
桂西北喀斯特和非喀斯特森林土壤初级氮转化速率 及其影响因素
2022-12-07 13:29:32贺姣段鹏鹏李德军
农业现代化研究 2022年5期
贺姣 ,段鹏鹏 李德军 *
(1.中国科学院亚热带农业生态研究所亚热带农业生态过程重点实验室,湖南 长沙 410125;2.中国科学院环江喀斯特生态系统观测研究站,广西 环江 547100;3.中国科学院大学,北京 100049)
氮是构成一切生命体的关键元素,在陆地生态系统的演化和发展过程中发挥着至关重要的作用。植物在生长过程中吸收的氮主要来源于土壤[1]。除部分水溶性有机氮外,植物根系主要从土壤中吸收无机氮来保障自身生长与代谢对氮的需求[2-3]。土壤中的无机氮主要由微生物分解有机氮产生,随后被植物与土壤微生物吸收利用[2,4]。若土壤无机氮含量超过了植物与土壤微生物的氮需求量,过量的氮素尤其是硝态氮,会通过反硝化过程或随径流、淋溶等过程流失,从而加剧地下水硝酸盐污染、地表水体富营养化、温室效应与大气污染等一系列负面的环境问题[5-6]。因此,土壤氮含量及其动态变化一直是生态系统氮循环研究的热点。尽管测定不同形态的无机氮含量可以在一定程度上指示土壤氮素的供应水平和淋失风险,但不能揭示土壤无机氮具体的来源和消耗过程[7]。因此,为了揭示土壤氮素动态变化的机制,有必要深入研究土壤初级氮转化过程。
土壤无机氮产生与消耗涉及多个微生物主导的初级氮转化过程[4]。有机氮矿化是土壤无机氮产生的主要途径,一定程度上控制着自然生态系统土壤氮的供应潜力[8-9]。有机氮矿化产生的铵态氮(NH4+)在硝化细菌的作用下进一步氧化为硝态氮(NO3-)[4]。
硝化速率的大小决定了土壤无机氮的主要存在形 态[10]。在部分硝化速率高的土壤中,NO3-的积累导致较高的氮淋失风险[11]。从以往研究来看,土壤pH、有机碳(Soil Organic Carbon, SOC)、全氮(Total Nitrogen, TN)、C∶N比及土壤质地是影响土壤初级 氮转化速率的主要非生物因子,常常被用于解释不同 生态系统之间土壤初级氮转化特征的差异[8,10]。由于微生物是土壤氮转化过程的主要参与者,很多研究将土壤微生物生物量碳(Microbial Biomass Carbon, MBC)、微生物生物量氮(Microbial Biomass Nitrogen, MBN)及磷脂脂肪酸等作为影响氮转化的生物因 素[12-13]。一些研究表明,上述非生物因子结合土壤微生物指标可以显著提升土壤初级氮转化速率变异的解释度[8,10]。此外,土壤微生物对氮转化过程的驱动多依赖于特定功能基因的表达,一些研究也将氮转化功能基因丰度作为影响氮转化速率的生物因素加以考虑[14]。然而,多数研究仅考虑了少数几个生物或非生物因素,因此无法评判生物与非生物因素对土壤初级氮转化影响的相对重要性。
喀斯特生态系统是地球表层系统的重要组成部分,覆盖了全球地表面积的近15%[15],其中我国桂西北山区是喀斯特生态系统三大集中连片分布区之一[16]。与地带性土壤相比,喀斯特土壤具有偏碱性、富钙镁、SOC和TN含量较高的特征[17]。独特的土壤性质可能导致喀斯特土壤氮转化特征有别于同纬度的地带性红壤[17-19]。目前已有的两项研究结果均指出,喀斯特森林土壤硝化速率、微生物对NH4+同化速率及NO3-异化还原成铵(Dissimilatory Nitrate Reduction to Ammonium, DNRA)速率均高于邻近的非喀斯特森林[17-18]。我们前期发现喀斯特森林土壤的初级氮矿化速率是邻近非喀斯特森林的两倍,表现出较强的供氮能力[17]。但另一项研究却表明喀斯特森林土壤初级氮矿化速率显著低于临近的非喀斯特森林[18]。尽管上述二个研究均比较了喀斯特和非喀斯特森林土壤初级氮转化速率,但未考虑氮转化相关功能微生物对初级氮转化速率的可能影响,也未比较初级氮转化速率随土层的变化特征。
本研究测定了桂西北喀斯特森林及邻近的非喀斯特森林0~10 cm、10~20 cm及20~40 cm 3个土层初级氮矿化速率、初级硝化速率、自养硝化速率和异养硝化速率,并结合土壤理化因子和氮转化相关功能基因丰度,分析了二个森林氮转化速率的主控因素。主要研究目的包括:1)揭示不同氮转化过程在二个森林之间及不同土层之间的变化特征; 2)阐明二个森林自养和异养硝化对初级硝化速率的贡献;3)探讨氮转化功能基因丰度是否为氮转化速率的主控因素。
1 材料与方法
1.1 研究区概况
研究区位于广西壮族自治区环江县,地处亚热带季风气候区,年平均气温为17.8~22.2 ℃,1月份平均气温最低(7.8~13.0 ℃),7月份平均气温最高(25.8~29.4 ℃)。年平均降水量介于1 346~1 640 mm, 集中在4~8月。该地区多山丘,且交织分布着喀斯特和非喀斯特生态系统。喀斯特生态系统的基岩为石灰岩、白云岩及其不同比例混杂,而非喀斯特生态系统的基岩多为碎屑岩。本研究分别在木伦国家级自然保护区选择喀斯特森林(25°4' N,108°4' E)和在华山国家林场选择非喀斯特森林(25°10' N,108°18' E)。所选择的二个森林相距约26 km,均为20世纪50年代末被砍伐后自然发展起来的森林。喀斯特森林的优势树种包括伞花木(Eurycorymbus cavaleriei)、厚壳桂(Cryptocarya chinensis)和化香树(Platycarya strobilacea)等,非喀斯特森林的优势树种为椎木(Castanopsis Fagaceae)和鸭脚木(Schefflera octophylla)等。
1.2 实验设计
在每个森林分别建立一个面积为100 m × 100 m 的采样区,在每个采样区内随机选取3个10 m× 10 m 的样方。在每个样方随机选取二个采样点,去除凋落物层后分别采集0~10 cm、10~20 cm、20~40 cm 三个土层的矿质土壤样品,并将同一样方同一土层的样品混合均匀,组成一个混合样品。样品立即运回实验室,过2 mm网筛以去掉根系、石块和植物残体及充分混匀。每个样品分成三部分,其中一部分新鲜样品保存于4 ℃冰箱,用于测定氮转化速率、NH4+、NO3-、MBC和MBN;一部分新鲜样品于-40℃ 保存,用于测定氮转化相关的功能基因丰度;一部分样品风干后用于测定pH值、SOC、TN、交换性钙镁离子和土壤质地。
1.3 初级氮转化速率
采用15N同位素稀释法测定初级氮转化速率。将每个充分混匀的新鲜土壤样品分成五份,每份20 g。 将其中一份样品立即用2 mol/L氯化钾溶液(液土比为5∶1)浸提,测定处理前NH4+和NO3-含量。另外四份样品分成二组,每组两份。每一组的二份样品分别添加等量的15NH414NO3溶液或14NH415NO3溶液。将其中一组标记的样品立即用 2 mol/L氯化钾溶液浸提。另外一组标记的样品在25 ℃和黑暗条件下培养36 h,培养结束后用2 mol/L氯化钾溶液浸提。浸提液中的NH4+和NO3-含量利用流动注射分析仪(FIAstar 5000,FOSS,Sweden)分析。为了测定15N丰度,用化学方法分别将浸提液中的NH4+和NO3-转化为N2O,然后用带自动预浓缩装置的同位素比值质谱仪(MAT253 IRMS,Thermo Fisher,Germany)测定N2O中的15N丰度。
自养硝化速率为培养前后NO3-的15N丰度(原子百分超)的差异,乘以培养期间NO3-平均含量的差值,并使用NO3-的平均滞留时间进行校正,其中NO3-的平均滞留时间利用初始NO3-含量除以NO3-消耗速率计算得到;异养硝化速率为初级硝化速率和自养硝化速率之间的差值[20]。
1.4 DNA提取和实时定量PCR
参照Ouyang等[14]的方法测定土壤微生物转录丰度。称取0.25 g土壤样品利用MoBio PowerSoil DNA 分离试剂盒(MoBio Laboratories Inc.,Carlsbad,USA) 提取土壤DNA。根据制造商的方案,使用Quant-iT™ PicoGreen dsDNA BR检 测 试 剂 盒 对DNA提 取 物进行定量。使用SsoAdvanced SYBR Green Supermix和 CFXCONNECT PCR检测系统(Bio-Rad laboratories,Hercules,CA,USA)进行实时荧光定量PCR分析。 编码参与土壤氮转化相关酶的基因包括枯草杆菌蛋白酶(sub)、中性蛋白酶(npr)、几丁质酶(chiA)、脲酶(ureC)、氨氧化细菌氨单加氧酶(AOBamoA)和氨氧化古菌氨单加氧酶(AOAamoA)基因。其中chiA、npr、ureC、sub与初级氮矿化过程紧密相关,AOAamoA和AOBamoA与氨氧化过程相关。
1.5 土壤理化性质
利用pH计(FE20K, Mettler- Toledo, Switzerland) 测定土壤pH(土水比1∶2.5)。SOC采用重铬酸钾氧化-硫酸外加热法(油浴)测定;TN采用半微量开氏法-流动注射分析仪(FIAstar 5000,FOSS,Sweden)测定。交换性钙和镁离子含量经pH为7.0及浓度为1 mol/L的乙酸铵溶液浸提后使用安捷伦720电感耦合等离子体发射光谱仪(Agilent, Santa Clara, CA, USA)测定。利用激光粒度分析仪(Mastersizer 2000,Malvern,England)测定土壤质地,包括粘粒、砂粒和粉粒含量。MBC和MBN通过氯仿熏蒸提取法测定[21]。
1.6 数据统计分析
使用SPSS 16.0 软件(SPSS Inc., Chicago, IL, USA) 进行统计分析。首先对所有数据进行方差齐性和正态分布性检验,如果数据不是正态分布,将它们在分析之前进行对数转换。采用双因素方差分析检验森林类型、土层以及它们的交互作用对土壤氮转化速率、土壤生物和非生物因子的影响。采用独立样本t检验分析两个森林之间氮转化速率、土壤生物和非生物因子的差异是否显著。采用单因素方差分析检验同一森林三个土层之间氮转化速率、土壤生物和非生物因子的差异是否显著。采用皮尔森相关性分析评估氮转化速率与土壤生物及非生物因子之间的关系。采用多元线性回归分析确定氮转化速率的最强解释因子。
2 结果与分析
2.1 土壤生物和非生物因子
双因素方差分析结果表明,森林类型显著(P< 0.05)影响所有测定的生物与非生物因子;除pH值、交换性钙和镁、异养硝化速率、chiA和ureC之外的其他因子均在土层之间差异显著,其中C∶N比、sub和AOA在P< 0.1水平差异显著;森林类型和土层对NH4+、NO3-、初级硝化、自养硝化和AOB有显著(P< 0.05)交互作用,而对粉粒和npr在P< 0.1 水平上有显著交互作用(表1)。

表1 森林类型、土层及其交互作用对土壤因子和氮转化速率的影响Table 1 E§ects of forest type, soil horizon and their interaction on soil properties and gross nitrogen transformation rates
土壤pH值、TN、NO3-、交换性钙和镁、MBC、 MBN和粘粒含量均表现为喀斯特森林显著高于非喀斯特森林,NH4+、粉粒和砂粒含量则表现为非喀斯特森林显著高于喀斯特森林(表2)。二个森林的SOC、TN、NO3-、MBC和砂粒含量均随土层深度增加而降低,交换性钙和镁含量随土层深度增加无显著变化(表2)。对于喀斯特森林,C∶N比和粉粒含量随土层深度增加而降低,NH4+和粘粒含量随土层深度增加而增加,而pH值和MBN随土层深度增加无显著变化。对于非喀斯特森林,土壤pH和粘粒含量随土层深度增加而增加,NH4+和MBN含量随土层深度增加而显著降低,而C∶N和粉粒含量随土层深度增加无显著变化(表2)。

表2 喀斯特和非喀斯特森林不同土层理化性质Table 2 Soil physicochemical properties in the three soil horizons for the karst and non-karst forests
土壤chiA、ureC、sub和AOA丰度均表现为喀斯特森林显著高于非喀斯特森林。对于喀斯特森林,除npr丰度随土层深度增加而显著降低外,其他功能基因丰度沿土层无显著变化(表3)。对于非喀斯特森林,AOB丰度在0~10 cm土层最高,而在10~20 cm土层最低,其他功能基因丰度随土层深度增加无显著变化(表3)。

表3 喀斯特和非喀斯特森林不同土层氮转化相关功能基因丰度(log(基因拷贝数/g干土))Table 3 Soil nitrogen transformation related gene abundances in the three soil horizons for the karst and non-karst forests (log (copies/g dry soil))
2.2 初级氮转化速率
双因素方差分析结果表明,森林类型显著(P< 0.05)影响所有测定的氮转化过程速率;除异养硝化速率外的其他过程速率均在土层之间差异显著;森林类型和土层对初级硝化和自养硝化速率有显著(P< 0.05)交互作用(表1)。土壤初级氮矿化速率、初级硝化速率、自养硝化速率和异养硝化速率均表现为喀斯特森林显著高于非喀斯特森林(图1)。对于喀斯特森林,土壤初级氮矿化速率、初级硝化速率和自养硝化速率均随土层深度增加而降低,异养硝化速率在土层间没有显著差异(图2)。对于非喀斯特森林,只有初级氮矿化速率随土层深度增加而显著降低(图2)。在0~10 cm和10~20 cm二个土层,初级硝化速率和自养硝化速率均是喀斯特森林显著高于非喀斯特森林,而在20~40 cm土层,上述二个过程速率无显著差异(图2)。森林类型显著(P< 0.05)影响自养硝化对初级硝化速率的贡献,在0~10 cm和10~20 cm二个土层均是喀斯特森林显著高于非喀斯特森林,而在20~40 cm土层二个森林之间无显著差异(图3);森林类型和土层对自养硝化的贡献有显著(P< 0.05)交互作用 (表1)。喀斯特森林土壤自养硝化对初级硝化速率的贡献平均为75.04%,且随土层深度增加呈下降的趋势;非喀斯特森林土壤自养硝化对初级硝化速率的贡献平均为28.51%,且20~40 cm土层显著高于0~10 cm和10~20 cm土层。
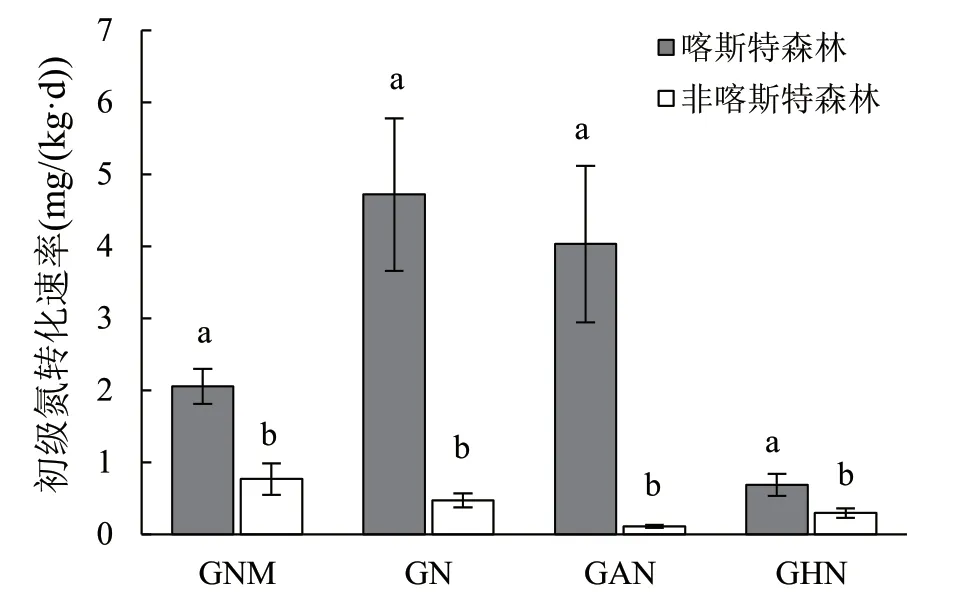
图1 喀斯特和非喀斯特森林土壤初级氮矿化和硝化速率 Fig.1 Gross N mineralization rate and gross nitrification rates in the karst forest and non-karst forest

图2 喀斯特和非喀斯特森林三个土层初级氮矿化和硝化速率Fig.2 Gross N mineralization rate and gross nitrification rates for three soil depths in the karst forest and non-karst forest

图3 自养硝化速率对初级硝化速率的贡献Fig.3 The contribution of autotrophic nitrification to gross nitrification rate
2.3 初级氮转化速率与土壤生物和非生物因子的相互关系
相关分析表明,对于喀斯特森林,初级氮矿化速率与SOC、T N、NO3-、MBC含量显著正相关,并与npr基因丰度显著正相关。初级硝化速率和自养硝化速率与土壤非生物因子和生物因子的关系表现出一致性,两者均与SOC、TN、NO3-、MBC和MBN含量正相关,与NH4+含量显著负相关,并与AOA和AOB基因丰度显著正相关(表4)。对于非喀斯特森林,初级氮矿化速率与SOC、TN、NH4+、NO3-和MBN含量正相关。自养硝化速率和MBC正相关。多元线性逐步回归分析结果表明,对于喀斯特森林,初级氮矿化速率、初级硝化速率和自养硝化速率的最强解释因子均为MBC,分别解释了氮转化速率变异的62%、84%和86%(表5)。对于非喀斯特森林,初级氮矿化的最强解释因子为TN,解释了速率变异的77%;自养硝化的最强解释 因子为AOA丰度,解释了速率变异的56%(表5)。

表4 土壤氮转化速率与生物和非生物因子之间的相关性Table 4 Correlation coeきcients between soil properties and gross N transformation rates
3 讨论
3.1 喀斯特与非喀斯特森林之间氮转化速率的差异
目前仅有两项研究比较了亚热带喀斯特森林和非喀斯特森林的初级氮转化速率[17-18]。其中一项研究显示喀斯特森林初级氮矿化和异养硝化速率显著低于非喀斯特森林,与本研究结果相反,可能是由于所选森林的代表性有关。在上述研究中,SOC和TN的含量在喀斯特和非喀斯森林之间均没有显著差异,但在本研究中喀斯特森林的SOC和TN含量显著高于非喀斯特森林[18]。在区域尺度上的研究也表明,喀斯特森林SOC和TN含量通常高于相邻的非喀斯特森林[17,22-24],因此,本研究所选择的喀斯特和非喀斯特森林可能更具有代表性。前述研究也表明自养硝化速率和初级硝化速率均是喀斯特森林显著高于非喀斯特森林[18],与本研究结果一致。我们前期也比较了喀斯特森林和非喀斯特森林土壤初级氮矿化和硝化速率[17],结果与本研究一致。
3.2 生物和非生物因子对喀斯特与非喀斯特森林之间氮转化速率的影响
基于全球数据整合分析表明,初级氮矿化速率与SOC、TN和土壤微生物量呈显著正相关关系,其中TN既是氮矿化作用的底物,也能通过增加微生物量促进氮矿化速率[8]。在本研究中,喀斯特森林土壤TN、MBC和MBN均显著高于非喀斯特森林,因此,上述机理能很好地解释喀斯特森林土壤初级氮矿化速率显著高于非喀斯特森林。另一项基于全球整合分析的研究表明,土壤pH、微生物量和TN与初级硝化速率呈显著正相关关系;自养硝化速率主要由C∶N和pH驱动,而异养硝化速率主要受TN控制[10]。NH4+是自养硝化的底物,因此很多研究报道自养硝化速率随NH4+含量增加而增 加[5,10]。在本研究中,喀斯特森林土壤NH4+含量显著低于非喀斯特森林,但前者的自养硝化速率却较高,意味着其他因素的作用超过了NH4+。有研究表明,当土壤pH值较低时,自养硝化往往受到抑制,因此异养硝化是酸性土壤硝化作用的主要形式[25]。本研究中,喀斯特森林土壤pH值显著高于非喀斯特森林,因此,较高的土壤pH水平是喀斯特森林土壤自养硝化速率较高的重要原因。此外,对于自然生态系统,土壤NH4+主要来源于初级矿化过程,因而TN能通过促进初级矿化而增加自养硝化速 率[5,10]。因此,喀斯特森林土壤较高的TN水平也能部分解释其较高的自养硝化速率。由于TN是异养硝化过程的底物,因此,喀斯特森林土壤较高的TN水平也是导致其较高的异养硝化速率的主要原因,这与全球整合分析结果一致[10]。由于喀斯特森林土壤更有利于自养硝化过程,导致硝化速率以自养硝化为主,而非喀斯特森林则以异养硝化占主导。
由于npr基因编码胞外蛋白酶,npr的丰度能一定程度上反映土壤蛋白酶的活性[26-27]。蛋白质是土壤有机质的重要组成部分,也是土壤有机氮的主要储存库[28]。微生物通过蛋白酶解聚蛋白质是土壤氮矿化过程的限速步骤,这可能是npr基因丰度对初级氮矿化有显著影响的重要原因。本研究结果显示,喀斯特森林和非喀斯特森土壤npr的丰度与初级氮矿化速率显著正相关,这与之前的研究一致[14]。以往研究表明,AOA对自然生态系统土壤自养硝化过程的作用高于AOB[29]。本研究中,我们发现喀斯特森林的AOAamoA基因丰度显著高于非喀斯特森林,因此该基因丰度的差异可能是导致前者自养硝化速率较高的重要原因。然而本研究只对AOA和AOB的基因丰度进行了研究,对于AOA和AOB对自养硝化过程的相对贡献有待进一步的探讨。
3.3 喀斯特与非喀斯特森林内部氮转化速率变异的主控因素
微生物量不仅是表征微生物群落丰度和活性的指数,而且被认为是土壤活性有机碳氮的来源[30-31]。多项研究指出,微生物量可以通过调控土壤初级氮矿化进而对硝化速率产生影响,因为初级氮矿化速率在一定程度上决定了NH4+的可利用水平[5]。此外,MBC通常与TN显著正相关,如本研究中二者的相关系数达0.93(P< 0.001)。如前所述,TN是初级氮矿化的底物,并进一步影响初级硝化速率。因此,MBC对喀斯特森林初级氮矿化与硝化速率的影响既有直接作用,也间接反映了TN对氮转化过程的影响。也正因如此,TN是非喀斯特森林初级氮矿化速率的最强解释因子。pH是影响AOA和AOB组成分布的主要驱动因子之一[32],且AOA相比于AOB更能适应酸性的土壤环境[29],因此,在酸性土壤中AOA对初级硝化速率的贡献远远高于AOB[33-34]。与此一致,我们发现AOA基因丰度是非喀斯特森林土壤自养硝化的主控因素。可见,虽然与氮转化相关的功能基因丰度与其相应的氮转化速率具有显著相关性,但其作用低于土壤理化性质。其他研究也得到了类似的结论[14,35-36]。
4 结论
本研究结果表明,土壤初级氮转化速率在不同森林及土层之间存在显著差异。
1)喀斯特森林土壤初级氮矿化速率和硝化速率显著高于非喀斯特森林。其中,喀斯特森林土壤硝化作用多为自养硝化作用,而非喀斯特森林土壤硝化作用则以异养硝化作用为主导。
2)在喀斯特森林土壤中,初级氮矿化和硝化速率均随土层深度增加而减少。在非喀斯特森林土壤中,初级氮矿化速率随土层深度增加而下降,初级硝化速率在不同土层间的差异并不显著。
3)喀斯特森林初级氮矿化、初级硝化和自养硝化速率主要受MBC影响,而非喀斯特森林氮矿化和自养硝化速率更多取决于TN和AOAamoA基因丰度。
综上所述,虽然与氮转化相关的功能基因丰度与其相应的氮转化速率具有显著相关性,但其作用低于土壤理化性质。该研究结果将有助于深入认识亚热带典型喀斯特和非喀斯特森林土壤氮循环特征及氮累积机制,为岩溶区植被恢复工程的深入实施提供科学支撑。
猜你喜欢
北京大学学报(自然科学版)(2022年4期)2022-08-18 06:57:34
草业学报(2021年9期)2021-09-22 08:55:16
阅读(低年级)(2020年8期)2020-11-06 06:23:19
乡村地理(2019年2期)2019-11-16 08:49:32
————水溶蚀岩石的奇观
家教世界(2019年4期)2019-02-26 13:44:20
农业环境科学学报(2017年2期)2017-03-20 14:57:30
亚热带资源与环境学报(2015年4期)2015-01-22 07:06:16
文化月刊·下旬刊(2014年6期)2014-08-28 00:58:59
东北师大学报(自然科学版)(2014年1期)2014-02-27 08:02:15
亚热带资源与环境学报(2014年3期)2014-01-23 01:52:46
