
中街山列岛海域曼氏无针乌贼栖息地偏向性研究
2022-12-12 11:41周永东刘姝含胡扬杰徐开达
浙江海洋大学学报(自然科学版) 2022年5期
吴 天,梁 君,周永东,刘姝含,胡扬杰,赵 瑞,徐开达
(浙江海洋大学海洋与渔业研究所,浙江省海洋水产研究所,浙江省海洋渔业资源可持续利用技术研究重点实验室,浙江舟山 316021)
曼氏无针乌贼Sepiella maindroni俗称墨鱼,是分布广泛的浅海种类,隶属软体动物门Mollusca、头足纲Cephalopoda、乌贼目Sepiida、乌贼科Sepiidae、无针乌贼属Sepiella,主要分布在日本海、马来群岛海域和印度洋东海岸,在我国东海和黄海均有分布,其中心分布区在浙江近海和闽东海域。历史上东海区曼氏无针乌贼最高年产超过7×104t,曾与大黄鱼Larimichthys crocea、小黄鱼Larimichthys polyactis、带鱼Trichiurus lepturus并称为我国四大海产[1-3]。但是20 世纪80 年代以来,由于过度捕捞和对资源的不合理利用,加之全球变暖造成栖息地退化,使得曼氏无针乌贼资源出现严重衰退现象,资源变动较大[4]。中街山列岛海域位于舟山渔场的中心区域,受沿岸水与外洋水的影响,水体交换快、饵料充足,是我国主要经济鱼类生长、索饵、繁殖的重要场所,历史上素有“鱼仓”之美誉,曾是曼氏无针乌贼的重要产卵场之一[5]。
栖息地是指生物个体和种群居住的场所,是由生物出现在环境中的空间范围和环境条件综合构成,栖息地的选择是指生物为了达到生存目的而寻找相对适宜栖息地的过程[6]。栖息地的研究一直以来都是动物生态学的研究重点,当栖息地质量下降时,会影响物种的数量,甚至导致物种灭绝[7]。在渔业资源研究中,栖息地的研究内容主要是生物栖息环境的变化对生物活动的影响,生物对栖息地的选择是受多种环境因子共同作用的结果,单因子分析并不能很好地解释生物对栖息地的选择机制,而广义可加模型(GAM)能够直接处理响应变量与解释变量之间的非线性关系,通过非参数的方法检验数据结构[8-9]。本研究基于2018-2021 年中街山列岛海域渔业调查数据和环境数据,通过GAM 构建非参数的拟合模型,探究中街山列岛海域曼氏无针乌贼资源分布与环境因子之间的关系,了解曼氏无针乌贼的栖息地选择机制,旨在为曼氏无针乌贼的种群保护及产卵场生物多样性保护提供理论基础,为浙江渔场修复振兴计划的实施提供科学依据。
1 材料与方法
1.1 研究区域和站位布设
于2018-2021 年春季(4 月)和秋季(10 月)在中街山列岛海域进行了8 个航次的底拖网渔业资源调查,调查范围如图1 所示。本研究中站位设置采取区域随机分布的原则,沿潮流主轴(西北-东南)共设置了16 个站位。
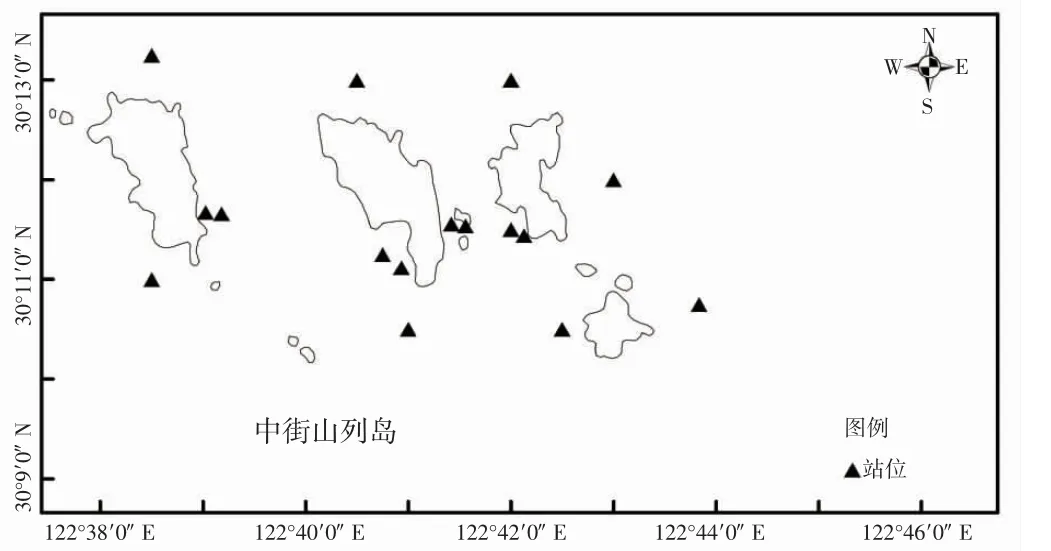
图1 调查站位示意图Fig.1 Sketch of survey station
1.2 采样方法和样品处理
调查船为58.8 kW 单拖渔船,拖速为2 kn,拖曳时间为30 min,调查网具网口周长25 m,囊网网目尺寸3.0 cm。在每一次调查时,使用温盐深测定仪(CTD)同步采集环境数据,并在各站位采集水样。共测定水深、表层海水水温、表层海水盐度、底层海水水温、底层海水盐度、叶绿素a、溶解氧、悬浮物、无机氮和活性磷酸盐10 类环境数据。样品采集、处理和分析等均按照《海洋渔业资源调查规范》[10]的规定执行。
1.3 数据处理和方法分析
1.3.1 资源密度和生物量估算
曼氏无针乌贼的资源密度估算和生物量估算按照以下公式[11]计算:

式中:D 为现存资源密度,ind.·km-2;C 为每小时取样面积内的渔获量,kg;q 为网具渔获率,由于曼氏无针乌贼属于沿岸暖水底栖种,q 取0.5;A 为网具每小时扫海面积,km2;B 为生物量,kg·km-2;为个体平均质量,kg·ind-2。
1.3.2 环境因子的相关性分析
在建模中,需要对环境因子进行相关性分析,克服共线性较高的解释变量对参数预估准确性的干扰,当2 个环境因子间存在共线关系时,保留1 个变量作为解释变量[12]。本研究采用Pearson 相关性分析对于环境因子之间可能存在共线性关系的环境因子进行相关性分析,并通过方差膨胀因子(variance inflation factors,VIF)对所有候选变量进行检验。
1.3.3 广义可加模型(GAM)分析
本研究以曼氏无针乌贼资源分布为响应变量,其中季节作为分类变量,各环境因子为解释变量。由于曼氏无针乌贼的零值站位出现的次数较多,假设曼氏无针乌贼分布模式为二项分布,GAM 模型选择logistic 为链接函数模拟中街山列岛海域曼氏无针乌贼资源分布同环境因子的关系[12-13]。GAM 的表达式为:

式中:P 为曼氏无针乌贼出现的概率;α 表示截距项;Si 表示样条平滑函数;xi表示第i 个解释变量;ε 表示误差项。
GAM 模型是在加性模型基础上提出的非参数拟合模型,能够直接处理响应变量与多个解释变量之间的非线性关系。在模型的拟合过程中,首先确定曼氏无针乌贼资源分布与单一环境因子的关系,通过逐步回归法将筛选后环境因子加入模型中,然后利用赤池信息准则(akaike information criterion,AIC)逐步检验加入不同环境因子后的模型拟合度,通过F 检验来评估各因子的显著性,以检验模型中每个环境因子的重要性,最终遴选出最优模型[13-14]。
数据通过Excel 软件计算和整理,曼氏无针乌贼资源分布图采用ArcGIS 10.2 软件绘制,Pearson 相关性分析和VIF 检验通过SPSS 23.0 实现,GAM 模型构建与检验及可视化分析通过R 4.2.0 实现。
2 结果
2.1 曼氏无针乌贼时空分布特征
曼氏无针乌贼资源空间分布季节变化明显。春季,曼氏无针乌贼主要分布于岛礁外侧,东北部居多(图2)。秋季,曼氏无针乌贼主要分布于岛礁内的东南和西南方向(图3)。时间因子月份对曼氏无针乌贼出现概率有明显影响,2018-2021 年曼氏无针乌贼的出现概率秋季明显高于春季。但春季曼氏无针乌贼的体质量高于秋季(图4)。

图2 2018-2021 春季曼氏无针乌贼尾数密度和生物量空间分布图Fig.2 2018-2021 spring distribution of S.maindroni tail density and biomass

图3 2018-2021 秋季曼氏无针乌贼尾数密度和生物量空间分布图Fig.3 2018-2021 autumn distribution of S.maindroni tail density and biomass
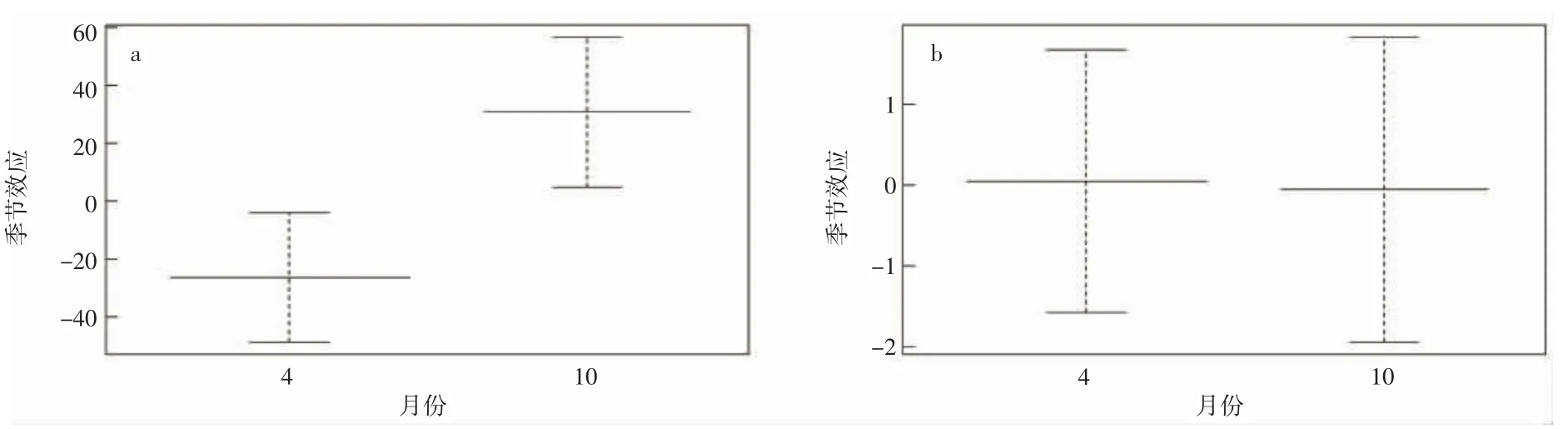
图4 2018-2021 曼氏无针乌贼尾数密度和生物量对照图(a.尾数密度;b.生物量)Fig.4 2018-2021 S.maindroni comparison of tail density and biomass(a.tail density;b.biomass)
2.2 关键环境因子识别
对于海洋生物,考虑的主要环境因子有温度、盐度、海表高度和叶绿素a等[5]。依据Pearson 相关性分析以及可视化结果(图5)显示,春季表层水温和底层水温、表层水温和底层盐度、底层水温和底层盐度、底层盐度和活性磷酸盐、叶绿素a和活性磷酸盐存在显著相关(P<0.05),秋季表层水温、表层盐度、底层水温、底层盐度之间均存在显著相关,无机氮和活性磷酸盐之间存在显著相关,同时对各解释变量进行多重线性检验,方差膨胀系数阈值设定为4,去掉相关性和方差膨胀系数均高于阈值的变量,因此本文选取水深、表层水温、表层盐度、悬浮物、叶绿素a、溶解氧和无机氮作为春季曼氏无针乌贼栖息地模型构建的解释变量;水深、悬浮物、底层水温、叶绿素a、溶解氧和活性磷酸盐作为秋季曼氏无针乌贼栖息地模型构建的解释变量。

图5 春季和秋季不同环境因子相关性分析热图(a 为春季;b 为秋季)Fig.5 Correlation analysis of different environmental factors in spring and autumn(a.spring;b.autumn)
2.3 GAM 模型构建
在GAM 模型中加入筛选后的环境因子,依据AIC 最小原则构建曼氏无针乌贼栖息地选择的最优模型。结果如下:
春季曼氏无针乌贼栖息地选择的最优模型:

秋季曼氏无针乌贼栖息地选择的最优模型:

式中:P为曼氏无针乌贼出现的概率;Depth 为水深,m;Chl-a为叶绿素a,μg·L-1;DO 为溶解氧,mg·L-1;ε 为相应的GAM 模型截距(表1)。
春季(GAM1)模型最佳预测变量组合为Depth+Chl-a(表1),模型累计偏差解释率为70.35%,水深和叶绿素a 对曼氏无针乌贼的出现概率具有显著影响(P<0.05),其贡献率分别为23.94%和46.41%(表2)。秋季(GAM2)模型最佳预测变量为DO(表1),模型累计偏差解释率为24.93%(表2)。
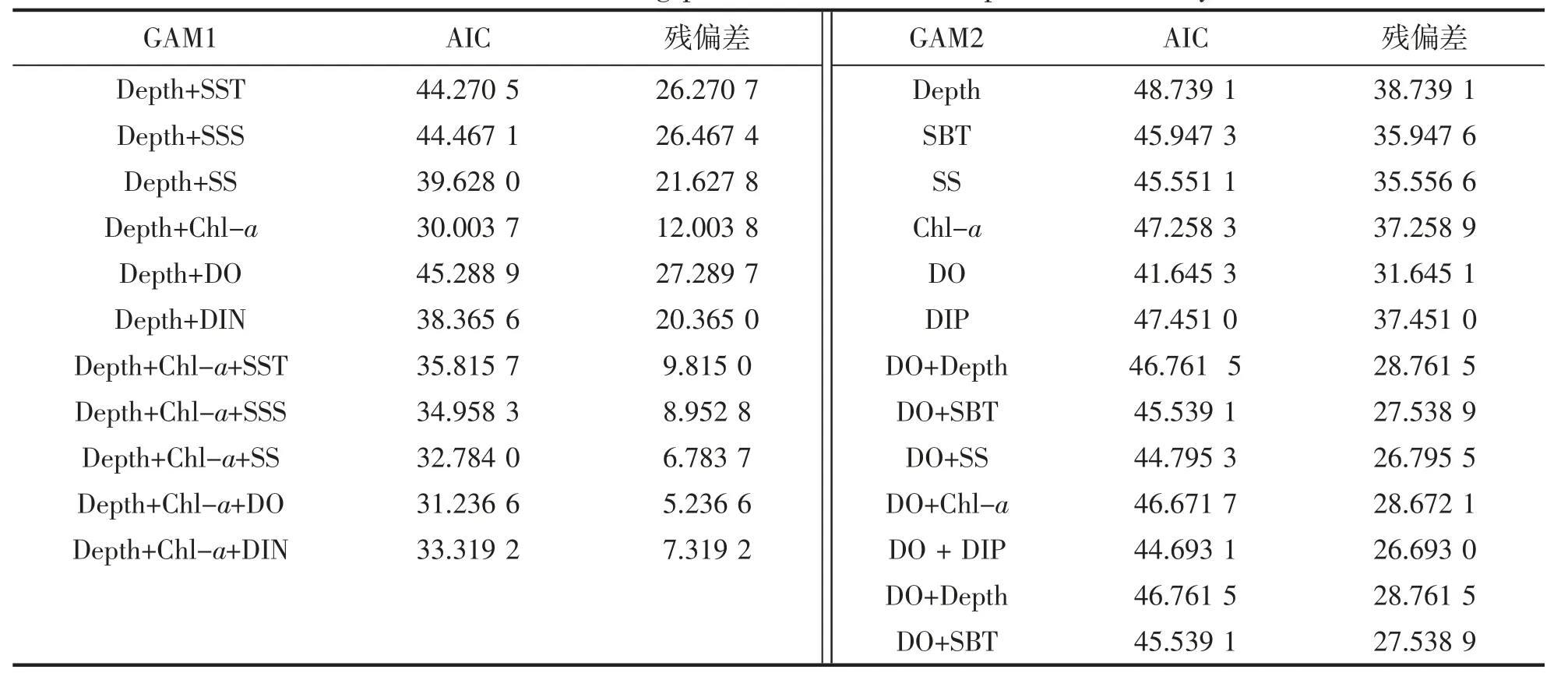
表1 GAM 模型变量筛选及影响因子的参数分析Tab.1 Variable screening process for GAM and parameters analysis

表2 模型拟合结果及各因子参数Tab.2 Result from models fitting and parameters of each factor
模型结果显示春季(GAM1)水深、叶绿素a 是影响曼氏无针乌贼栖息地选择的主要因素。春季曼氏无针乌贼主要出现的水深范围在25~35 m 之间,呈先上升后下降的趋势,30 m 时曼氏无针乌贼出现的概率最高(图6a)。叶绿素a 在0.5~1 μg·L-1时曼氏无针乌贼出现概率为上升趋势,当叶绿素a 超过1 μg·L-1时曼氏无针乌贼出现的概率开始下降(图6b)。秋季,影响曼氏无针乌贼出现概率的单一环境因子为溶解氧,范围在5.6~6.2 mg·L-1之间,呈先上升后下降的趋势(图6c)。

图6 GAM1 和GAM2 环境因子对曼氏无针乌贼分布的影响Fig.6 Effects of GAM1 and GAM2 of environmental factors on the S.maindroni
3 讨论
3.1 曼氏无针乌贼的时空分布变化
中街山列岛海域海洋性气候明显,春季在季风的影响下,东海表层水进入浙江近岸,与陆地径流(长江、钱塘江)交汇,形成锋区,渔场近岸水温较高,向东部水温逐步递减,近岸水与外侧水盐度差距较大。秋季季风转换,偏北风频率的增加,渔场内受冷暖水团交汇影响较强,由近岸往东水温逐步升高,盐度自西向东逐步增大[15]。曼氏无针乌贼属沿岸暖水底栖种,生命周期仅有1a,春季由外海低温高盐的越冬场游向近岸岛礁附近高温低盐的产卵场繁殖产卵,秋季随季风转换,开始向外海高盐海域移动,进行索饵、越冬洄游[16-19]。研究结果显示,春季(4 月)曼氏无针乌贼主要分布于岛礁外侧的东北部,秋季(10 月)曼氏无针乌贼主要分布于岛礁内部偏南方向,这与倪正雅等[17]和李成林等[20]的研究结果相一致。故不同季节曼氏无针乌贼的时空分布与海水温度、海水盐度和洄游路径有关。
本研究中,秋季曼氏无针乌贼的尾数密度明显大于春季,但春季的生物量却远高于秋季。春季为曼氏无针乌贼的繁殖期,主要为个体较大的曼氏无针乌贼产卵亲体,而秋季为曼氏无针乌贼的补充群体,故春季乌贼的体质量要高于秋季。而造成尾数密度秋季大于春季的原因可能与增殖放流和海洋保护区的资源养护作用有关。从2013 年开始,在中街山列岛海洋保护区内实施了大规模曼氏无针乌贼增殖放流,累计放流受精卵达7 835×104ind.,一定程度上增加了曼氏无针乌贼的补充群体[21]。中街山列岛国家级海洋特别保护区的设立有利于海洋生物的资源保护和修复,乌贼增殖礁的投放,也有利于自然海域曼氏无针乌贼受精卵的附着,提高了曼氏无针乌贼的附卵量,增加补充群体[22]。
3.2 栖息地选择的关键影响因子
不同生活史阶段对于生境的需求也有所不同,模型拟合结果显示春季曼氏无针乌贼的分布受水深和叶绿素a2 个关键因子的影响,秋季曼氏无针乌贼的分布受单一因子溶解氧的影响。
曼氏无针乌贼主要分布于近海浅水海域,其产卵场位于岛礁周围50 m 范围内,在繁殖期时从60~80 m的越冬场游向10~40 m 的产卵场作生殖洄游[3]。李星颉等[23]的研究表明,中街山列岛海域曼氏无针乌贼的附卵水深范围在10~20 m。本研究通过GAM 模型显示春季曼氏无针乌贼会选择水深范围在25~35 m 的海域作为产卵场和育幼场,在水深为30 m 时,曼氏无针乌贼的资源密度达到最高值,原因可能是本研究基于二项分布的GAM 模型来表达曼氏无针乌贼的出现概率,从已知数据预测整体分布特点,推断曼氏无针乌贼出现理论分布区,而前人研究以产卵附着基(柳珊瑚)上受精卵的附着率为调查内容,故本研究与前人研究结果略有差异。
叶绿素a的浓度值反映了海域初级生产者的现存量,能直接反映海区的肥瘪程度,是养育生物资源的直接指标,也能间接反映海区初级生产力的高低[24-26]。叶绿素a作为浮游植物丰富度的重要指标,能为浮游生物提供有利的生长条件[27]。叶绿素a浓度与头足类的渔场分布有一定关联性,如剑尖枪乌贼Uroteuthis edulis、神户乌贼Sepia kobiensis、金乌贼Sepia esculenta、长蛸Octopus variabilis等渔场分布均受到叶绿素a浓度值的影响[28]。根据GAM 模型分析的结果,叶绿素a对曼氏无针乌贼的分布影响较大,随着叶绿素a浓度的增加曼氏无针乌贼的资源密度也呈上升趋势,浮游生物的增多,可以为乌贼提供充足的饵料[29]。成体曼氏无针乌贼主要以小型鱼类、甲壳动物和少量软体动物为主,幼乌贼主要以浮游动物,如箭虫、桡足类等,特别是夜光藻可作为乌贼渔汛的直观指标[23,30]。
海水中充裕的溶解氧是维持海洋异养生物生命活动、避免水体有机物发生厌氧分解而散发臭味的必要条件,故海水中溶解氧被作为保障海洋生物生存的水化学指标和水体的感官指标[31]。李星颉等[23]对中街山列岛海域曼氏无针乌贼产卵区的生态环境进行调查,结果显示4-6 月自然海域曼氏无针乌贼产卵场的溶解氧为7.0~8.8 mg·L-1。蒋霞敏等[32]和刘振勇等[33]的研究表明曼氏无针乌贼室内孵化期的溶解氧为4.3~4.8 mg·L-1,在养殖过程中溶解氧应维持在5.0 mg·L-1以上。头足类的所有活动均需要消耗氧气,且曼氏无针乌贼的耗氧率与水温成正比例关系,个体体重成反比例关系,在低溶解氧的状态下,会造成曼氏无针乌贼生长缓慢,使其新陈代谢率降低,影响个体活动[34-35]。本研究结果显示,春季中街山列岛海域平均溶解氧为7.0 mg·L-1,这与李星颉等[23]的研究结果一致。模型拟合结果显示秋季曼氏无针乌贼偏向栖息于溶解氧浓度在5.5~6.5 mg·L-1之间的海域,同春季相比产卵区溶氧含量有所降低,蒋国昌等[36]的研究结果表明,东海区海水氧含量季节变化明显,春季含量最高,秋季含量最低。故季节变化同产卵区溶解氧浓度变化有明显关系。此外,近年来曼氏无针乌贼小型化明显,人工养殖的曼氏无针乌贼个体普遍偏小,生长速度快,寿命周期为4 个月左右,且人工养殖环境中溶氧可调控,故自然海域和养殖环境下曼氏无针乌贼所需溶氧有所不同[37]。
在构建曼氏无针乌贼栖息偏向性模型时,除水深、水温、盐度、叶绿素a、溶解氧等环境因子外,饵料生物与捕食者、天然附着基柳珊瑚与海藻、底质类型、流速等均可对曼氏无针乌贼栖息地的选择产生影响。因此,今后还需考虑生物与非生物因素共同影响下曼氏无针乌贼栖息地的选择,以期进一步提高模型的预测能力,全面客观地分析曼氏无针乌贼栖息地选择偏好与分布特征。
猜你喜欢
基层中医药(2021年8期)2021-11-02
科学与信息化(2020年11期)2020-06-19
计算机测量与控制(2017年6期)2017-07-01
水利科技与经济(2017年6期)2017-04-28
家教世界·创新阅读(2016年12期)2017-01-09
阅读与作文(小学高年级版)(2016年1期)2016-03-04
小朋友·快乐手工(2015年4期)2015-05-07
集美大学学报(自然科学版)(2015年1期)2015-02-28
海峡科学(2012年6期)2012-07-25
浙江海洋大学学报(自然科学版)(2011年5期)2011-07-12
