录的数据之间的一致性,以及量化评估解译人员对源自音频行为数据的监听分类的准确性,统计结果表明受过训练的人员能够从声音记录中分辨出10种野化培训大熊猫幼仔的行为,音频采样法与视频采样法对于每个单独的行为具有高度的一致性,10种行为的Kappa系数均高于0.90;解译人员的行为监听准确率较高,大多数观察者相关的Kappa系数均大于0.91,单个观察者系数为0.81,行为分类的准确率达93.80%~99.50%(Yanet al.,2019)。
1.4.2.2 季节性食性转换
为了跟踪野化培训大熊猫幼仔和母兽的活动位置和生存状况,在进入第二阶段野化培训圈之前,我们给每只大熊猫佩戴了无线电颈圈(幼仔)和具有无线电发射功能的GPS颈圈(幼仔、母兽),使其适应佩戴颈圈的生活。无线电颈圈重量为50 g(幼仔),GPS颈圈重量300~350 g(幼仔)或800~1 200 g(母兽),颈圈对大熊猫的活动不会产生影响(Durninet al.,2004;周世强等,2012)。
在第二阶段野化培训圈,每月对每只大熊猫(包括幼仔和母兽)采集2~3份新鲜粪便样品进行分析,以确定其食物选择特征。由于竹子通过大熊猫的消化系统后形态特征相对不变,因此很容易分离出竹笋、竹茎和竹叶(Schalleret al.,1985;胡锦矗等,1985;Weiet al.,1999,2017)。粪便的收集和分析遵循通用的方法,即研究人员根据野化培训大熊猫幼仔和母兽的无线电信号或GPS定位信息,进入培训圈内,使用胶手套和可密封的塑料袋采集留存的新鲜粪便样品。将样品放置于60℃的恒温烤箱中烘干至恒重,并通过粪便中的干重来估计食物中所含竹笋、竹茎和竹叶的比例(Schalleret al.,1985;胡锦矗等,1985;Weiet al.,2017)。2016年3月—2018年12月,共收集150份野化培训大熊猫幼仔和母兽的新鲜粪便样品。
1.5 数据处理与统计分析
1.5.1 发育性食性转换中母乳与竹子比例的计算
以单位时间内,动物某一行为出现次数(视频观察)或出现时间(音频解译)作为该行为的计量数值(Martin and Bateson,1986;周小平等,2005)。为此,我们把在一定时间范围内(以幼仔月龄为单位时间),视频观察或音频监听中野化培训大熊猫幼仔食母乳或竹子的频次或解译时数,与视频观察或监听解译的采食行为总频次(N)或总时数(T,min)之比,作为大熊猫幼仔发育性食性转换中母乳与竹子的比例,公式如下:
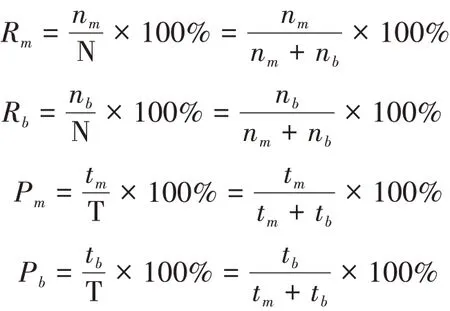
式中,Rm为大熊猫视频中食母乳占采食行为的比例(%),Rb为视频中食竹子占采食行为的比例(%),nm为视频中食母乳的频次(n),nb为视频中食竹子的频次(n),Pm为音频解译中食母乳占采食行为的比例(%),Pb为音频解译中食竹子占采食行为的比例(%),tm为音频解译的食母乳的时长(min),tb为音频解译的食竹子的时长(min),N为视频中采食行为总频次(n),T为音频解译的采食行为总时长(min)。
根据不同月龄野化培训大熊猫采食母乳和竹子比例的变化曲线,对发育性食物转换规律进行阶段划分。
1.5.2 季节性食性转换中竹笋、竹茎和竹叶所占比例的计算
鉴于第二阶段野化培训圈内主要分布的是拐棍竹,因此季节性食性转换中竹笋、竹茎和竹叶所占比例的计算主要以粪便中拐棍竹进行分析,野生大熊猫的季节性食性转换资料采用20世纪80年代的采样分析结果(Schalleret al.,1985;胡锦矗等,1985)。不同季节(月份)竹笋、竹茎和竹叶在粪样中所占百分比的计算公式如下:
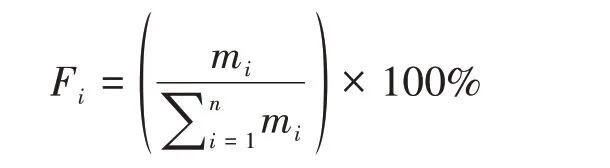
式中,Fi为竹笋、竹茎和竹叶在粪样所占百分比,mi为不同季节(月份)内竹笋、竹茎和竹叶在粪样中的测定重量(g),n为粪样中竹笋、竹茎和竹叶的类别总数。
1.5.3 数据的统计分析
野化培训大熊猫行为发育和觅食行为数据来源于2010—2020年《野化培训大熊猫行为观察记录表》、音频颈圈记录数据的解译和第二阶段野化圈内粪便样品的烘干测定。所有数据的统计、分析和图形制作都在Origin 2020软件中进行;野化培训大熊猫食母乳和竹子的比例,幼仔、母兽和野生大熊猫不同季节对主食竹竹笋、竹茎和竹叶的利用率均在Microsoft Excel 2016软件中计算。采取One-Sample Kolmogorov-Smirnov Test方法检验各项数据的正态分布性和方差齐性,对正态分布的行为发育数据和不同季节的食性转换数据(P>0.05),采用单因素方差分析(One-Way ANOVAs)和多重比较(Fisher LSD法);非正态分布的发育性食性转换数据(P<0.05),使用非参数检验(Kruskai-Wallis ANOVA法或Mann-Whitney Test法)和多重比较(Dunn’s Test法)(叶卫平等,2019)。数据以平均值±标准差(mean±SD)表示,统计检验的显著性水平设置为0.05。
2 结果
2.1 野化培训大熊猫幼仔的行为发育进程
通过红外视频监控系统(SCDMS)和现场观察,对野化培训大熊猫幼仔的行为发育时间进行了记录和统计,结果表明,与大熊猫幼仔觅食竹子和警戒逃避相关的行为,除食乳外(大熊猫幼仔通常在出生4~5 h后便可食乳),共有8种主要行为,包括爬行、走动、攀爬、玩耍物品、爬树、咬玩竹子、饮水和采食竹子(图2),行为出现时间(月龄)在不同行为间有显著性差异(df1=7,df2=86,F=43.481 1,P<0.000 1);行为出现时间与发育年龄(月龄)具有很强的时间关联性(线性拟合:Person’s=0.847 2,R2=0.717 7,P<0.000 1),依次为爬行、走动、攀爬、玩耍物品、咬玩竹子、饮水和采食竹子(图2)。野化培训大熊猫幼仔对竹子的最初采食时间平均出现在10月龄左右,一般8、9月龄时开始采食竹子(主要以竹叶为主),最早观察到5月龄时尝试采食竹叶的行为,最晚约在14月龄(图2)。此外,具有警戒逃避或进行“日光浴”的爬树行为约在5月龄出现(图2)。

图2 野化培训大熊猫幼仔的行为发育进程.图中小空方框为某一行为的平均出现时间,上下短横线为某一行为出现的最晚时间和最早时间Fig.2 The behavior development process of pre-released giant panda cubs.The small empty box in the figure is the average occurrence time of a certain behavior,and the upper and lower short horizontal lines are the latest time and the earliest time of a certain behavior
2.2 野化培训大熊猫幼仔的发育性食性转换
根据野化培训大熊猫幼仔食性中母乳和竹子所占百分比的月龄动态,发现随着大熊猫幼仔身体的发育成长,培训个体的食性变化从主要以带仔母兽的乳汁为食物来源,逐渐转换成母乳与竹子混合的食物谱,最终过渡到以培训圈内分布的主食竹为摄食对象(图3)。为此,可以将野化培训大熊猫幼仔的发育性食性转换过程划分成3个阶段:以大熊猫母兽乳汁为食的食乳期(Milk feeding period)(1~7月龄)、以食母乳—食竹子混合为食的转换期(Conversion period)(8~28月龄)和以竹子为主食的食竹期(Bamboo feeding period)(29~39月龄);其中食物转换期又可细分为关键期(Critical period)(8~18月龄)和过渡期(Transition period)(19~28月龄)两个阶段。
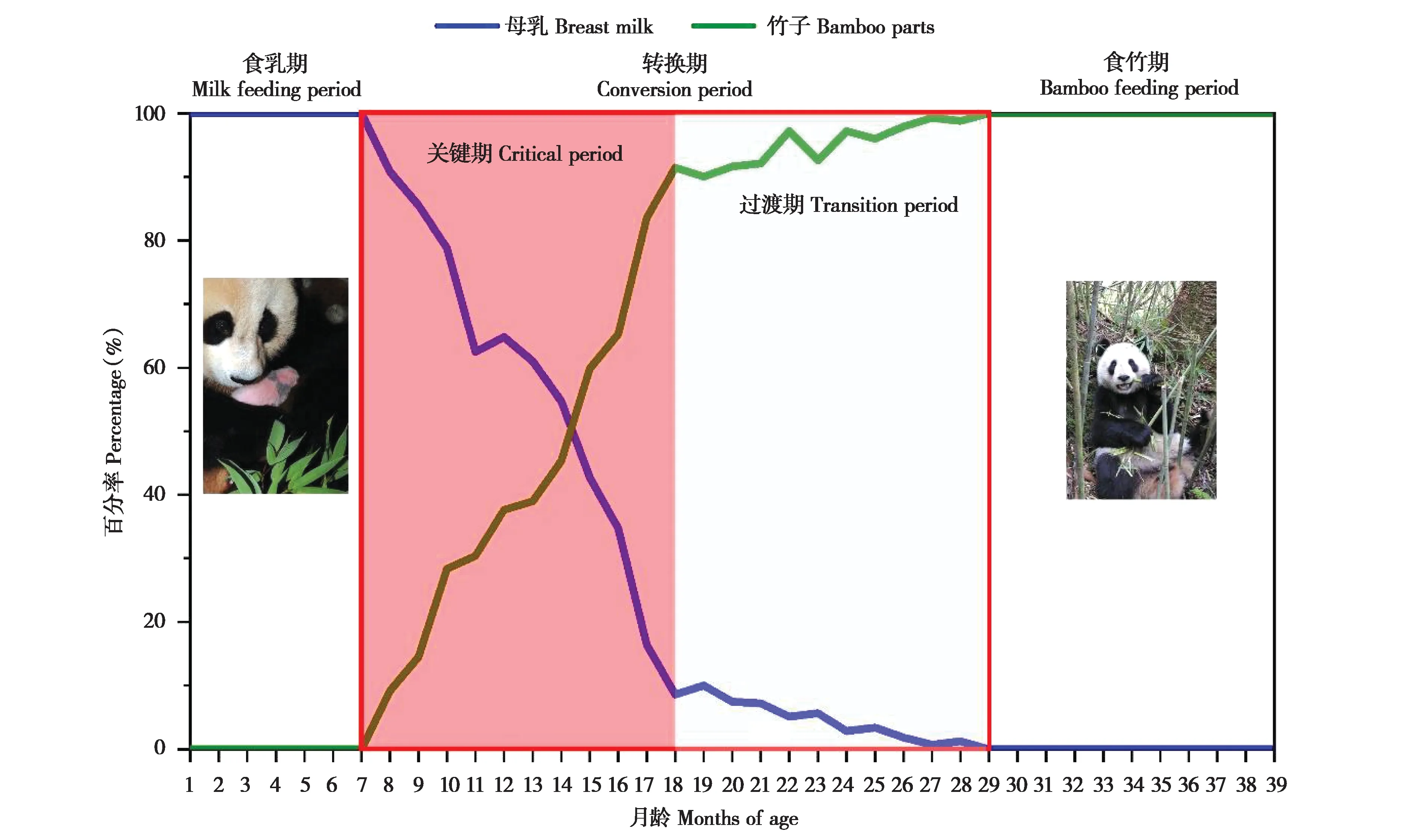
图3 野化培训大熊猫幼仔不同月龄食物组分的变化Fig.3 Changes of food components of pre-released giant panda cubs at different months of age
结果表明,以带仔母兽乳汁所占百分比计算,食乳期大熊猫幼仔食乳为100%,转换期平均为(37.39±39.55)%,食竹期为0%,三期之间及两两之间的非参数检验都差异极显著(P<0.000 1);以主食竹所占百分比计算,大熊猫幼仔食乳期为0%、转换期为(62.65±39.05)%,食竹期为100%,不同食性阶段之间及两两之间差异极显著(P<0.000 1)。如果将转换期内的两个小阶段纳入总体进行统计分析,无论是以母乳还是竹子计算,除过渡期与食竹期两者之间无显著差异之外(母乳:P=1.000 0;主食竹:P=0.982 8),总体和其他两两之间都差异极显著(P<0.000 1);关键期中母 乳 占(55.80±38.36)%、竹 子 占(44.30±37.93)%,过渡期中母乳占(4.78±6.25)%、竹子占(94.93±6.39)%。
2.3 野化培训大熊猫幼仔的季节性食性转换
野化培训大熊猫幼仔在不同季节对于竹笋、竹茎和竹叶的采食利用与带仔母兽和野生大熊猫具有相似的变化规律(图4a,b,c),表现为春季(4—6月)主要以竹笋(拐棍竹)为食,占(80.00±41.40)%;少量采食竹茎和竹叶,分别占(17.86±37.58)%和(2.14±8.00)%。夏秋季(7—10月)偶尔取食竹笋,占(19.04±40.24)%;主要以竹茎(幼嫩竹子)和竹叶为食,分别占(41.70±42.94)%和(39.25±42.36)%。冬季(11月至次年3月)主要取食竹叶与竹茎,分别占(65.11±42.03)%和(34.89±42.03)%。
统计检验表明:野化培训大熊猫幼仔不同季节粪便中所含竹笋、竹茎和竹叶的百分比与母兽之间无显著差异,春季、夏秋季和冬季分别为P=0.302 2~0.362 4、P=0.212 9~0.516 7和P=0.433 4(图4d,e,f),与野生大熊猫比较,除春季竹叶(P=0.637 5)、夏秋季竹笋(P=0.363 7)含量之间差异不显著外,其他季节的竹笋、竹茎和竹叶含量都具有极显著差异(P<0.000 1)(图4g,h,i)。
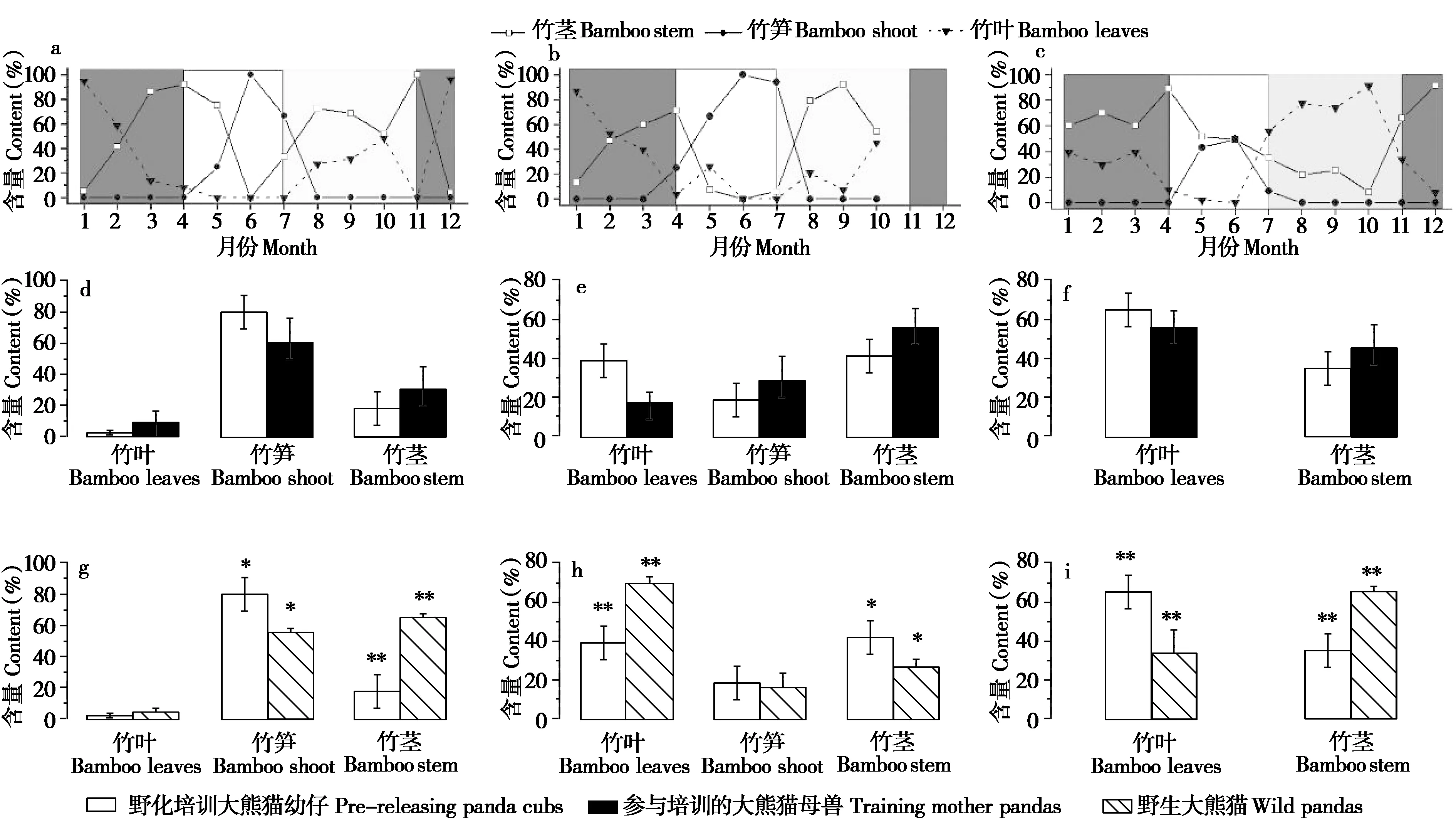
图4 野化培训大熊猫幼仔与母兽及野生大熊猫不同月份食物组分的比较.*P<0.05,**P<0.01;a:野化培训大熊猫;b:带仔母兽;c:野生大熊猫(Schaller et al.,1985;胡锦矗等,1985);d:野化培训大熊猫幼仔与母兽春季食物的比较;e:野化培训大熊猫幼仔与母兽夏秋季食物的比较;f:野化培训大熊猫幼仔与母兽冬季食物的比较;g:野化培训大熊猫与野生大熊猫春季食物的比较;h:野化培训大熊猫与野生大熊猫夏秋季食物的比较;i:野化培训大熊猫与野生大熊猫冬季食物的比较Fig.4 Comparison of food components among pre-released panda cubs,female mothers and wild pandas in different months.*P<0.05,**P<0.01;a:Pre-released panda cubs;b:Training mother pandas;c:Wild pandas(Schaller et al.,1985;Hu et al.,1985);d:Comparison of spring diet between pre-released cubs and mother pandas;e:Comparison of summer-autumn diet between pre-released cubs and mother pandas;f:Comparison of winter diet between pre-released cubs and mother pandas;g:Comparison of spring diet between pre-released cubs and wild pandas;h:Comparison of summer-autumn diet between pre-released cubs and wild pandas;i:Comparison of winter diet between pre-released cubs and mother pandas
3 讨论
3.1 野化培训大熊猫幼仔的行为发育进程
野化培训大熊猫幼仔在4~6月龄之前与圈养大熊猫幼仔都是由繁殖母兽进行哺乳和育幼,因而具有相似的生长发育规律,只是所居小环境不同(前者生长于半野化圈,后者成长在人工兽舍);野化培训大熊猫幼仔采食竹子为和母兽共同生活期间的自主采食,而圈养幼仔则为人工提供、被动选择。从2010—2020年15只野化培训大熊猫幼仔的行为发育进程统计结果来看,除食乳外,其他行为(如爬行、走动、攀爬、玩耍物品、爬树、咬玩竹叶、饮水和采食竹子等)的发育时间与圈养大熊猫幼仔基本相似(Zhanget al.,1996;Penget al.,2001)。
在母兽带仔野化培训期间,个别大熊猫幼仔在发育到5月龄时就有食竹叶的行为,一般在8~10月龄时摄食竹茎和竹叶,此结论与卧龙地区野生大熊猫的研究结论相同(Schalleret al.,1985;胡锦矗等,1985)。本研究观察到1例大熊猫幼仔14月龄才取食竹子,这一现象可能与大熊猫个体发育迟缓,或视频监控期间停电,或第二阶段佩戴音频颈圈时机选择不佳,造成数据遗漏有关,提示我们今后需加强数据收集的完整性、实时性和准确性。
3.2 野化培训大熊猫幼仔的发育性食性转换
由于野生大熊猫繁殖育幼的隐蔽性和抗干扰能力弱等特点,致使难以获得野生大熊猫幼仔的生长发育、母幼关系等数据,无法了解幼仔完整的发育性食性转换过程。目前仅见卧龙地区间断性的大熊猫觅食行为特征(食乳、食竹等)描述,秦岭地区通过视频监控系统的短时间观察结果(Schalleret al.,1985;胡锦矗等,1985;Luet al.,1994;Zhuet al.,2001;潘文石等,2001)。圈养繁殖的大熊猫幼仔常常因为母兽参与下一个繁殖季节的发情配种,而在4~6月龄时被强制断奶,后续主要以人工奶、精饲料和竹子为食,无法保证发育性食性转换的自然特性(张和民等,2003;张志和和魏辅文,2006)。因此,本研究通过第一阶段的红外视频监控系统(SCDMS)和第二阶段的音频颈圈数据监听解译(ARU listening interpretation),使母兽带仔野化培训技术为大熊猫母兽哺乳、幼仔食乳、食母乳—食竹子转换规律等研究提供了条件(李德生等,2011;张和民等,2013;Yanet al.,2019)。
通过分析野化培训大熊猫幼仔不同月龄食母乳和竹子行为比例,发现大熊猫幼仔的发育性食性转换过程大致划分为3个阶段:食乳期(1~7月龄)、食母乳—食竹子转换期(8~28月龄)和食竹期(29~39月龄);食母乳—食竹子转换期细分成关键期(8~18月龄)和过渡期(19~28月龄)两个小阶段,统计检验表明关键期与过渡期、食乳期和食竹期之间具有极显著差异,而过渡期与食竹期之间差异不显著。由此可以看出,野化培训大熊猫与野生大熊猫的断奶时间显著不同,野生大熊猫母兽大约在幼仔10~11月龄时就停止哺乳(Schalleret al.,1985;胡锦矗等,1985),而野化培训大熊猫则大多数在8~10月龄才开始少量采食竹叶,18月龄时母乳仍占据其食物组分较大比例(44.30±37.93)%,8~14月龄母乳高于竹子,15~18月龄则是竹子高于母乳。造成野化培训与野生大熊猫之间断奶时间差异的原因,可能与野外难以近距离跟踪观察、无线电颈圈信号强弱无法准确判定采食行为,以及很难捕捉到母兽和幼仔生活在一起的时机等有关,致使觅食行为等相关数据缺失;也可能与野化培训圈的空间范围较小,竹子资源有限,导致大熊猫母兽与幼仔常常生活在一起,大熊猫幼仔对母兽具有较强的依赖性,对此需要进一步的研究。从过渡期[竹子平均占(94.93±6.39)%]与食竹期(100%)之间无显著性差异来看,18月龄后大熊猫幼仔可以离开母兽独立生活,这从培训期间部分母兽发情、驱离幼仔和因母兽意外死亡、幼仔仍生存良好的现象得到佐证,并且与野外大熊猫的研究结论一致(Schalleret al.,1985;胡锦矗等,1985)。
3.3 野化培训大熊猫幼仔的季节性食性转换
竹笋在大熊猫食物组分以及生长发育、繁殖生理等方面具有重要性,将竹笋、竹茎和竹叶作为分析大熊猫食性转换中主食竹资源的组分,以洞悉野化培训大熊猫幼仔的季节性食性转换规律,及其与带仔母兽、野生大熊猫食性转换特性的异同。统计野化培训大熊猫幼仔不同月份粪便中竹笋、竹茎和竹叶的含量以及与带仔母兽和野生大熊猫的比较,结果表明,野化培训大熊猫幼仔的季节性食性转换随着季节的变化具有与带仔母兽、野生大熊猫较为相似的动态格局,即野化培训大熊猫幼仔在主食竹新笋萌发的春季主要取食竹笋,夏秋季以嫩竹茎和竹叶为食,冬季采食竹叶与竹茎。该结论不仅与圈养大熊猫的季节性觅食行为变化相似(Dierenfeldet al.,1982;张和民等,2003;张志和和魏辅文,2006;Hansenet al.,2009),而且与不同山系大熊猫的季节性食性转换规律一致(Schalleret al.,1985;胡锦矗等,1985;Weiet al.,1999,2017;胡锦矗,2001;潘文石等,2001;Nieet al.,2015;Liet al.,2016),从而说明母兽带仔野化培训方式的可行性和科学性。同时,野化培训大熊猫的季节性食性转换规律也佐证了圈养动物虽然为了适应人工环境,其遗传、生理、形态和行为等在一定程度上有所改变,甚至某些功能丧失,但经过放归前的“野化训练”或阶段性适应,可以获得或激发野外环境生存的技能和本能(Kleiman,1989;Sarrazin and Barbault,1996;Vickery and Mason,2003;Tenhumberget al.,2004;Mcphee and Carlstead,2010;Pérezet al.,2012;Germanoet al.,2015)。食物为动物生存和种群繁衍的物质基础,对食物的获取和选择是形成动物最佳觅食策略的行为体现(Goldizenet al.,1988;Fryxell,1991;Rubenstein and Wikelski,2003;Lambert and Rothman,2015;Houet al.,2018;Heckeret al.,2021),因此,野化培训大熊猫幼仔觅食行为的季节性转换规律不仅是评估野化培训成效的依据,也是培训大熊猫放归自然后能否健康生存的预测指标。
驱使野化培训大熊猫幼仔形成季节性食性转换的因素主要包括两个方面,一是跟随母兽的行为学习,探寻竹笋、竹茎和竹叶的可食性;二是与野生大熊猫和圈养大熊猫具有相同的原因,就是竹笋、竹茎和竹叶中的营养成分和次生化合物的季节变化以及大熊猫对这种变化的进食反应,这可能是最为主要的动力机制(Dierenfeldet al.,1982;Schalleret al.,1985;胡锦矗等,1985;Weiet al.,1999,2017;胡锦矗,2001;潘文石等,2001;Hansenet al.,2009;Nieet al.,2015;Liet al.,2016;Wanget al.,2017,2020;Huanget al.,2018;周世强,2021)。
圈养繁殖大熊猫幼仔在母兽带仔野化培训期间获得了良好的生存训练与环境适应,随着个体的生长发育,展现出与圈养和野生大熊猫极为相似的行为发育进程和季节性觅食行为策略,以及阶段性特征明显的发育性食物转换规律,这为今后圈养大熊猫的野化放归研究带来了一些有益的启示。(1)分析野化培训大熊猫幼仔在半野化圈、野化培训圈和野外环境的行为发育与生存适应,不仅说明圈养动物环境丰容(Environmental enrichment)技术的科学性,也体现出野化培训条件多样(生境结构、食物资源、海拔梯度)的重要性,有利于形成大熊猫的自然特性(Swaisgoodet al.,2001;张和民等,2013;Yanget al.,2018),而且暗示人们可以采用在野生大熊猫小种群区域内进行“就地培训、就地放归”模式,从而提高野化培训大熊猫幼仔的环境适应性与生存力,有利于野化大熊猫从培训环境到放归环境的自然过渡,降低培训个体回捕运输、放归转运过程中的应激反应(唐丹等,2019)。(2)野化培训大熊猫幼仔发育性食性转换过程揭示了母兽带仔野化培训的必要性,因为母乳在提高初生幼仔成活率方面具有至关重要的作用,且幼仔食乳时间较长,甚至接近2.5岁(李德生等,1999;张和民等,2003;张志和和魏辅文,2006);同时野化培训幼仔食性转换中过渡期(18~28月龄)与食竹期(29~39月龄)之间无显著差异,表明可以适当缩短母兽带仔野化培训的时间长度,幼仔1.5岁后就可适时放归野外,以便减轻对野化培训圈竹子资源的利用压力、延长野化培训圈的使用寿命、提高圈养繁殖大熊猫的野化培训效率。(3)综合野化培训大熊猫幼仔的发育性和季节性食性转换特征,提示一方面培训幼仔放归野外的时间可以选择在大熊猫1.5岁(18月龄)和2.5岁(30月龄)的初春时节(3—4月)进行,此时正值大熊猫栖息地内主食竹新笋萌发,幼仔放归后有充足的优质食物资源供其选择利用;另一方面成年大熊猫母兽可能出现发情迹象(3—5月),并主动驱离幼仔,这在野化培训期间已有观察,当采取母兽分离措施后,培训幼仔仍生长发育良好。同时,也验证了大熊猫幼仔1.5~2.5岁离开母兽,独立生活的野外研究结论(Schalleret al.,1985;Luet al.,1994;胡锦矗等,1985;胡锦矗,2001;潘文石等,2001;Zhuet al.,2001)。
致谢:中国大熊猫保护研究中心卧龙核桃坪基地的技术人员,以及曾参与圈养大熊猫野化培训的工作人员为数据收集付出了艰辛的劳动;四川省南充市林业局的罗蓉同志参与了部分工作;美国佛罗里达大学野生动物生态与保护系Vanessa Hull助理教授帮助修改英文。特此致谢。
猜你喜欢
四川畜牧兽医(2022年5期)2022-06-30 10:48:16
当代水产(2021年10期)2022-01-12 06:21:02
小天使·五年级语数英综合(2021年5期)2021-07-11 11:10:31
上海包装(2019年2期)2019-05-20 09:10:56
生态文明新时代(2018年2期)2018-03-21 05:18:20
广东石油化工学院学报(2016年3期)2016-05-17 05:16:24
中国粮油学报(2016年5期)2016-01-23 02:44:49
生物进化(2010年2期)2010-01-17 03:16:56
