
9 株绵羊肺炎支原体内蒙古分离株P113 基因克隆及蛋白序列分析
2022-12-19 01:43:16戴伶俐张月梅达来宝力格李晓艳王根云赵世华
畜牧与饲料科学 2022年6期
戴伶俐,王 娜,白 帆,张 帆,宋 越,张月梅,达来宝力格,李晓艳,王根云,赵世华
(1. 内蒙古自治区农牧业科学院, 内蒙古 呼和浩特 010031;2. 呼和浩特市动物疫病防控中心, 内蒙古 呼和浩特010020)
绵羊肺炎支原体(Mycoplasma ovipneumoniae,MO)是重要的羊呼吸道病原体,能引起绵羊和山羊非典型性肺炎。 患病羊只出现不同程度的临床症状,主要表现为咳嗽、流鼻涕、渐进性消瘦、腹泻和渗出性间质性肺炎[1],给养羊产业造成巨大的经济损失。MO 在我国广泛存在,主要集中在肉羊、奶羊主产区,如内蒙古、新疆、甘肃、四川等地[2]。羊只一旦感染MO,即便未出现明显的临床症状和肺部病变, 也会严重影响生产性能, 如日增重下降、胴体质量降低等[3]。 应用抗生素治疗并不能降低MO 在羊体内的定植率[4],通过抗生素治疗无法达到有效的治疗效果, 患病羊只常常出现反复发病的情况。 非典型性肺炎是影响养羊业健康发展的重要呼吸道疫病[5-6],因此,急需开发高效的诊断方法和疫苗产品,用于该病的有效防控。目前对于MO 的重要功能蛋白研究较少, 严重影响了其分子致病机制的研究以及防控技术的开发。
肺炎支原体入侵宿主造成感染的第1 步是黏附,因此,确定支原体黏附因子是防治感染的有效途径。 P97 蛋白是猪肺炎支原体的黏附因子,介导猪肺炎支原体与呼吸道纤毛黏附[7],是重要的毒力因子[8]。 根据生物信息学分析结果推测MO 的P113 蛋白与猪肺炎支原体黏附蛋白P97 直系同源,可能与MO 的黏附相关[9]。P113 蛋白C 末端具有良好的免疫原性[10],且含有与P97 蛋白类似的重复序列。 杨发龙等[11]对MO 的P113 蛋白C末端重复区域进行原核表达, 证明该区域具有良好的免疫原性。 根据序列及结构分析推测,P113 蛋白可能是MO 的黏附因子, 是潜在的毒力因子,因此,对该蛋白进行深入分析有利于进一步验证其生物学功能。 该研究对9 株MO 内蒙古分离株P113 基因序列进行扩增测序,分析P113 蛋白C 末端序列重复区域在不同菌株之间的变化情况, 为研究MO 黏附因子分子机制提供参考。
1 材料与方法
1.1 试验材料
1.1.1 MO 菌株
MO 内蒙古分离株9 株,由内蒙古自治区农牧业科学院兽医研究所分离、 鉴定, 菌株信息见表1。MO 标准株Y98,由内蒙古自治区农牧业科学院兽医研究所保存。

表1 MO 内蒙古分离株信息
1.1.2 主要试剂
支原体培养基, 青岛海博生物技术有限公司产品;马血清,北京索莱宝科技有限公司产品;细菌基因组DNA 提取试剂盒,天根生化科技(北京)有限公司产品;2×高保真PCR Mix 预混液、氨苄西林、琼脂糖、PCR 产物胶回收试剂盒,购自生工生物工程(上海)股份有限公司。
1.1.3 主要仪器设备
恒温摇床(THZ-98AB,上海一恒科学仪器有限公司),PCR 仪(Verit,ABI 公司),凝胶成像系统(Universal HoodⅡ,Bio-Rad 公司), 电泳仪(DYCP-31DN,北京六一生物科技有限公司),生物安全柜(AirstreamRA2,Esco 公司)。
1.2 试验方法
1.2.1 MO 培养及基因组DNA 提取
按照支原体培养基说明书配制培养基, 按照1∶10 比例接种MO 菌液, 在37 ℃、150 r/min 恒温摇床培养箱中连续培养至培养基颜色变为橙黄色;收集培养产物,4 ℃、13 000 r/min 离心20 min,弃尽上清液, 菌体沉淀用于提取基因组DNA,操作过程按照细菌基因组DNA 提取试剂盒说明书。获得的各菌株基因组DNA 保存于-20 ℃,备用。
1.2.2 P113 基因克隆与测序
根据GenBank 公布的支原体GZ-QX1 株P113 基因序列 (GenBank 登录号:KR270152.1),使用Oligo 7.37 软件设计P113 基因片段扩增引物,上游引物P113-F:5′-CGCGGATCCGCGGCAAA ACAAGATTACTAT-3′;下游引物P113-R:5′-CCGG AATTCCGGTTATTCAAACTCTTTTAG-3′。
以各MO 菌株基因组DNA 为模板, 扩增P113 基因片段。 PCR 反应体系:2×高保真PCR Mix 预 混 液25 μL,P113-F (10 μmol/L)1 μL,P113-R(10 μmol/L)1 μL,基因组DNA 模板1 μL,用无菌无酶水补至50 μL。 PCR 扩增条件为:95℃预变性5 min;95 ℃变性30 s,55 ℃退火30 s,72 ℃延伸80 s,30 个循环;72 ℃最终延伸10 min;4 ℃保温。 PCR 阳性对照模板为MO Y98 株基因组DNA,阴性对照模板为超纯水。PCR 产物经过1%琼脂糖凝胶电泳, 切下目的片段凝胶块,用PCR 产物胶回收试剂盒回收目的片段, 纯化PCR 产物送生工生物工程(上海)股份有限公司测序。
1.2.3 P113 蛋白序列分析
测序获得的各菌株P113 基因序列用NCBI 在线BLSAT 软件进行初步序列比对。 从NCBI 下 载3 株MO 菌 株ATCC 29419 (Gen-Bank 登录号:KR021380.1)、GZ-QX1(GenBank登 录 号:KR270152.1)、NCTC 10151 (GenBank登录号:LR215028.1)的P113 基因DNA 序列,采用DNASTAR 和MEGA 7.0 软件将各菌株P113 基因序列翻译为氨基酸序列并分析其进化关系。
2 结果与分析
2.1 P113 基因片段扩增结果
以不同MO 分离株基因组DNA 为模板扩增P113 基因片段, 各菌株扩增产物大小不完全一致,片段长度介于1 100~1 300 bp(见图1),提示P113 基因具有一定的多样性。

图1 MO 不同分离株P113 基因片段PCR 扩增结果
2.2 P113 蛋白序列比对结果
将测序获得的不同菌株P113 基因片段序列翻译为氨基酸序列进行比对,与NCBI 下载的3 株MO 菌 株ATCC 29419、GZ-QX1、NCTC 10151 的P113 蛋白序列相比,9 株MO 内蒙古分离株P113蛋白序列存在缺失或者插入。 缺失或者插入的序列存在一定的规律,P113 蛋白序列内部存在重复序列,以KKAEGA(S)QNQG 为主要重复序列单元(见图2)。

图2 截短P113 蛋白序列比对结果
不同菌株包含的KKAEGA (S)QNQG 重复序列单元数量不完全相同。 从NCBI 下载的3 株MO菌株P113 蛋白序列中, 该重复序列单元含有10个;MO 内蒙古分离株NM01-MO、CK-MO、NM03-MO、DMQ-ZM-MO 所含重复序列单元数量均高于10 个, 其余分离株所含重复序列单元数量较少,出现序列缺失(见表2)。

表2 MO 内蒙古分离株及参考菌株P113蛋白C 末端氨基酸重复序列单元数量 单位:个
2.3 P113 蛋白序列进化树分析
MO 不同分离株P113 蛋白序列进化树分析结果表明,ATCC 29419、GZ-QX1 和NCTC 10151的遗传距离最近,内蒙古分离株DMQ-ZM-MO 和LK-MO 与前述3 株菌的遗传距离最近,同属一个分支;D00188-MO、SZW03-MO 和DMQ-BT-MO与其他菌株遗传距离较远(见图3)。
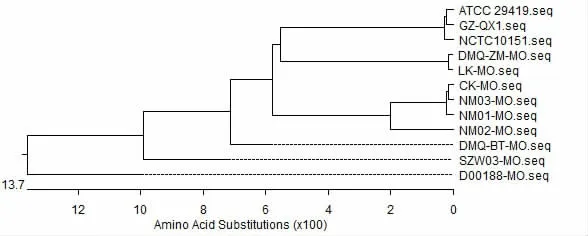
图3 截短P113 蛋白序列遗传进化树
3 讨论
黏附蛋白是支原体侵害宿主机体的关键毒力因子之一[12]。 目前已有MO 分子致病机制相关研究[13-16],但通过探究MO 关键蛋白功能来解析其分子致病机制方面的研究还十分有限。
由于MO 的P113 蛋白与猪支原体黏附因子P97 蛋白存在较高的同源性, 且C 末端都具有重复序列,因此,推测P113 蛋白是MO 潜在的黏附因子,但是目前缺少其直接影响MO 黏附作用的证据。已有研究证明MO 的P113 蛋白具有良好的免疫原性,具有开发为亚单位疫苗的潜力[10,17]。 由于P113部分基因序列较为保守,属于MO 特异性序列,研究者们已经据此研发相应的检测方法,例如:制备P113 蛋白单克隆抗体,用于MO 间接免疫荧光检测和血清抗体检测[18];建立基于P113 基因的常规PCR 检测方法和荧光定量PCR 检测方法[19-20]。
该研究通过对2017—2022 年在内蒙古地区分离的MO 菌株P113 基因片段克隆测序,发现不同菌株P113 基因片段长度存在一定差异,这种差异主要是由于C 末端重复区域的长度不一致而引起。 该研究发现,MO 内蒙古分离株P113 蛋白C末端重复序列单元为KKAEGA(S)QNQG,不同分离株的重复数量并不完全相同,但是这种差异与MO分离株分离的时间和地点没有明显的相关性,可能与菌株自身的生物学特性相关。由于P113 蛋白是潜在的毒力因子,因此,重复序列单元的数量可能与MO 的毒力强弱有关,这有待于进一步研究。
4 结论
成功扩增9 株MO 内蒙古分离株的P113 基因片段,这些菌株的P113 蛋白C 末端重复序列单元数量存在一定差异,该结果可为后续研究MO P113蛋白生物学功能以及MO 致病机制提供参考。
猜你喜欢
今日农业(2021年11期)2021-08-13 08:53:24
少儿美术(快乐历史地理)(2020年3期)2020-07-24 09:02:32
草原歌声(2019年2期)2020-01-06 03:11:12
兽医导刊(2019年1期)2019-02-21 01:14:26
兽医导刊(2019年1期)2019-02-21 01:13:58
草原歌声(2018年2期)2018-12-03 08:14:06
草原歌声(2017年3期)2017-04-23 05:13:44
妈妈宝宝(2017年3期)2017-02-21 01:22:14
兽医导刊(2016年12期)2016-05-17 03:51:45
遗传(2014年3期)2014-02-28 20:58:49
