
中街山列岛海域鱼类群落特征及与环境因子的关系
2022-12-12 11:41:36周永东王忠明李振华
浙江海洋大学学报(自然科学版) 2022年5期
渠 遥,黄 波,周永东,王忠明,梁 君,李振华
(1.浙江海洋大学海洋与渔业研究所,浙江省海洋水产研究所,农村农业部重点渔场渔业资源科学观测试验站,浙江省海洋渔业资源可持续利用技术研究重点实验室,浙江舟山 316021;2.舟山市生态环境局岱山分局,浙江舟山 316200)
中街山列岛位于浙江北部海域,位于舟山群岛中部偏东,介于黄大洋和岱衢洋之间[1]。海域内岛礁众多,受到台湾暖流和黄海水团的影响,有丰富的饵料聚集,是东海主要经济鱼类的重要栖息地、生长繁殖场所和洄游通道[2]。中街山列岛所在的舟山渔场是我国最大的渔场,曾是我国渔业资源最丰富的渔场之一,被誉为“东海鱼仓”[3]。由于人类活动的干扰,超强度的酷渔滥捕,舟山渔场的生态环境和渔业资源发生令人担忧的变化[4]。为了保护海洋生态系统,促进海洋渔业资源的可持续利用,2006 年5 月,国家海洋局批准设立了中街山列岛国家级海洋特别保护区。鱼类是海洋生态系统的重要组成部分,人类活动的干扰和环境条件的变化对鱼类会产生一系列影响[5],维持鱼类的多样性是渔业可持续发展的重要支撑[6],因此研究分析中街山列岛海域鱼类群落特征及与环境因子的关系具有重要意义。为了进一步了解中街山列岛海域的鱼类资源与环境的现状,2021-2022 年在该海域进行了4 个航次的调查,对鱼类种类组成、优势种、群落多样性以及它们与环境因子的关系进行分析研究,以期丰富中街山列岛海域鱼类群落生态学内容,为恢复该地区的渔业资源和保护生物多样性提供参考,并为建立该地区的生态平衡提供理论支持。
1 材料与方法
1.1 数据来源
于2021 年4 月(春季)、7 月(夏季)、10 月(秋季)和2022 年1 月(冬季)在中街山列岛海域共设置18 个调查站位(图1),进行渔业资源和环境调查。
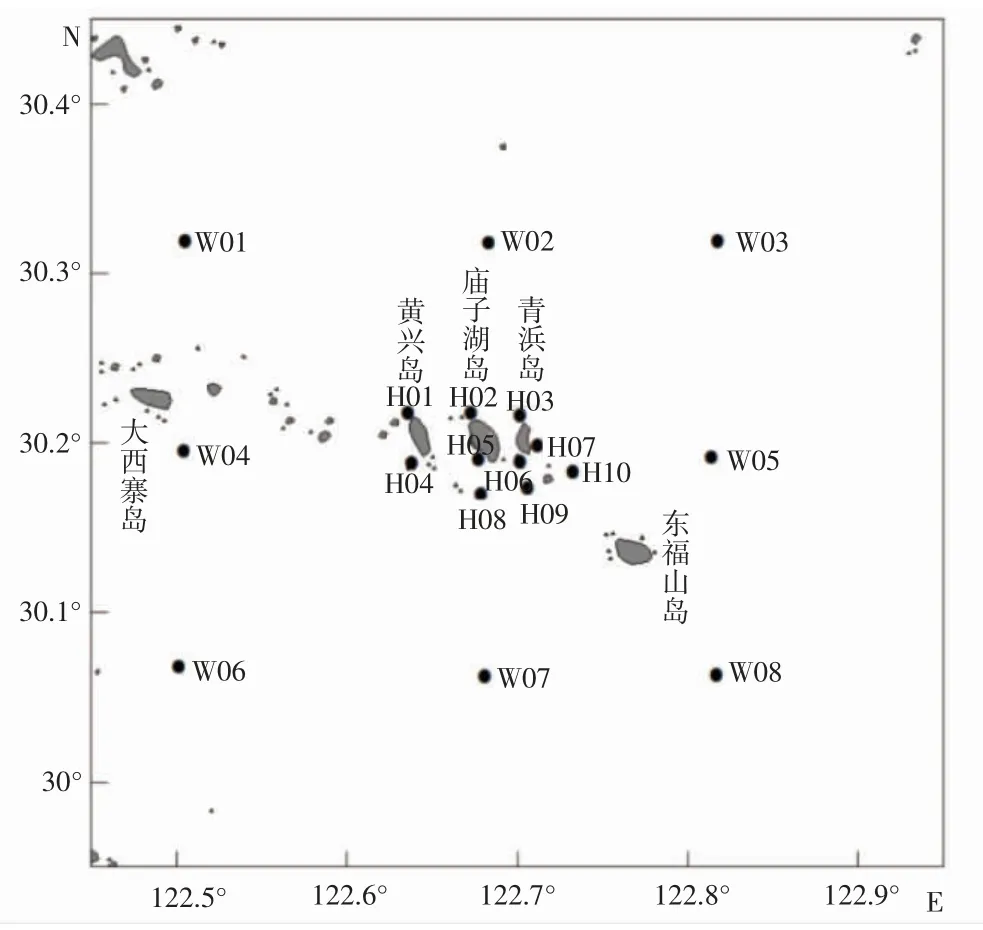
图1 2021-2022 年中街山列岛海域采样站位Fig.1 Sampling stations in the waters of Zhongjieshan Islands in 2021-2022
渔业资源调查采用单船拖网,调查船主机功率184 kW,船长26.2 m,底拖网网囊网目为2.5 cm,网口宽9.9 m。渔获物的采集、样品分析、生物学测定和数据分析处理按照海洋调查规范(GB/T 12763.6-2007)[7]和海洋渔业资源调查规范(SC/T 9403-2012)[8]的要求进行。每个站位拖网0.5 h,拖速3 kn。渔获物样品标记后进行低温冰鲜保存,带回实验室分析。鱼类尽量鉴定到种,对每一种鱼进行尾数和质量统计,质量精度为0.1 g。
环境因子包括2 部分,在每个站位使用多功能水质仪(AAQ171)测量水温、水深、盐度、溶解氧和pH 等环境因子数据;同时采集水样和泥样,测定的环境因子包括悬浮物(mg·L-1)、COD(mg·L-1)、无机氮(包括氨氮、亚硝酸盐和硝酸盐)、磷酸盐(mg·L-1)、叶绿素a(μg·L-1)和有机碳(%)。样品的采集和分析均按照《海洋调查规范第4 部分海水化学要素调查》(GB/T 12763.4-2007)[9]和《海洋沉积物质量》(GB 18668-2002)[10]标准执行。
1.2 数据处理
1.2.1 鱼类质量密度的估算
本次计算采用底拖网扫海面积法估算鱼类质量密度:

式中:D 为质量密度,kg·km-2;C 为平均每小时拖网渔获量,kg·h-1;a 为每小时网具扫海面积,km2·h-1;q 为逃逸率,取值范围为0~1,本研究取0.5[11]。
1.2.2 生态优势度
使用Pinkas 相对重要性指数(IRI)[12]衡量不同鱼类的生态优势度。

式中:P 为某一种渔获物的尾数占总尾数的百分比,%;W 为某一种渔获物的质量占总渔获质量的百分比,%;F 为某种渔获物出现的站位数占总站位数的百分比,%。本文以IRI≥1 000 的种类定为主要优势种,将500≤IRI<1 000 的种类定为优势种[13]。
1.2.3 多样性指数计算
采用Margalef 物种丰富度指数(D)[14]、Shannon 物种多样性指数(H′)[15]和Pielou 均匀度指数(J′)[16]分析群落的物种多样性。
(1)Margalef 物种丰富度指数:

式中:S 为渔获鱼类种类总数;N 为鱼类总尾数。
(2)Shannon 物种多样性指数:

式中:S 为鱼类种类总数;Pi为第i 种鱼类尾数占总尾数的比例。
(3)Pielou 均匀度指数:

式中:H′为鱼类Shannon 多样性指数;S 为鱼类总种类数。
1.2.4 数据处理
通过Excel 2010 对每个位点每网的鱼类种数(S)、个体数(N)、体重(W)等原始数据进行统计,计算Margalef 物种丰富度指数(D)、Shannon 多样性指数(H′)和Pielou 均匀度指数(J′)。
利用Primer 5.0 软件,以每个季节鱼类物种尾数密度为原始矩阵,进行log(x+1)转换后,运用等级聚类(Cluster)分析鱼类群落的空间结构。
利用Canoco 5.0 软件对2021-2022 年4 个季度鱼类调查数据和环境因子(水温、盐度、深度、溶解氧和pH 等)的关系进行分析。在RDA 排序图中,空心箭头代表环境因子,实心箭头代表物种,箭头之间的夹角代表物种与环境因子之间的相关性。分析前先对物种种类优势度进行筛选,排除出现3 次以下的物种,以减少机会种对数据分析结果的干扰[17]。
2 结果与分析
2.1 种类组成
通过4 个季度的调查,中街山列岛海域共鉴定鱼类70 种,隶属于17 目37 科58 属。以鲈形目种类数最多,共30 种(占种类总数的42.85%);其次为鲽形目和鲱形目,分别为8 种和7 种。春季共捕获鱼类40种,隶属于13 目22 科34 属;夏季共捕获鱼类33 种,隶属于9 目21 科29 属;秋季共捕获鱼类41 种,隶属于11 目20 科34 属;冬季共捕获鱼类42 种,隶属于12 目18 科33 属(表1)。
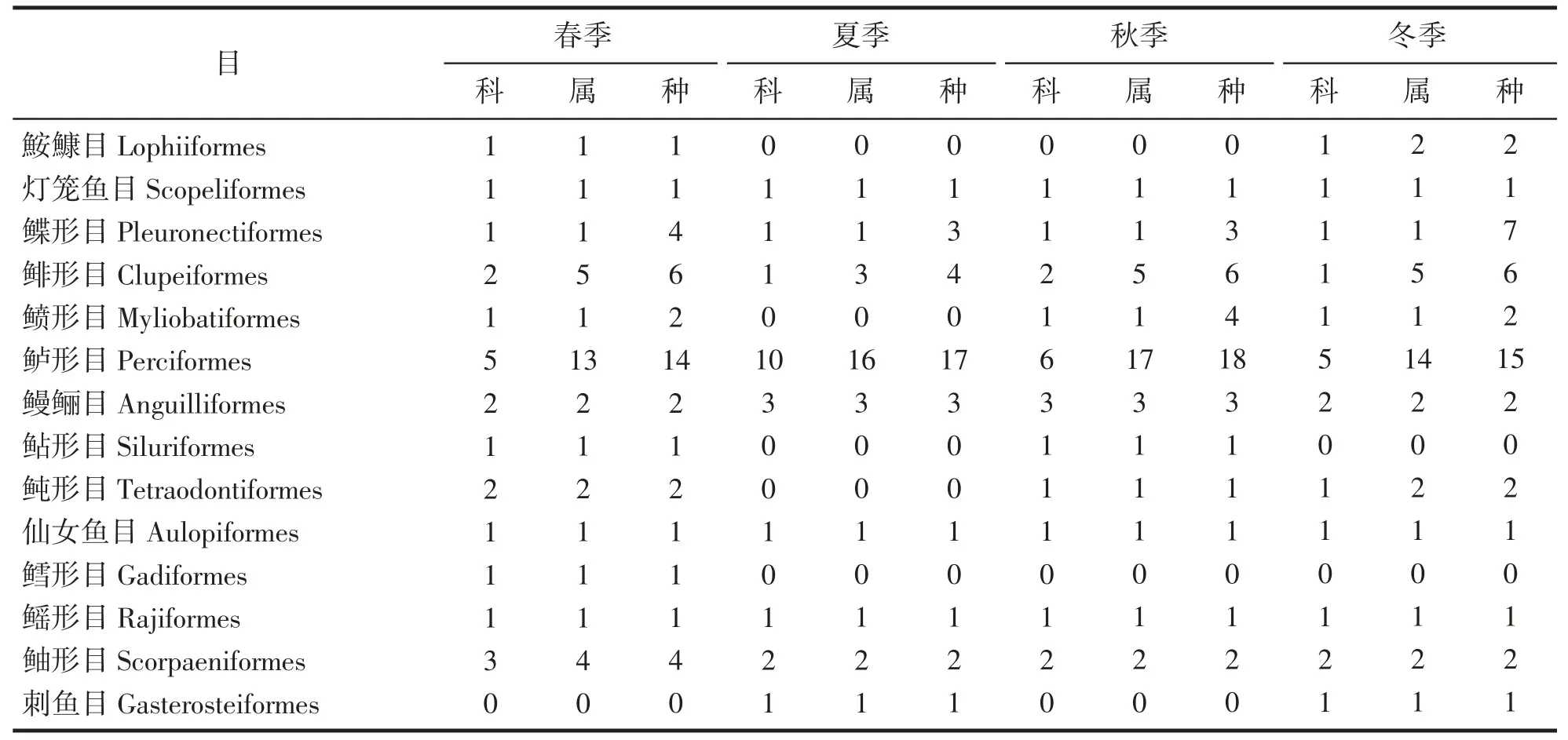
表1 中街山列岛海域2021-2022 年四季鱼类组成Tab.1 Species composition of fishes in the waters of Zhongjieshan Islands in 2021-2022
2.2 优势种
根据相对重要性指数的计算结果,中街山列岛海域鱼类的优势种共有10 种(表2)。春季优势种为龙头鱼Harpadon nehereus、凤鲚Coilia mystus和黄鲫Setipinna taty;夏季优势种为龙头鱼、凤鲚、小黄鱼Larimichthys polyactis、六丝钝尾虾虎鱼Amblychaeturichthys hexanema、带鱼Trichiurus japonicus、镰鲳Pampus echinogaster和拉氏狼牙虾虎鱼Odontamblyopus lacepedii;秋季优势种为龙头鱼、黄鲫、海鳗Muraenesox cinereus和刀鲚Coilia nasus;冬季优势种为龙头鱼、凤鲚、黄鲫和刀鲚。其中,龙头鱼为全年优势种。
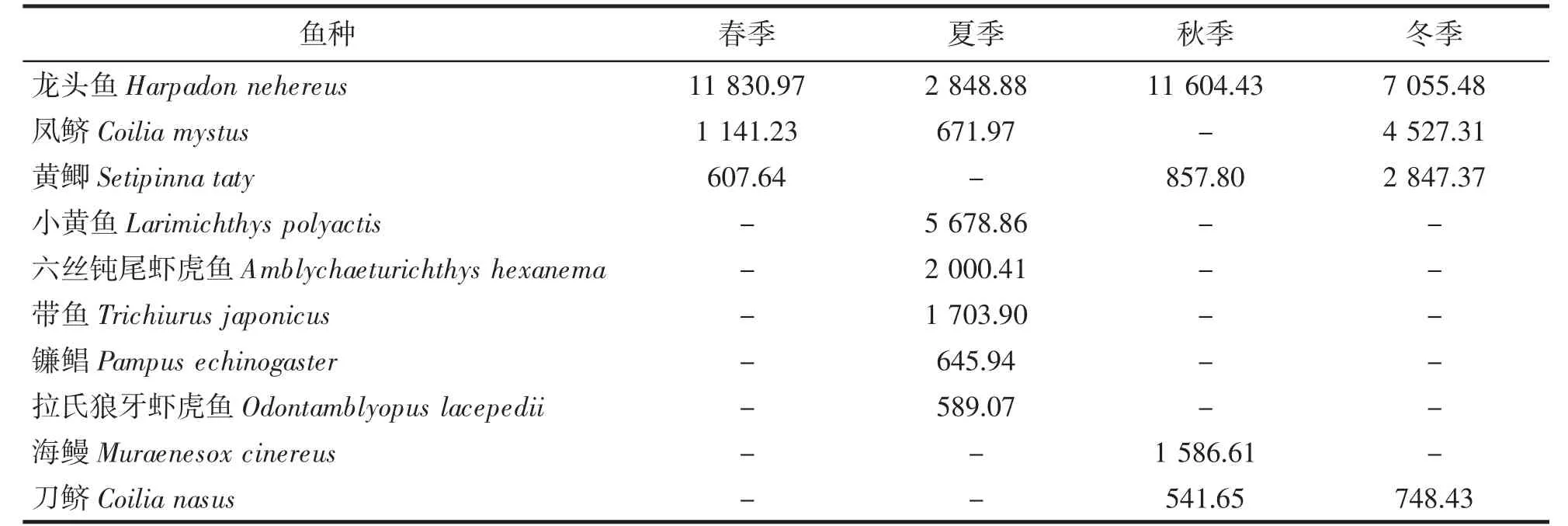
表2 中街山列岛海域2021-2022 年主要鱼类IRI 值的季节变化Tab.2 Seasonal variation of IRI values of main fishes in the waters of Zhongjieshan Islands in 2021-2022
2.3 渔获量分布
根据平均质量密度(kg·km-2)和数量密度(103ind.·km-2)分别研究调查海域鱼类相对资源量的季节变化。春季鱼类的平均质量密度为788.96 kg·km-2,平均数量密度为80.74×103ind.·km-2;夏季鱼类的平均质量密度为1 025.53 kg·km-2,平均数量密度为126.43×103ind.·km-2;秋季鱼类的平均质量密度为881.48 kg·km-2,平均数量密度为41.38×103ind.·km-2;冬季鱼类的平均质量密度为514.65 kg·km-2,平均数量密度为35.69×103ind.·km-2。个体平均质量(平均质量密度/数量密度)为秋季大于冬季大于春季大于夏季,平均数量密度最大值为最小值的3.54 倍(图2)。
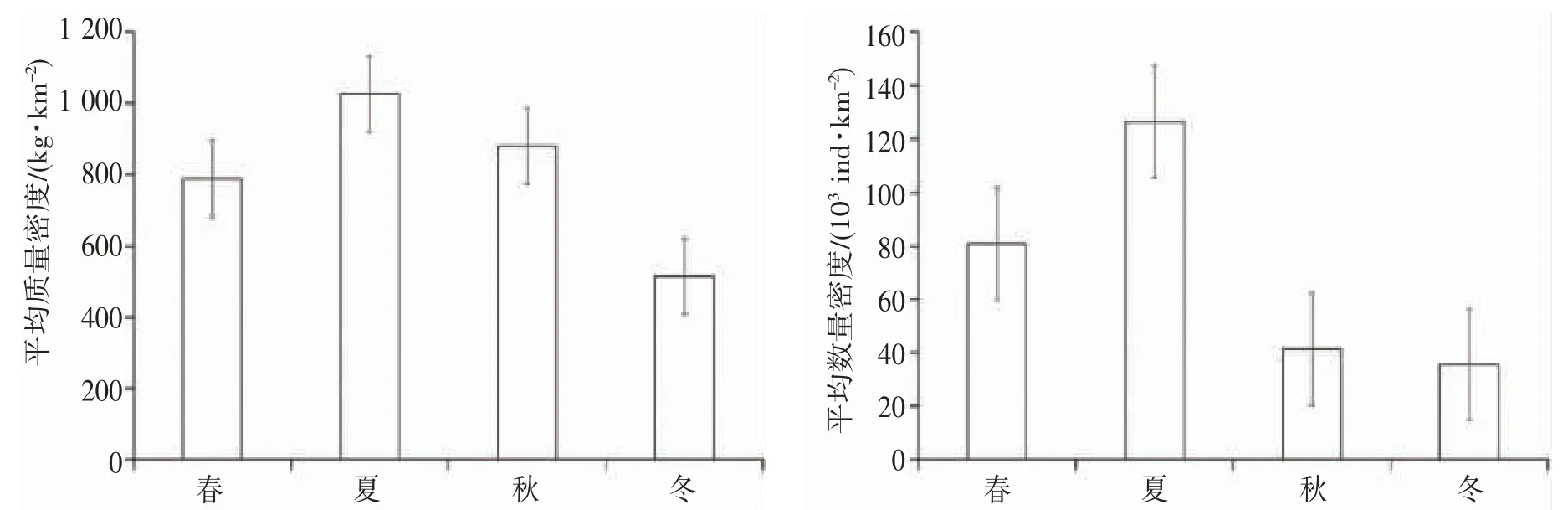
图2 中街山列岛海域2021-2022 年鱼类平均质量密度和数量密度的季节变化Fig.2 Seasonal variation of mean values of fish mass and density in the waters of Zhongjieshan Islands in 2021-2022
2.4 物种多样性
中街山列岛海域在2021 年4 月(春)、7 月(夏)、10 月(秋)和2022 年1 月(冬)鱼类的Margalef 物种丰富度指数(D)、Shannon 多样性指数(H′)和Pielou 均匀度指数(J′)的变化范围如图3 所示。物种丰富度指数D 变化范围为2.59~3.83,春季最高,夏季最低;Shannon 多样性指数H′变化范围为2.48~3.35;Pielou 均匀度指数(J′)变化范围为0.66~0.95。其中夏季的多样性指数H′和均匀度指数J′最高,冬季其次,秋季最低,最大值分别为最小值的1.35 倍和1.43 倍。Margalef 物种丰富度指数(D)春季最高,冬季其次,夏季最低。

图3 中街山列岛海域2021-2022 年鱼类多样性指数均值的季节变化Fig.3 Seasonal variations of mean values of fish diversity indexin the waters of Zhongjieshan Islands in 2021-2022
2.5 鱼类群落空间结构
等级聚类分析结果表明,中街山列岛海域鱼类群落呈现明显的空间结构(图4)。调查海域的鱼类群落在春季、夏季和冬季均出现3 组相似度较大的站位组群(图4a、4b 和4d),而在秋季出现4 组相似度较大的站位组群(图4c)。
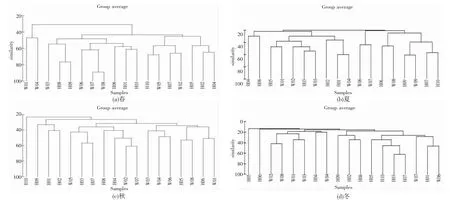
图4 中街山列岛海域2021-2022 年鱼类空间结构的聚类分析Fig.4 Hierarchical clustering analysis of spatial structure of the fish community in the waters of Zhongjieshan Islands in 2021-2022
2.6 鱼类群落时空分布与环境因子的关系
RDA 分析是直接梯度分析,它从分析结果中提取最能代表所有指标解释能力的最小的变量组合,而不是简单地分析一组变量或将几个变量组合成一个复合虚拟变量[18]。根据RDA 分析结果,得出春季时第一轴的特征值为0.277,第二轴的特征值为0.106 8,由此计算出物种与9 个主要环境因子累计方差为38.38%。实施蒙特卡洛显著性检验,得出第一排序轴和所有排序轴不显著,(第一轴:F=3.1,P=0.092;所有轴:F=1.5,P=0.07),而与叶绿素、水温、盐度和深度呈较强正相关(图5a)。
夏季时第一轴的特征值为0.288 9,第二轴的特征值为0.184 8,由此计算出物种与9 个主要环境因子累计方差为47.37%。同理得出第一排序轴和所有排序轴不显著(第一轴:F=3.3,P=0.154;所有轴:F=1.6,P=0.062),而与温度呈较强正相关(图5b)。
秋季时第一轴的特征值为0.165 1,第二轴的特征值为0.145 2,由此计算出物种与9 个主要环境因子累计方差为31.03%。同理得出第一排序轴和所有排序轴不显著(第一轴:F=1.6,P=0.852;所有轴:F=1.0,P=0.558),而与有机碳呈较强正相关(图5c)。
冬季时第一轴的特征值为0.341 3,第二轴的特征值为0.154 8,由此计算出物种与11 个主要环境因子累计方差为49.61%。同理得出第一排序轴和所有排序轴不显著(第一轴:F=3.1,P=0.314;所有轴:F=1.2,P=0.37),而与有机碳、悬浮物、pH 和磷酸盐呈较强正相关(图5d)。
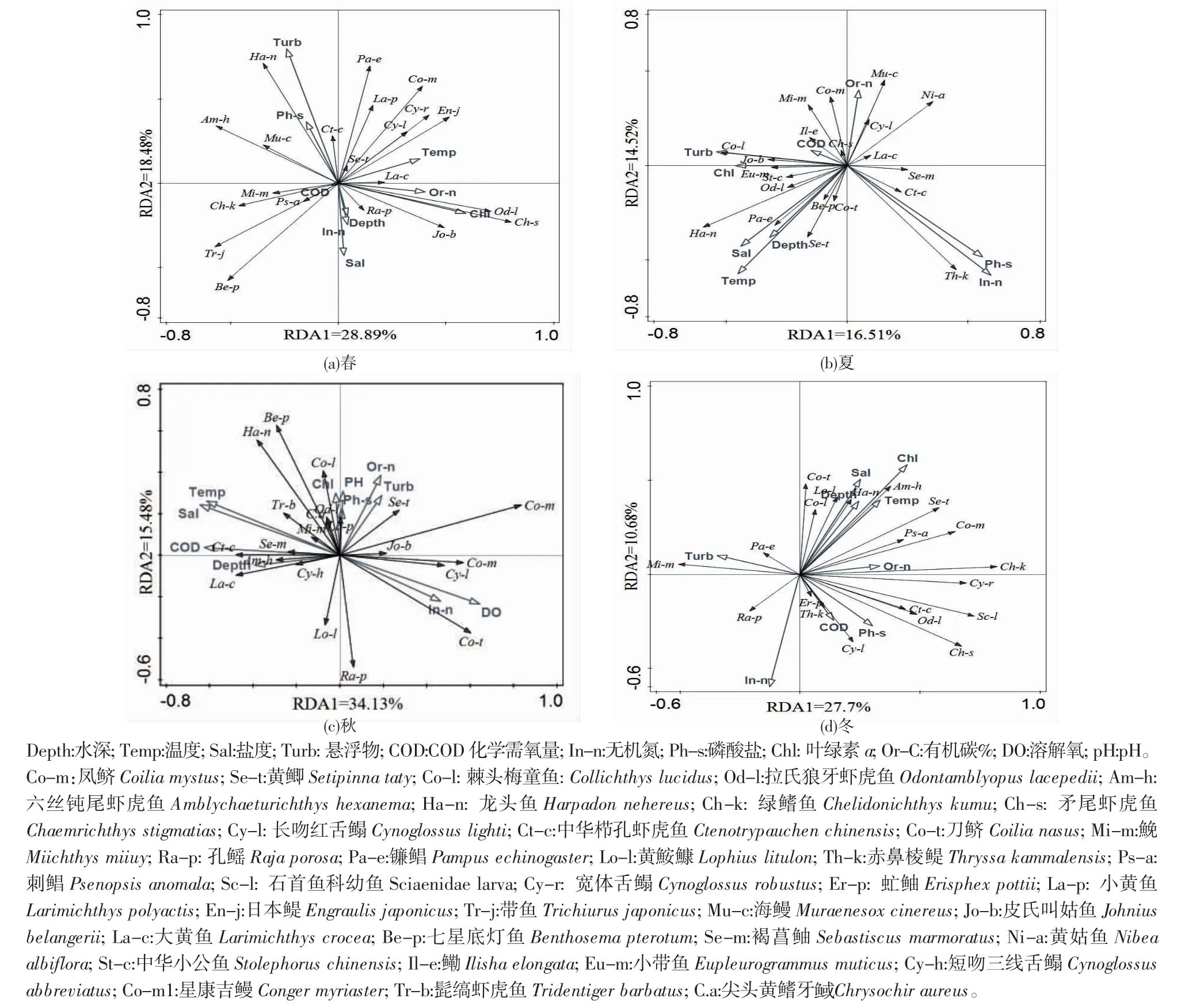
图5 中街山列岛海域鱼分布与环境因子的RDA 排序Fig.5 RDA ranking of fish community distribution and environmental factors in the waters of Zhongjieshan Islands
3 讨论
3.1 鱼类资源状况
3.1.1 种类组成的变化
中街山列岛海域鱼类种类丰富。梁君等[19]2010-2012 年在中街山列岛海域的调查结果显示,鱼类有55种,以鲈形目鱼类种类数最多,共计26 种,占所有种类的47.3%,本研究发现,在中街山列岛的底拖网捕获鱼类70 种,其中以鲈形目鱼类的种类数最多,共计30 种,占所有种类的42.85%。与10 年前的调查相比,鲈形目鱼类种类增多,但占比下降。种类数变动可能的原因有,一是调查站位数不同,本研究在调查海域共设置了18 个站位,10 年前梁君等[19]的调查仅设置了10 个站位。二是2 次调查的水文环境不同,拖网调查仅在4 个季度的代表月份各进行1 个航次的调查,结果存在一定的偶然性。三是鱼类种类数的增加可能与环境保护和资源养护有关。自2006 年在中街山列岛海域设置国家级特别保护区以来,中街山列岛的生物资源和环境的保护取得了显著的效果,因此鱼类种类的增加可能与此有关。
3.1.2 优势种的变化
优势种的变化在一定程度上反映了鱼类群落的组成和能量流动的变化[20]。本次调查结果表明,中街山列岛海域鱼类的优势种共有10 种。其中,龙头鱼在4 个季节均为优势种;凤鲚在春季和秋季2 个季节为优势种;黄鲫、小黄鱼、带鱼、六丝钝尾虾虎鱼和拉氏狼牙虾虎鱼仅在某一个季节为优势种。与其他鱼类相比,龙头鱼的优势度较为明显。根据2006-2007 年的调查结果,龙头鱼在舟山渔场是优势种之一,但不是主要优势种[21]。2010-2012 年中街山列岛海域的调查表明,龙头鱼已经成为3 个季节的优势种[19]。在邻近的马鞍列岛海域,2016-2017 年的数据表明,龙头鱼为全年优势种[22]。本次调查结果发现龙头鱼在中街山列岛海域的优势度明显上升,成为全年优势种鱼类。有研究表明,不可持续的渔业资源利用方式会导致群落的平均营养级水平下降,渔获对象有从长寿命、高营养级水平的物种向短寿命、低营养级水平的种类逐渐转变的趋势[23]。
中街山列岛海域夏季鱼类的优势种较为分散,可能是因为夏季是很多鱼类产卵季节,容易聚集在岛礁附近海域产卵[24],夏季受水温高的影响,鱼类资源较为丰富,所以优势种鱼类较多。
舟山渔场2007 年的优势种主要是小黄鱼、海鳗、绿鳍鱼、六丝钝尾虾虎鱼、星康吉鳗、黑鮟鱇Lophiomus setigerus 和日本红娘鱼Lepidotrigla japonice 等16 种[21],2010-2012 年中街山列岛海域的优势种有矛尾虾虎鱼、龙头鱼、长吻红舌鳎、小黄鱼、银鲳Pampus argenteus、黄鲫和凤鲚等共11 种。2016 年马鞍列岛海域鱼类的优势种共9 种,分别是龙头鱼、刺鲳、黑鳃梅童鱼Collichthys niveatus、宽体舌鳎、小黄鱼、带鱼、六丝钝尾虾虎鱼、凤鲚和棘头梅童鱼[22]。与这些调查结果相比,中街山列岛海域鱼类的优势种种类减少,优势度主要集中在龙头鱼等少数几种鱼类,可能是因为近些年过度捕捞导致渔业资源衰退和种群结构变化[21],也可能是因为作业方式不同和调查海域不同。
3.1.3 资源量的变化
2021 年中街山列岛海域生物量秋季最高,其次是夏季和春季,冬季最低。2021 年中街山列岛海域鱼类资源尾数密度夏季最高,其次是春季和秋季,冬季最低。秋季生物量最高,但尾数密度不高,表明秋季单个渔获的质量较大,经过伏季休渔鱼类经过数月的生长,个体质量有明显增加。夏季尾数密度最高,但生物量不高,表明鱼类处于幼鱼阶段,渔获数量多质量小。
中街山列岛海域2012 年鱼类的尾数密度指数春季最高为1 222×103ind.·km-2,2021 年春季尾数密度指数为1 442×103ind.·km-2,与2012 年相比,鱼类的尾数密度有所增加,表明资源量在一定程度上有所增长。
3.2 鱼类物种多样性
通过分析中街山列岛海域鱼类的物种多样性,鱼类物种多样性指数季节差异较大,调查海域多样性指数H′和均匀度指数J′夏季最高,冬季其次,秋季最低。Margalef 物种丰富度指数(D)春季最高,冬季其次,夏季最低。夏季物种多样性指数高,影响因素可能是夏季台湾暖流势力增强,中街山列岛海域夏季水温升高,大量鱼类洄游到该海域进行生殖产卵活动,因此,夏季多样性指数H′较高[25]。与胶州湾海域相比,中街山列岛海域的D 较高[26];与海南东部外海海域相比,中街山列岛海域D 较低[11]。
生物多样性指数越高,表明生物群落受到人类活动的干扰越小[27]。生物群落的H′和D 的值可分成(H′>4.0,D>4.0)、(3.0<H′<4.0,D>4.0)、(2.0<H′<3.0,2.5<D<4.0)、(1.0<H′<2.0,D<2.5)4 个等级[28]。中街山列岛海域鱼类的多样性指数H′为2.48~3.35,丰富度指数D 为2.59~3.83。中街山列岛海域鱼类群落处于良好状态,目前的海域环境和生态保护取得了一定的效果。
3.3 鱼类群落空间分布与环境因子的关系
通常鱼类群落空间分布与鱼类的栖息环境密切相关,栖息环境异质性高,鱼类多样性就高[29]。在不同的季节,中街山列岛海域鱼类的物种多样性指数、丰富度指数以及均匀度指数均存在差异,这表明生物环境因子是对中街山列岛鱼类群落影响的重要因素之一[30]。叶绿素、水温、盐度、深度、有机碳、悬浮物、pH 和磷酸盐等环境因子对中街山列岛海域鱼类群落分布有一定的影响。RDA 排序图表明,春季,环境因子对鱼类群落分布的累计贡献率高达61.46%;夏季,环境因子对鱼类群落分布的累计贡献率高达74.22%;秋季,环境因子对鱼类群落分布的累计贡献率高达59.96%;冬季,环境因子对鱼类群落分布的累计贡献率高达72.65%;春夏季是鱼类的主要产卵季节,环境因子对其影响较大,也是鱼类长期对适应自然环境的结果。本研究中,水温、叶绿素、有机氮、悬浮物是影响中街山列岛海域鱼类群落结构的主要环境因子。
猜你喜欢
海洋信息技术与应用(2022年1期)2022-06-05 07:38:24
中国宝玉石(2021年5期)2021-11-18 07:42:32
石油化工建设(2020年1期)2020-08-24 07:02:02
探索科学(2017年5期)2017-06-21 21:16:16
海洋信息技术与应用(2017年2期)2017-06-21 19:51:29
金色少年(奇趣科普)(2016年8期)2016-09-21 02:04:58
东南文化(2016年4期)2016-08-22 09:35:42
中学生天地(A版)(2015年7期)2015-08-06 05:39:07
小星星·阅读100分(低年级)(2014年5期)2014-06-24 00:42:45
读者(乡土人文版)(2013年10期)2013-04-12 07:16:43
