
中度锰胁迫对降香黄檀幼苗光合特性的影响
2022-12-13 11:53杨红兰黄秋婵韦玉淼冯守富
广东蚕业 2022年11期
杨红兰 黄秋婵 韦玉淼 冯守富
(1.广西民族师范学院化学与生物工程学院 广西崇左 532200;2.宜州区山谷高级中学 广西河池 546300)
锰(Mn)是植物体生长发育过程中所必需的微量元素,在植物的光合作用中,其在维持细胞器的正常结构与活化酶活性等方面有着不可替代的作用[1]。然而,土壤中有效锰含量过高时会对植物产生毒害作用,导致植物自身结构及功能的改变,也会影响植物的形态、生理特性和超微结构,还会降低光合色素含量,削弱光合作用,破坏细胞膜结构,引发渗透相关指标的变化,从而抑制植物的生长发育,对植物的生长造成不利影响[2-4]。锰在植物光合作用过程中所发挥的作用是锰在植物体内最重要的生理功能。叶绿体中分布着较多的锰,锰不仅在维持叶绿体结构上是必需的,而且能够直接参与植物光合作用过程中的光合放氧阶段[5-6]。
降香黄檀(Dalbergia odoriferaT.Chen),又叫作黄花梨、花梨木,属于豆科蝶形花亚科黄檀属半落叶常绿乔木,被列为国家二级保护植物[7]。其因材质坚硬、结构致密,耐湿耐腐蚀且伴有芳香等特性,常用来制造上等家具和工艺品[8]。此外,降香黄檀还具有止血止痛、行血活血等功效,有较高的经济价值和药用价值[9]。近些年来,随着人们生活水平的不断提高,降香黄檀产品越来越受到人们追捧和青睐,市场常常供不应求。目前我国对降香黄檀的研究主要集中在栽培技术和药用功能等方面。本研究以降香黄檀幼苗为实验材料,测定在中度锰胁迫下降香黄檀幼苗的光合参数,以了解降香黄檀在锰胁迫下的光合作用情况,为引种栽培和合理管理提供一定的理论依据。
1 材料和方法
1.1 实验材料及处理
降香黄檀成熟种子浸泡24 h,洗净沥干插播在沙基中。培养至幼苗长出两片羽叶时,选取各方面长势类似的幼苗植株移栽到口径为15 cm的塑料盆中,每盆5棵,各种植盆中的混合土壤质量相同且配比为营养土∶黑土∶红土∶沙土=5∶2∶2∶1。待幼苗长大到5片羽叶时,选取6盆各方面长势相近的降香黄檀幼苗植株,平均分为2组进行实验。实验组以临近根部填埋的方式一次埋入Mn含量为10 mg/kg的MnSO4进行胁迫处理,空白对照组以自来水浇灌。
1.2 光合参数的测定
胁迫后每隔14 d于晴朗天气采用便捷式光合测定仪测量光合参数,共测3期(以下称为早期、中期、后期)。每期测定时间从8:00到15:00,每1 h测1次。测量时,每盆选取长势相同朝向一致的3片成熟叶进行测量,每叶记录5组数据。测定的光合环境指标包括大气CO2浓度(air CO2concentration, Ca)、大气相对湿度(relative air humidity, RH)、大气温度(air tamperature, Ta)、光合有效辐射(photosynthetically active radiation, PAR),生理指标包括幼苗的净光合速率(net photosynthetic rate, Pn)、蒸腾速率(transpiration rate, Tr)、胞间 CO2浓度(intercellular CO2concentration, Ci)、气孔导度(stomatal conductance, Gs)等。
1.3 数据处理
实验数据使用Excel 2016和SPSS软件进行作图和处理。光合利用率(LUE)和水分利用率(WUE)计算公式如下:
光能利用率(light use efficiency, LUE)=净光合速率(Pn)/光合有效辐射(PAR)×100%;
水分利用率(water use efficieney, WUE)=净光合速率(Pn)/蒸腾速率(Tr)×100%[10]。
2 结果与分析
2.1 环境因子日变化
环境因子日变化曲线如图1所示。大气CO2浓度(Ca)在8:00—10:00缓慢下降,11:00后趋于稳定;大气温度(Ta)保持在30 ℃~40 ℃,波动不大。二者在3个时期的变化不大。早期大气相对湿度(RH)在10:00时有明显下降,之后迅速回升,然后逐渐趋于稳定;中期和后期大气相对湿度(RH)在8:00—10:00缓慢下降,之后趋于稳定。胁迫早期的光合有效辐射(PAR)变化曲线呈双峰型,从8:00开始逐渐下降,9:00—10:00明显上升,达到第一个峰值,第二个峰值出现在12:00。胁迫中期与胁迫后期的光合有效辐射日变化趋势相似,曲线呈单峰型。胁迫中期的光合有效辐射峰值出现在10:00,胁迫后期光合有效辐射的峰值出现在11:00。
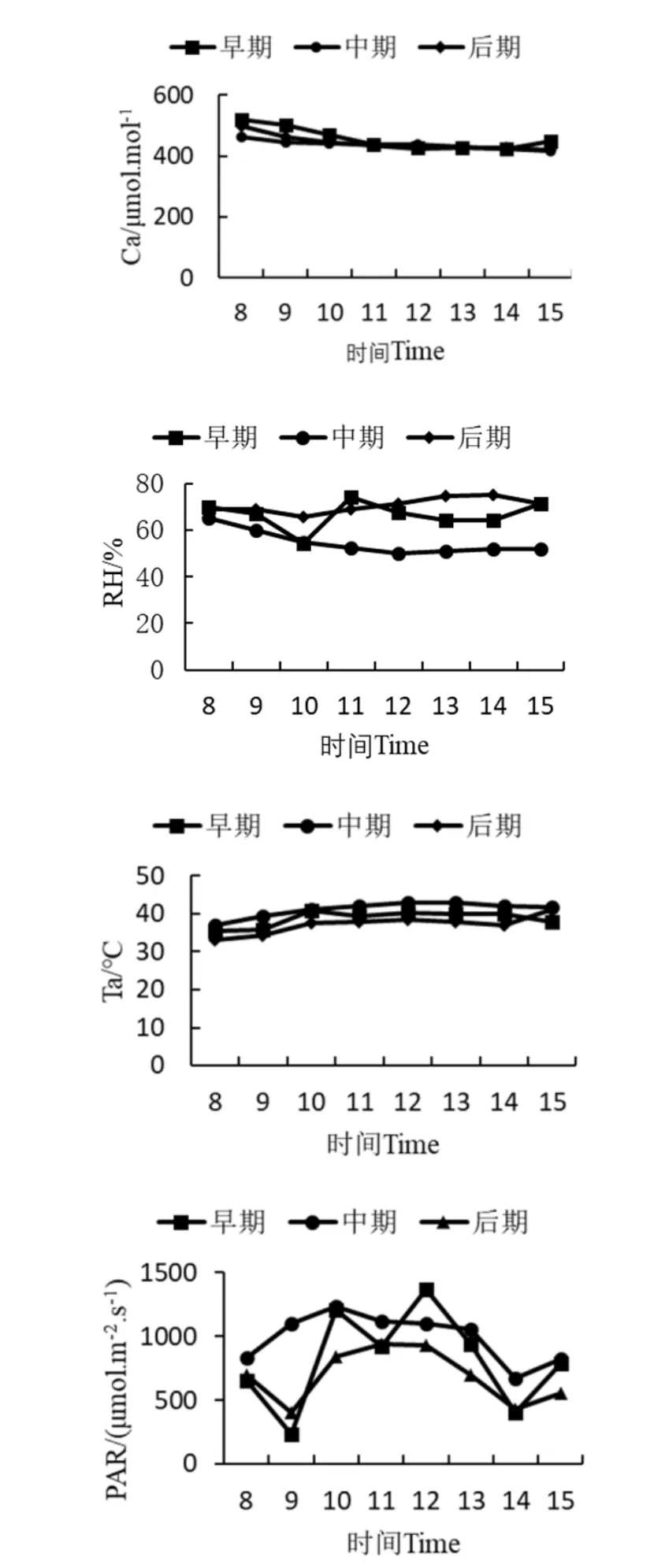
图1 降香黄檀幼苗胁迫早、中、后期环境因子日变化
2.2 中度锰离子胁迫下降香黄檀幼苗生理因子日变化
2.2.1 降香黄檀幼苗叶片净光合速率(Pn)日变化
降香黄檀幼苗叶片的净光合速率日变化曲线如图2所示。降香黄檀实验组前期和中期的净光合速率变化趋势相似,在8:00—9:00急剧下降,9:00—10:00有明显的上升趋势,而后从11:00开始下降,至12:00 Pn降到最低。12:00—13:00及14:00—15:00 Pn均呈上升趋势,此时Pn变化主要受光强影响。在后期,实验组Pn变化趋势和空白组相似,呈“W”形,出现光合“午休”现象。这三个时期中,每一个时期的净光合速率在多个时间点上都表现出实验组低于空白组。
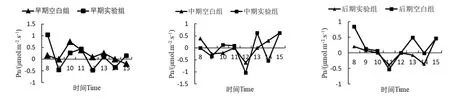
图2 降香黄檀幼苗净光合速率日变化曲线
2.2.2 降香黄檀幼苗叶片蒸腾速率(Tr)日变化
降香黄檀幼苗叶片的蒸腾速率日变化曲线如图3所示。胁迫早期,实验组Tr日变化曲线呈双峰型,峰值出现在10:00和13:00;空白组Tr日变化曲线为单峰型,峰值出现在12:00,此时正好为当天光合有效辐射的最高点,12:00后Tr开始下降,直至15:00达到最低点。胁迫中期空白组Tr日变化规律不明显,实验组Tr的日变化曲线呈单峰型,峰值出现在9:00。胁迫后期实验组Tr日变化曲线呈不明显的“W”形。综合三个时期Tr的变化规律可以看出,降香黄檀幼苗的蒸腾速率在多个时间点上都表现出实验组低于空白组。蒸腾速率能够反映出植物根系吸收水分能力的强弱,如蒸腾速率增大则表示植物根系吸收的水分增多,反之则减少[12]。由此可以看出,中度锰离子胁迫可以使降香黄檀幼苗根系吸收水分的能力减弱从而降低了降香黄檀幼苗的蒸腾速率。
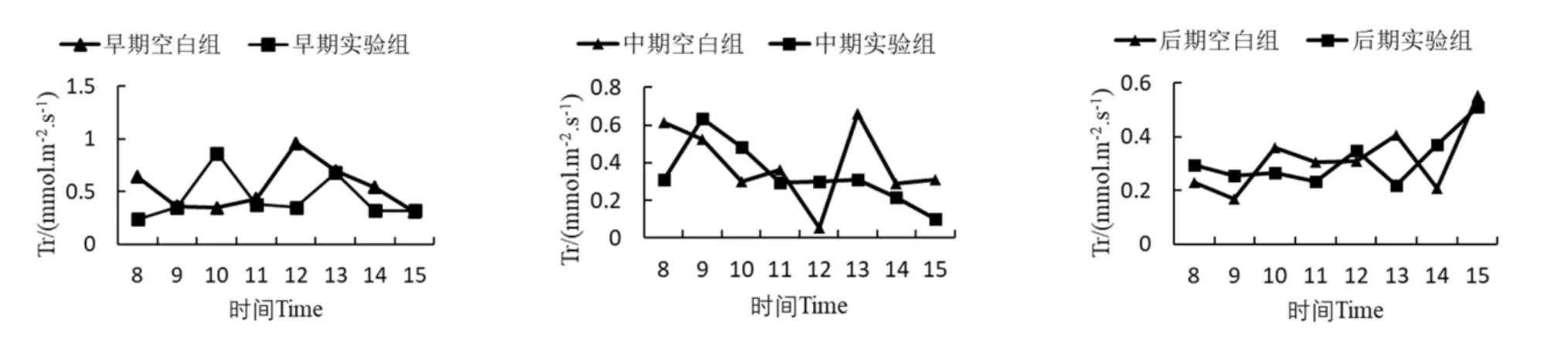
图3 降香黄檀幼苗蒸腾速率日变化曲线
2.2.3 降香黄檀幼苗叶片胞间CO2浓度(Ci)日变化
降香黄檀幼苗叶片胞间CO2浓度日变化曲线如图4所示。胁迫早期,空白组的Ci从早上8:00开始明显下降,在9:00达到当日最低值,9:00后开始上升并于10:00达到峰值,之后Ci的变化不明显;而实验组降香黄檀幼苗Ci日变化曲线呈双峰型,Ci最高值出现在13:00,最低值出现在8:00。胁迫中期,实验组和空白组的Ci日变化规律均不明显。胁迫后期空白组和实验组的Ci日变化曲线相对平缓,其中空白组呈双峰型,Ci最高值出现在9:00;实验组呈单峰型,Ci最高值出现在11:00。此外,锰胁迫后降香黄檀幼苗Ci的日平均值均明显高于空白组,在胁迫早期,实验组较空白组增加了56.4434 μmol/mol,胁迫中期实验组比空白组增加了44.1449 μmol/mol,胁迫后期实验组比空白组增加了24.2767 μmol/mol。由此可得,中度锰离子胁迫能提高降香黄檀的胞间CO2浓度。但是,胁迫时间越久,提高程度越低。
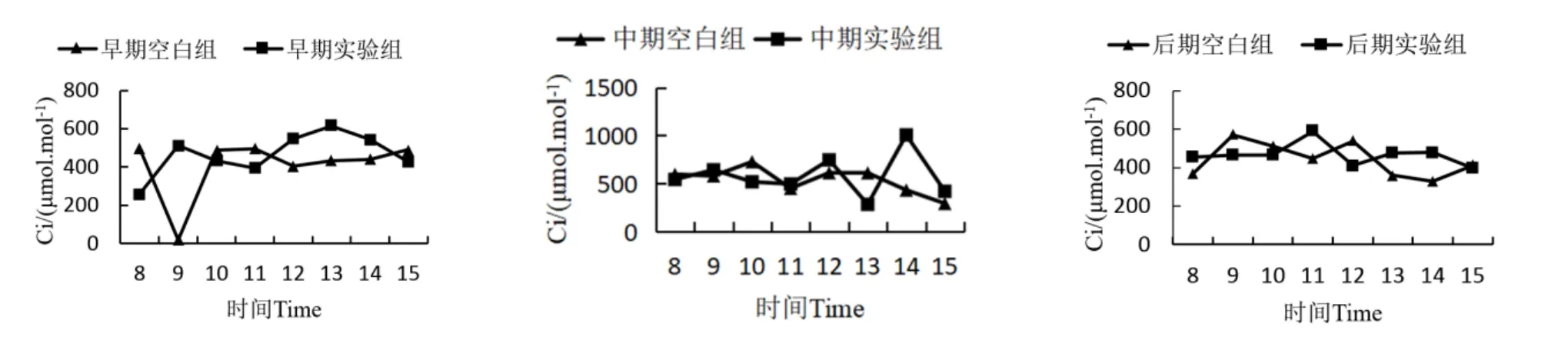
图4 降香黄檀幼苗胞间CO2浓度日变化曲线
2.2.4 降香黄檀幼苗叶片气孔导度(Gs)日变化
降香黄檀幼苗叶片气孔导度日变化曲线如图5所示。胁迫早期,在光合有效辐射达到最大值时(12:00),实验组降香黄檀幼苗叶片Gs明显低于空白组。且在光合有效辐射达到最大值前2 h,空白组Gs呈上升趋势,而实验组则呈下降趋势。胁迫中期,空白组和实验组的Gs总体上呈下降趋势。胁迫后期,两组的Gs在总体上呈平缓上升趋势。在多个时间点上,实验组的气孔导度都低于空白组,说明,锰胁迫会降低降香黄檀幼苗叶片的气孔导度。
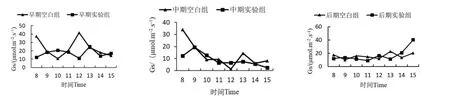
图5 降香黄檀幼苗气孔导度日变化曲线
2.2.5 降香黄檀幼苗叶片光能利用率(LUE)日变化
由图6可知,三个时期空白组和实验组的LUE日变化曲线存在明显差异。与空白组相比,实验组胁迫早期平均光能利用率降低了0.0282%,胁迫中期降低了0.02%,胁迫后期降低了0.0547%,说明锰胁迫会降低降香黄檀幼苗对光能的利用率。此外,三个时期的LUE日变化波动较大,这与当天的PAR变化有关。
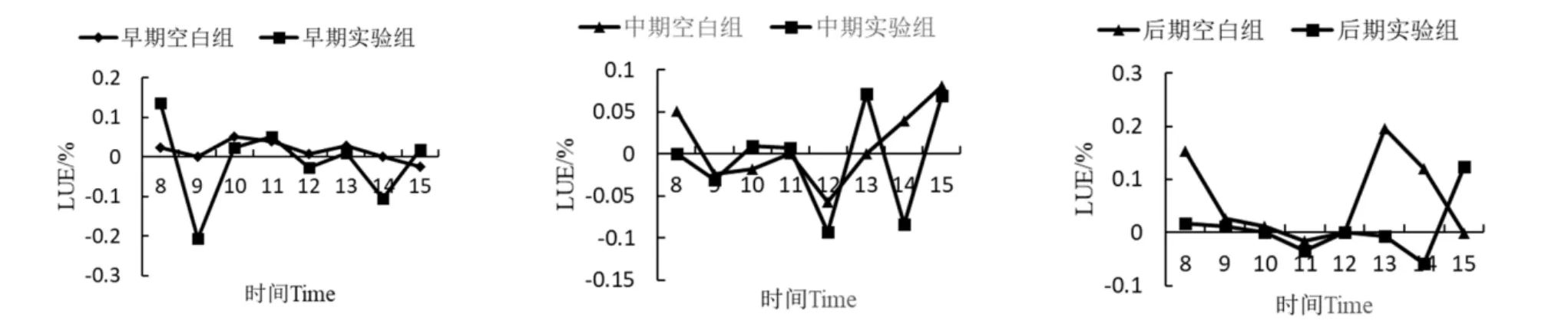
图6 降香黄檀幼苗光能利用率日变化曲线
2.2.6 降香黄檀幼苗叶片水分利用率(WUE)日变化
如图7所示,降香黄檀幼苗叶片的WUE日变化趋势与LUE相似。在PAR达到峰值时降香黄檀幼苗的WUE有明显的下降趋势。胁迫早期和中期,两组WUE变化规律均不明显。胁迫后期,实验组的日变化曲线呈“W”形,第一个低谷出现在11:00,第二个低谷出现在14:00。从日均值上看,与空白组相比,实验组胁迫早期WUE降低了0.04%,胁迫中期降低了0.15%,胁迫后期降低了0.85%。由此可以得出锰离子胁迫降低了降香黄檀幼苗叶片的水分利用率。
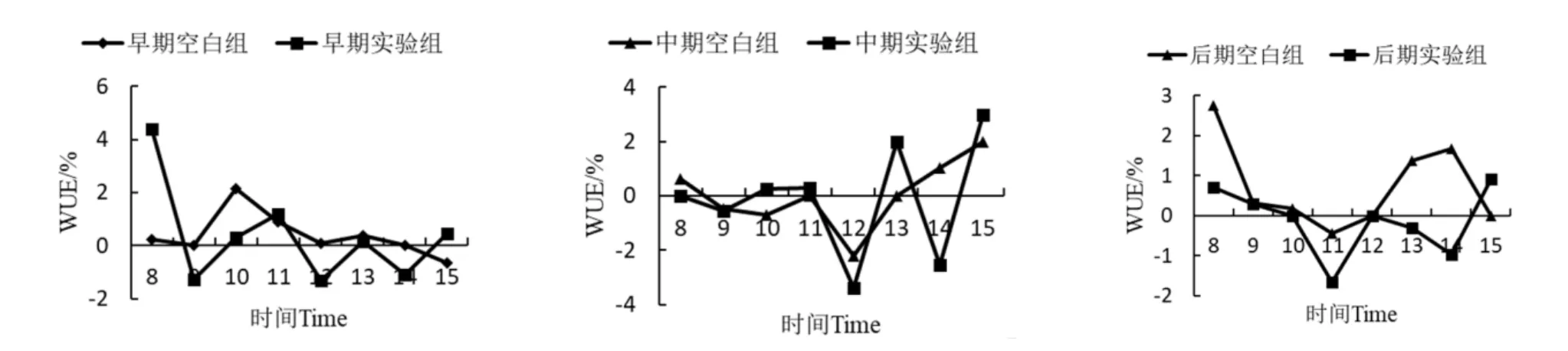
图7 降香黄檀幼苗水分利用率日变化曲线
3 讨论与结论
镁(Mg2+)是叶绿素的重要组成成分,但过量的锰离子会抑制镁离子的吸收和活性,导致叶绿素的合成受阻,致使植物的光合速率降低[11]。在锰离子的胁迫下,与空白组相比,降香黄檀幼苗叶片的净光合速率均值在胁迫早期降低了48.17%,而胁迫中期和后期实验组的Pn均值均为负值;蒸腾速率均值在胁迫早期降低了18.30%,中期降低了15.09%,后期降低了2.01%;气孔导度均值在胁迫早期降低了25.73%,中期降低了28.73%,后期则升高了6.35%;胞间CO2浓度均值在胁迫早期升高了13.78%,中期升高了8.22%,后期升高了5.48%。这说明随着锰胁迫的持续作用,降香黄檀幼苗的光合作用能力不断减弱。
光合作用为植物进行物质代谢和能量转换提供物质源,是植物赖以生存的基础,而净光合速率体现了植物有机物的积累[12-13]。引起净光合速率降低的因素有很多,其中主要有气孔限制因素和非气孔限制因素。气孔限制因素是由于植物气孔导度降低使得CO2进入叶肉细胞受到阻碍,从而引起光合速率的下降,非气孔因素则是因为植物叶肉细胞本身的光合活性受到了限制,进一步影响植物进行光合作用[14]。也就是说,当净光合速率下降时,细胞间隙CO2浓度和气孔导度同时下降,那么就说明引起植物净光合速率下降的主要因素是气孔限制因素;相反,如果净光合速率下降时,细胞间隙的CO2浓度升高而气孔导度却降低了,那就说明导致植物净光合速率降低的主要因素是非气孔限制因素。结合上文的数据分析可知,当用Mn含量为10 mg/kg的MnSO4进行胁迫处理时,在降香黄檀幼苗的净光合速率以及气孔导度降低时,细胞间隙的CO2浓度升高,因此可得,中度锰胁迫下引起降香黄檀幼苗光合作用速率降低的主要因素是非气孔限制因素。
猜你喜欢
林业与生态(2022年5期)2022-05-23
天然产物研究与开发(2018年5期)2018-06-13
新课程·下旬(2017年7期)2017-08-14
广东药科大学学报(2016年1期)2016-04-11
广西林业科学(2016年2期)2016-03-20
中国继续医学教育(2015年4期)2016-01-07
西北林学院学报(2015年5期)2015-01-03
家具与室内装饰(2013年8期)2013-02-20
植物营养与肥料学报(2011年2期)2011-10-26
植物营养与肥料学报(2010年3期)2010-11-16
