
大白菜COBRA 基因家族鉴定及其低温胁迫表达分析*
2022-12-16 02:29:00廉瑞婷陈连水汤晓辛
云南农业大学学报(自然科学) 2022年6期
廉瑞婷,任 宏,翟 文,陈连水,汤晓辛
(1.贵州师范大学 生命科学学院,贵州 贵阳 550025;2.东华理工大学 化学生物与材料科学学院,江西 南昌 330013)
大白菜(Brassica rapaL.)是十字花科(Brassicaceae)芸苔属(Brassica)蔬菜,起源于中国[1],根据生长形态可分为结球型和非结球型[2]。大白菜原分布于中国华北,目前在中国各地广泛栽培,耐寒,喜好冷凉气候,不适于栽植在排水不良的黏土地上;其营养丰富,菜叶可供炒食、生食、盐腌和酱渍,外层脱落的菜叶可作饲料,且具有一定的药用价值[3]。大白菜是中国传统的重要蔬菜,年种植面积约180 万hm2,约占中国蔬菜种植面积的15%[4]。
大白菜COBRA基因编码糖基磷脂酰肌醇锚定蛋白(glycosylphosphatidylinositol- anchored protein),在植物细胞壁纤维素的合成、细胞的定向伸长和纤维素的沉积过程中起重要作用[5],其结构特点是:(1) N-端信号肽将该蛋白质定位于内质网;(2)碳水化合物结合区域(CBM)富含芳香族氨基酸残基;(3)高度保守的半胱氨酸富集区(CCVS);(4)在C-端具有GPI 锚定位点和疏水尾巴。COBL 蛋白 (即COBRA 蛋白) 被分泌到高尔基体囊泡中,然后位于质膜上,以调节纤维素的组装和结晶[6]。根据COBRA 蛋白序列相似性可将其分为2 个亚组,其中一个亚组的N 端氨基酸序列较长,比另一组约长45%,外显子数量较少[7]。COBRA基因最早发现于拟南芥根细胞异常膨大的突变体中[8]。拟南芥中的AtCOBL4是次生细胞壁纤维素合成所必需,其突变会使茎秆易折断[9];AtCOBL9则会影响拟南芥根毛的生长和形态发育[10];COBL2 蛋白功能缺失会使拟南芥种子表皮黏液分泌细胞中的纤维素大幅减少[11-12]。水稻Os-BC1L4(COBRA-like 蛋白)突变会导致叶片细胞壁纤维素含量降低、果胶含量增加[13];玉米中COBRA-like 蛋白bk2突变会使植株变得脆弱,茎和叶中的纤维素含量显著降低,木质素增加[14]。
在拟南芥、玉米、水稻、番茄和大豆等植物中鉴定了COBRA 基因家族成员的数量分别为12、11、9、17 和24 个[15-19],但大白菜COBRA 基因家族的研究尚未有报道。纤维素含量与蔬菜产量、食用口感、营养品质和栽培方式等密切相关。研究表明:温度对细胞壁合成物质和纤维素含量有一定的影响。35 ℃高温下,非结球白菜叶片的纤维素含量增加,而在4 ℃低温时降低[20];15 ℃处理下,烟草中的纤维素含量低于25 ℃,但木质素含量高于25 ℃[21]。本研究利用生物信息学方法对大白菜全基因组中的COBRA 基因家族进行鉴定,并对其基因序列结构、系统进化、启动子元件和低温胁迫下的表达模式进行分析,以期为深入研究 COBRA 家族各成员功能以及低温环境下COBRA基因对植物纤维素含量的调控提供理论基础。
1 材料与方法
1.1 COBRA 基因家族鉴定
从十字花科植物数据库(http://brassicadb.cn/)获取大白菜(B.rapa,AA)[22]、拟南芥(Arabidopsis thaliana)、甘蓝(B.oleracea,CC)、芥菜(B.juncea,AABB)、甘蓝型油菜(B.napus,AACC)的参考基因组及相关注释信息。查询大白菜的COBRA基因,利用HMMER 程序(http://hmmer.org/)和Pfam (http://pfam.xfam.org/)结果对COBRA 蛋白保守结构域进行确认。使用 BlastP 在大白菜全基因蛋白质数据中进行搜索,筛选阈值(e-value)小于等于10-5,获得大白菜和其他4 种十字花科植物的COBRA 基因家族成员。
1.2 COBRA 基因序列结构和系统进化分析
利用 Expasy 在线分析蛋白质的氨基酸长度、分子量和理论等电点;在大白菜基因组注释信息中提取COBRA 家族成员染色体位置信息。使用MEME 程序(Version 5.3.0,http://meme-suite.org/tools/meme)分析大白菜COBRA 蛋白质序列的保守基序,迭代次数为18 000,nmotifs≤10。使用MEGA-X 软件的ClustalW 工具对蛋白质序列进行比对排序(默认参数);采用邻接法(neighbor-joining)构建系统发生树,Bootstrap 校验值为1 000 次。
1.3 COBRA 基因表达的分析
提取白菜COBRA 家族基因上游2 kb 序列作为启动子区域,用PlantCARE 软件预测大白菜COBRA 家族基因的启动子顺式调控元件。
在常温(25 ℃)下将大白菜(品种:KTRG-B49)培养至四叶一心期,然后分为以下6 个处理组:(1) 4 ℃ (低温)分别处理1、4 和6 h;(2) -2 ℃处理2 h;(3) -4 ℃处理1 h;(4) 低温处理后常温恢复1 d。各处理组均以常温(25 ℃)为对照组。所有处理均取3 个生物学重复,共7 组21 个样品。采用Illumina HiSeqTM2500 测序仪对各组处理的叶片进行转录组测序。计算每个样品每千个碱基的转录每百万映射读取的片段(fragments per kilobase of exon model per million mapped fragments,FPKM)均值,分析大白菜COBRA基因在不同条件下的表达量,将其表达量通过TBtools 软件绘制热图。
1.4 荧光定量qPCR 的基因表达分析
应用Primer3 设计引物(表1),内参基因为白菜BraActin。使用HiScript ll QRT SuperMix for qPCR (诺维赞)试剂盒对样品RNA 进行反转录得到cDNA,用ChamQ SYBR qPCR Master Mix(诺维赞)试剂盒进行定量PCR 试验。反应体系为20 μL,包括:cDNA 2 μL,上、下游引物各0.4 μL,SYBR 10 μL,灭菌ddH2O 补至20 μL。PCR 反应程序为:95 ℃ 30 s,95 ℃ 10 s,60 ℃22 s,40 个循环。基因的相对表达量以2-ΔΔCt法计算,用Excel 软件整理数据并绘图。

表1 qPCR 引物信息Tab.1 Primer sequence of qPCR
2 结果与分析
2.1 大白菜COBRA 基因家族的鉴定及理化性质
大白菜基因组COBRA 家族含有23 个成员(表2),命名为BrCOBL1~BrCOBL23,分布在大白菜基因组的8 条染色体上(图1)。由表2 可知:BrCOBL 蛋白质的分子量在8.61~74.53 ku之间,大部分成员的相对分子量在45~55 ku 之间;氨基酸长度在98~673 aa 之间,有73.91% 成员的氨基酸长度大于400 aa;等电点在5.16~10.41 之间,有86.96% 成员的等电点大于7。鉴定出的大多数COBRA基因具有CCVS 基序、N 端信号肽和潜在的ω 位点。
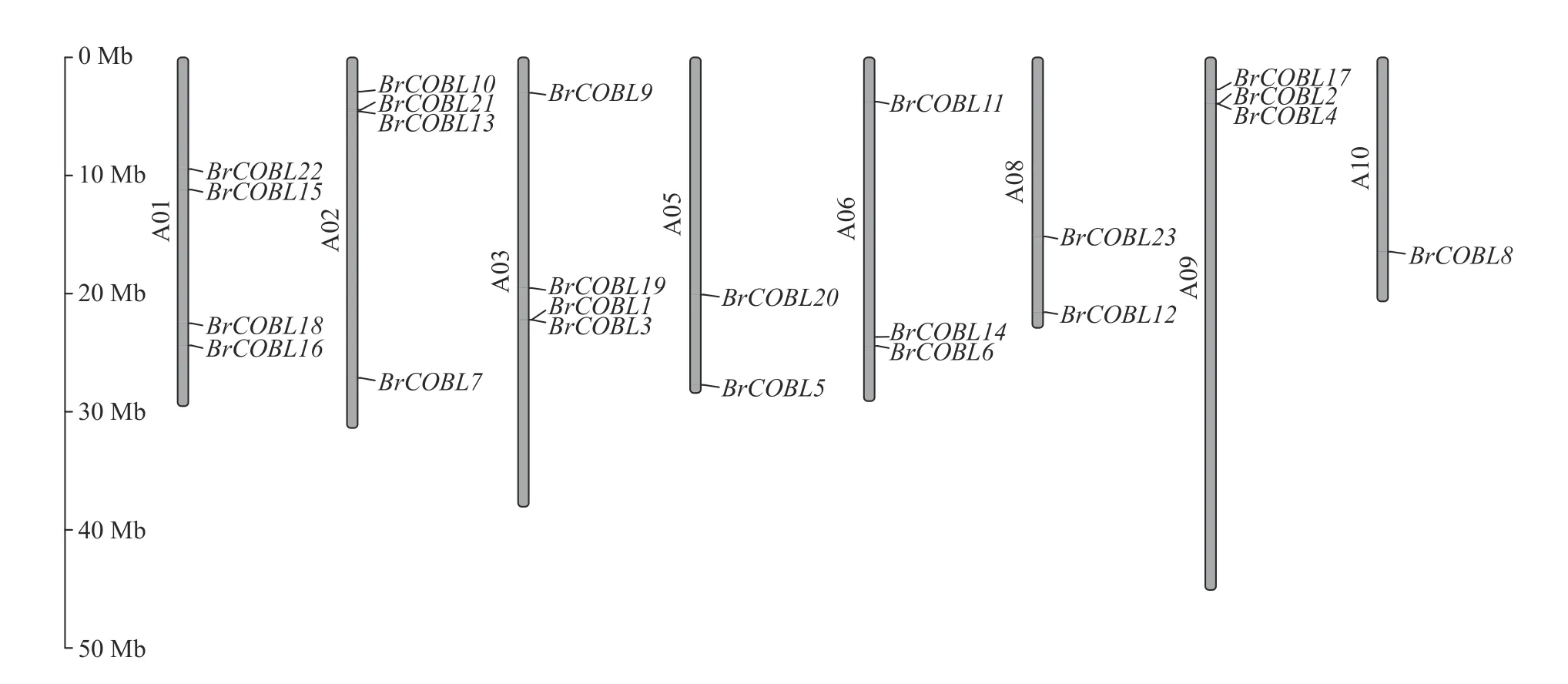
图1 大白菜COBRA 基因家族在染色体上的分布Fig.1 Distribution of COBRA gene family of Brassica rapa on chromosomes
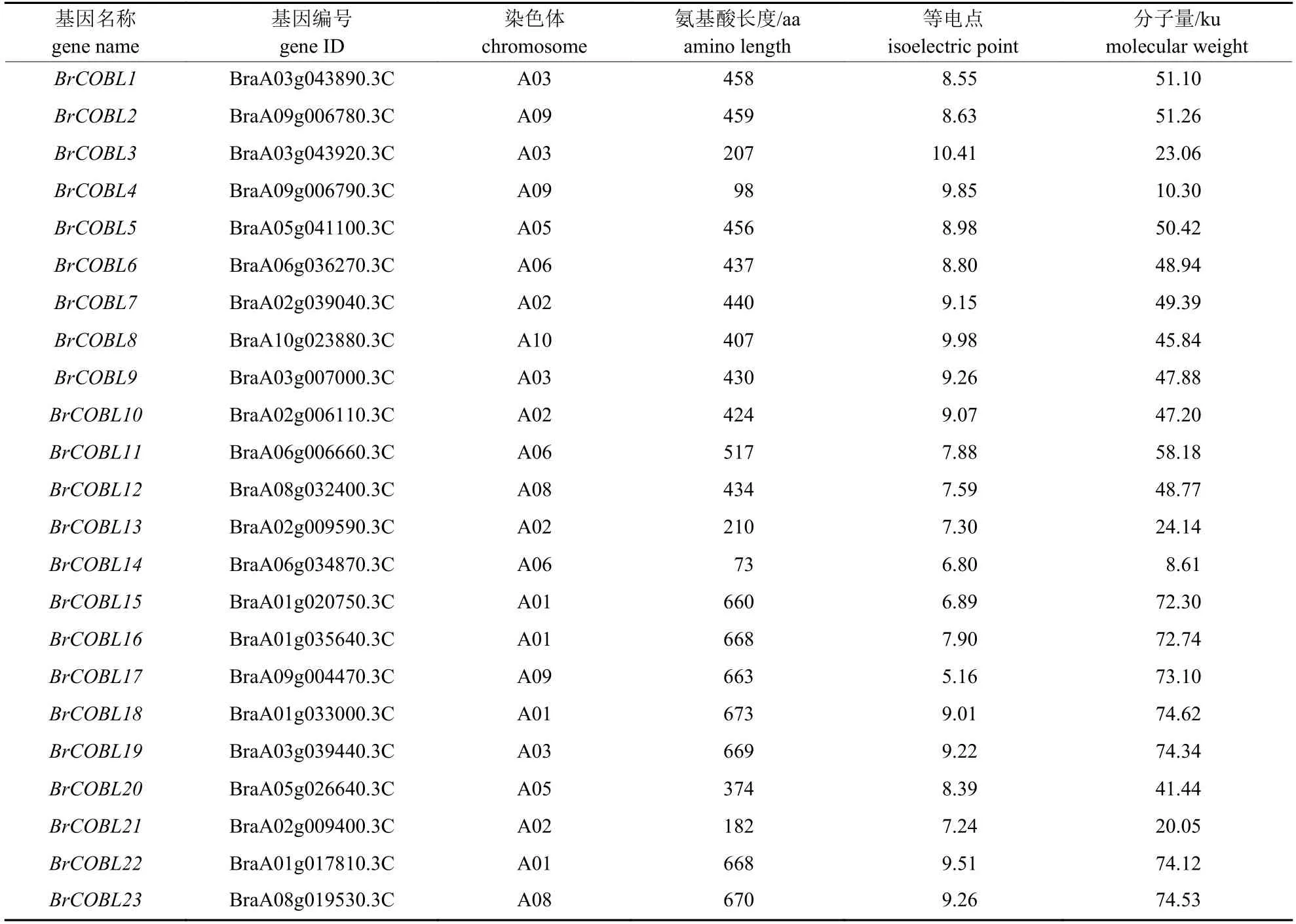
表2 大白菜COBRA 基因家族成员的理化性质Tab.2 Physicochemical properties of COBRA gene family members of Brassica rapa
2.2 大白菜COBRA 基因家族的序列结构及保守基序
由图2 可知:大白菜COBRA 基因家族可分为2个亚家族(Group1 和Group2),Group1 有13 个成员,包括BrCOBL1~BrCOBL13,Group2家族有10 个成员,包括BrCOBL14~BrCOBL23。对大白菜23 个COBRA 家族成员的蛋白质保守基序进行分析(图2a 和表3)可知:除BrCOBL3、BrCOBL4 和BrCOB20 外,其他20 个蛋白都具有 motif2;除BrCOBL4、BrCOBL13 和BrCOBL14外,其他20 个蛋白均含有motif6,说明motif2和motif6 为 COBRA 家族的特征motif。此外,有19 个蛋白含有 motif1,有19 个蛋白含有 motif3,有18 个蛋白含有 motif5、motif7 和motif8,说明这些motif 在COBRA 蛋白中也相当保守,其中motif5 含有Cys-Cys-Val-Ser (CCVS)结构域。Group1 中除BrCOBL4 外,其他均含有motif2 和motif3;Group2 中除BrCOBL14 外,其他均含有motif1、moti5、motif6 和motif9,且motif4 只存在于Group1,motif9 和motif10 只存在于Group2,表明这些motif 可能与各亚族成员的功能分化有关。

表3 大白菜COBRA 基因家族的motif 序列及长度Tab.3 Motif sequence and length of COBRA gene family of B.rapa
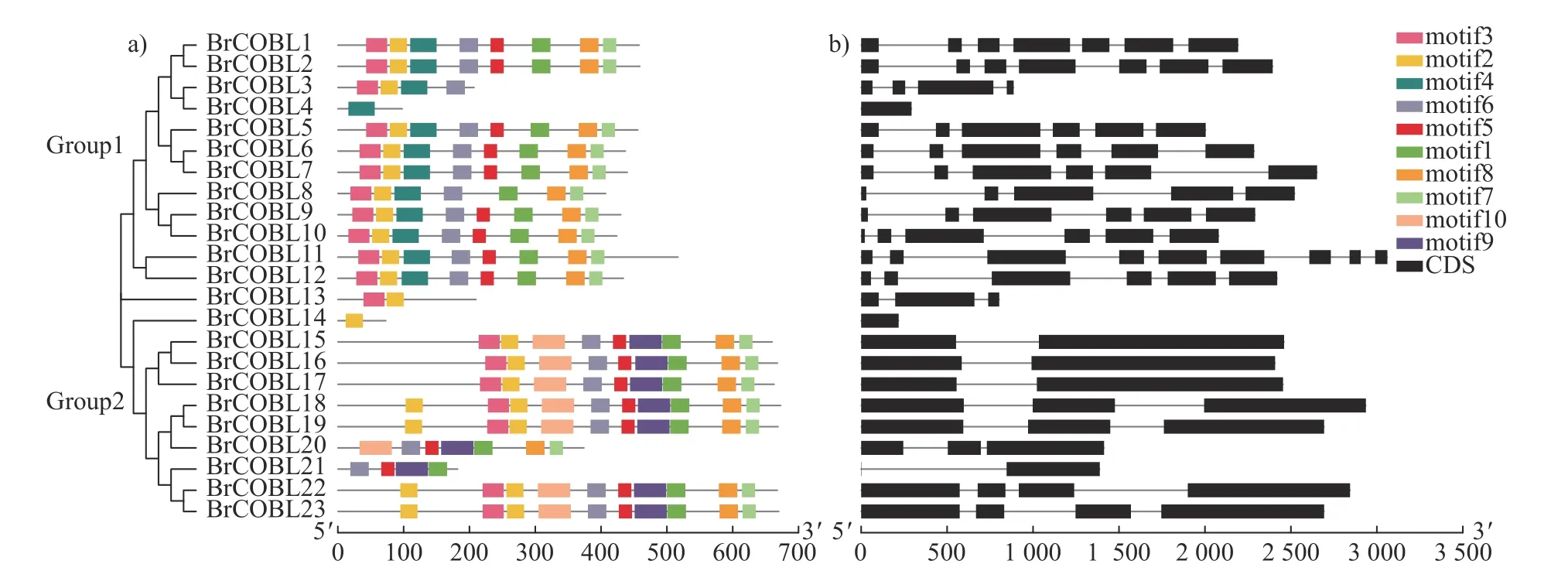
图2 大白菜COBRA 基因家族系统进化及基因结构Fig.2 Phylogenetic evolution and gene structure of COBRA gene family of B.rapa
对大白菜COBRA 家族成员的基因结构分析(图2b)可知:Group1 家族的外显子数量较多。COBRA 家族中有10 个成员含有5 个及5 个以上外显子,它们都位于Group1。Group2 家族中外显子数量较少,其中8 个成员含有1~3 个外显子,2 个成员含有4 个外显子。结合系统进化分析可知:进化关系较近的基因其结构也相似,进一步表明系统进化树聚类的可靠性。
2.3 大白菜近缘物种的COBRA 蛋白系统进化
由表4 可知:十字花科的5 个物种共鉴定COBRA 成员147 个。根据氨基酸序列相似性构建的系统发生树(图3)可将基因组成员分为8 个不同的亚组(I~ VIII)。I 亚组所含成员最多,大白菜有6 个成员在I 亚组(BrCOBL5~BrCOBL10),占总数的18.75%,这6 个基因跟拟南芥的3 个基因(AT3G02210、AT3G29810和AT5G15630)位于同一亚组,推测它们可能有类似的功能;BrCOBL1和BrCOBL2位于第II 亚组;BrCOBL3和BrCOBL4位于第III 亚组;BrCOBL11和BrCOBL12位于第V 亚组;BrCOBL18、BrCOBL19和BrCOBL20位于第 VI 亚组;BrCOBL15、BrCOBL16和BrCOBL17位于第VII 亚组;BrCOBL21、BrCOBL22和Br-COBL23位于第VIII 亚组。
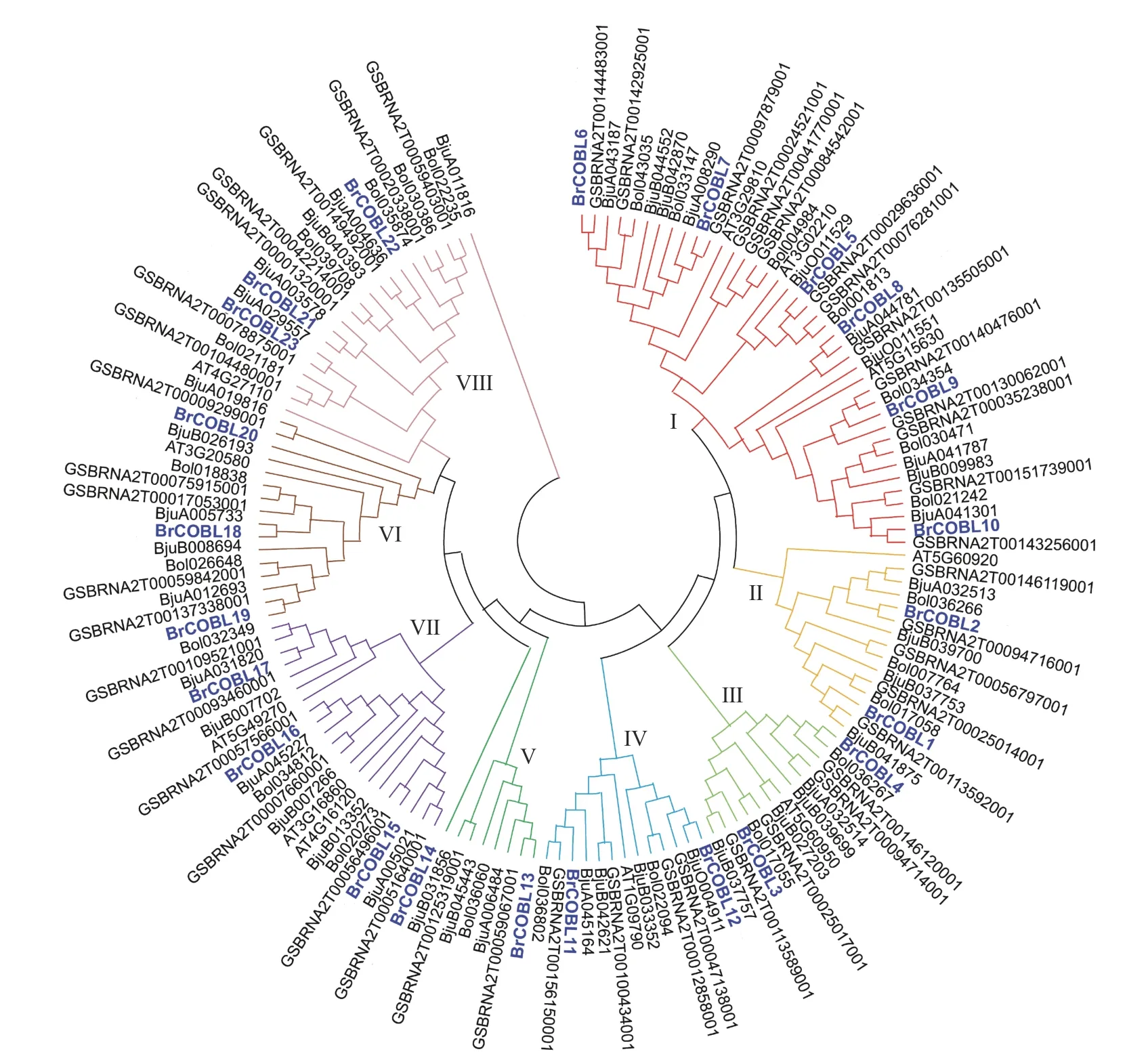
图3 十字花科物种的COBRA 家族系统发生树Fig.3 COBRA family phylogenetic tree of Brassicaceae species

表4 系统发生树中的亚组Tab.4 Subgroups in phylogenetic tree
2.4 大白菜COBRA 基因家族对低温胁迫的响应
由图4 和图5 可知:BrCOBL1、BrCOBL2、BrCOBL5、BrCOBL10、BrCOBL15和BrCOBL16基因均在大白菜叶片中表达,但另外17 个基因在叶片中几乎不表达,推测大白菜COBRA基因功能可能存在冗余。BrCOBL15和BrCOBL16基因在常温下表达量较低,在低温处理下表达量显著增高,恢复常温后,其表达量下降。BrCOBL1和BrCOBL2基因在所有样品中都有表达,尤其是在4 ℃处理6 h、-2 ℃处理2 h 和-4 ℃处理1 h的样品中表达量较高,在-4 ℃处理1 h 的样品中BrCOBL1和BrCOBL2基因表达量分别是对照的4 倍和3.5 倍;样品在常温恢复1 d 后的表达量又显著降低,说明低温胁迫能增加BrCOBL1和BrCOBL2基因的表达量,且温度越低或在低温下处理时间越长其表达量增量越大。可见,BrCOBL1、BrCOBL2、BrCOBL15和BrCOBL16基因与大白菜响应低温胁迫和低温下调控白菜纤维素合成有密切关系。
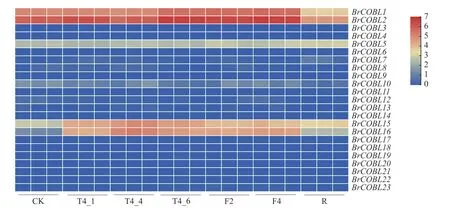
图4 应用转录组测序分析COBRA 在低温胁迫下的表达量Fig.4 Expression level of COBRA analyzed by RNA-seq at low temperature stress

图5 BrCOBL1、BrCOBL2、BrCOBL15 和BrCOBL16 在低温胁迫下的qPCR 表达Fig.5 qPCR expression of BrCOBL1, BrCOBL2, BrCOBL15 and BrCOBL16 at low temperature stress
2.5 大白菜COBRA 启动子区域顺式元件预测
由图6 可知:在15 种启动子元件中,每个COBRA 成员启动子区域都含有共同的1 个或多个TATA-box 和CAAT-box 元件;分别有73.9%、73.9%和56.5%的COBRA 启动子含有ARE、Gbox 和ABRE 元件,这些与植物抗氧化反应、光应答反应以及脱落酸和茉莉酸调控相关。Br-COBL2基因启动子区域的ABRE 元件最多(7个),而赤霉素调控元件TATC-box 在BrCOBL1中特异存在,这可能与BrCOBL1和BrCOBL2的特异性表达有关。以上结果说明COBRA基因参与大白菜的非生物胁迫响应及激素应答调控。

图6 大白菜COBRA 基因家族启动子顺式调控元件Fig.6 Cis-elements in COBRA gene family promoters
3 讨论
COBRA 基因家族是植物特有的基因家族,其主要功能是参与植物细胞的扩增,且与籽粒产量和纤维素的生物合成密切相关[23-25]。本研究基于生物信息学方法在大白菜中共鉴定到23 个COBRA 基因家族成员,从其蛋白序列整体分析结果可知:COBRA基因分为2 个亚族,同一亚族的基因具有相似的基因结构,不同亚族之间蛋白长度差异明显。在基因结构上,Group1 家族COBRA基因的外显子数量多于Group2 家族,该结果与对拟南芥[8]、水稻[24]、棉花[26]、冬枣[27]、高粱[28]和大豆[22]的研究结果基本一致。大白菜COBRA 蛋白结构较保守,有82%以上的成员含有motif2、motif6、motif1 和motif3,且2 个亚族成员也有各自特有的基序,不同亚族的保守基序不同暗示着基因功能的多样性。
在系统发生树中,遗传距离相近或在同亚组的基因可能执行类似的功能。大白菜COBRA 家族成员在低温胁迫下表达的2 个基因(BrCOBL15和BrCOBL16)均在第VII 亚组上,在这个亚组的还有AtCOBL9(AT5G49270),它会影响拟南芥根毛的生长和形态发育[10]。BrCOBL5~BrCOBL10的6 个成员在Group1 家族,与拟南芥的AtCOBL4(AT5G15630)均位于I 亚组,AtCOBL4为拟南芥次生细胞壁纤维素合成所必需,其突变会使茎秆易折断[23]。
‘cob-5’ (AT5G60950)功能缺失会严重影响植株的生长发育,同时会积累大量应激化合物(如花青素和胼胝体等),还会导致参与花青素合成的基因上调、参与细胞生长的基因下调并大量诱导防御胁迫相关的基因,如植物茉莉酸信号通路[29]。大白菜的BrCOBL3和BrCOBL4与拟南芥At-COBL5都在第III 亚组,推测它们可能有类似的基因功能。
纤维素是细胞壁的重要成分,低温会影响植物细胞壁各物质的含量或改变细胞壁的机械结构。研究表明:4 ℃低温下,非结球白菜的纤维素含量降低了22.16%[20];15 ℃处理下,烟草的纤维素含量降低而木质素含量升高[21];低温下棉花纤维的长度会显著缩短[30]。同属于第II 亚组的BrCOBL1和BrCOBL2在常温下少量表达,同属于第VII 亚组BrCOBL15和BrCOBL16在常温下几乎不表达,这4 个COBRA基因在低温时的表达量显著升高,推测与纤维素合成、细胞壁形成以及细胞的定向生长密切相关。
4 结论
低温环境下,大白菜BrCOBL1、BrCOBL2、BrCOBL15和BrCOBL16在其叶片显著表达,根据启动子分析结果推测BrCOBL基因可能参与大白菜的非生物胁迫响应及激素应答调控。COBRA 蛋白序列含有共同的保守基序,且某些基序特异存在于不同的亚组,表明这些基序可能与各亚组的功能分化有关。
猜你喜欢
动物营养学报(2024年2期)2024-03-05 12:17:22
实用心脑肺血管病杂志(2023年12期)2023-12-14 07:47:10
新医学(2023年10期)2023-12-09 15:04:51
中国-东盟博览(政经版)(2023年1期)2023-03-15 01:05:18
南方医科大学学报(2022年3期)2022-04-13 01:51:26
园林科技(2021年3期)2022-01-19 03:17:32
浙江大学学报(农业与生命科学版)(2021年3期)2021-07-10 07:07:36
心电与循环(2020年1期)2020-02-27 07:48:26
蔬菜(2016年8期)2016-10-10 06:49:14
阅读与作文(小学高年级版)(2016年3期)2016-03-08 23:38:58
