
氮添加对沂蒙山区麻栎和黑松林土壤碳固存的影响
2022-12-07 09:43:00李利萍张义群田会梅
农村科学实验 2022年24期
杨 柳 李利萍 张义群 丁 彬 刘 琳 田会梅
(1.山东省林业保护和发展服务中心,山东 济南 250014;2.山东省林草种质资源中心(山东省药乡林场),山东 泰安 271043;3.黄河下游森林培育国家林业和草原局重点实验室/山东泰山森林生态系统国家定位观测研究站,山东泰安 271018)
随着工业革命和经济的持续发展,由于化石燃料的燃烧和农业中化肥的使用,大气中的氮排放量急剧增加。中国现在正在成为世界三大氮沉降区之一,并且预计氮沉降量将继续增加。我国年平均氮沉降量为12.9 kg hm-2yr-1,而山东省的氮沉降量为23.6 kg hm-2yr-1,远高于我国平均氮沉降量(陈红等,2019)。并且沂蒙山区是受到氮限制比较强烈的地区,其生态系统对氮的增加特别敏感。
氮沉降通过增加土壤氮素有效性,从而影响有机质的分解。这对生态系统的土壤碳循环和碳固存具有重要意义。研究表明,氮沉降可显著减少土壤微生物的生物量和丰度,减缓土壤呼吸,降低土壤酶活性,从而增加土壤对碳的吸收(汲常萍等,2014)。
土壤碳汇对氮沉降的响应主要受土壤微生物调控。氮沉降会通过改变土壤理化生物性质(如氮的有效性、土壤酸化、碱基阳离子组成等)直接影响土壤微生物多样性,还可以通过陆地植被的生理和生态反应间接影响土壤微生物(樊后保等,2008)。研究表明,土壤微生物通过多种胞外酶,参与元素的生物化学循环,影响土壤碳的周转(周正虎等,2016)。研究表明,在森林生态系统中,氮沉降能够明显提高β-1,4-N-acetyl-glucosaminidase(βG)的活力。这会加速土壤碳的周转,并影响土壤碳固存(鲁显楷等,2019)。因此本研究通过探究土壤碳汇对模拟氮沉降的响应,以期为预测未来碳汇变化提供理论依据。
1.材料与方法
1.1 研究区概况
试验地于沂蒙山区国有大洼林场(35°32′46″N,117°54′47″E)。当地属暖温带季风气候,年均气温13.2 ℃,年均降雨量750 mm,无霜期约196天。母岩多为花岗片麻岩和石灰岩,土壤以棕壤土和褐土为主,pH 5.0~7.0。当地主要为麻栎林( Quercus acutissima Carruth.)和黑松林( Pinus thunbergii Parl.)。林下灌木主要有荆条(Vitex negundo),草本覆盖主要有京芒草(Achnatherum pekinense)和沿阶草(Ophiopogon bodinieri)等。样地概况如下(表1)。

表1 麻栎和黑松样地概况
1.2 试验设计
于2020年10月21日在大洼林场选择坡向、坡度相似且相对平坦、林分密度相对均匀的两种具有代表性的麻栎林和黑松林作为研究对象。根据当地大气氮沉降背景值利用倍增法进行模拟氮沉降。共设置4个氮添加处理,分别为:对照(0 kg N hm-2yr-1,CK),低氮添加(50 kg N hm-2yr-1,LN),中氮添加(100 kg N hm-2yr-1,MN),高氮添加(200 kg N hm-2yr-1,HN),每种处理均设置3个重复,每个林分共12个样方。为减少样方之间因为氮添加梯度而产生的相互干扰,样方至少间隔10m。模拟氮沉降于2020年10月28日开始,每隔两个月一次。模拟氮沉降时将NH4NO3溶于水后采用背式喷雾器均匀喷洒对应的处理样方(各样方施氮面积9 m2)。为避免额外降水导致的差异,对对照(CK)样方喷洒同样体积的水。除施氮水平不同外,其他措施保持一致。
1.3 土壤取样及实验室分析
在2021年1月1日、3月4日、4月16日、7月6日、9月8日、10月28日,在各样方中利用五点取样法,采集0~20cm土壤样品。同时,在样方内挖土壤剖面并以环刀取0-20cm土层的原状土。土壤样品带回实验室后,剔除石块及动植物残体,将土样分成两部分,一份放入冰盒带回实验室过2mm筛,放入低温冰箱冷藏保存并在一星期内测完土壤酶活性。另一份自然风干,过100目筛,用于测定元素含量。原状土带回实验室后于105℃烘干至恒重利用环刀法测定土壤容重。土壤碳含量采用重铬酸钾外加热法测定。土壤β-1,4-N-acetyl-glucosaminidase(βG)酶活性采用酶标仪测定。
1.4 数据处理
土壤有机碳密度(kg/m2)利用公式(1)计算:

使用SPSS 22.0、Office 2019、和Canoco 5.0进行数据分析,使用Origin 2018和R语言作图。
2.结果与分析
2.1 氮沉降对土壤有机碳含量的影响
麻栎的CK、LN、MN和HN土壤有机碳分别平均为3.24±0.49%、3.05±0.47%、3.32±0.49%和3.53±0.48%(图1)。黑松的CK、LN、MN和HN土壤有机碳含量平均3.62±0.53%、3.75±0.46%、3.79±0.43%和3.86±0.39%。随着不同浓度的氮的添加,麻栎林分中,中氮和高氮处理的平均有机碳含量有轻微的增加,在黑松林分施氮处理的有机碳的平均含量都高于对照。

图1 麻栎(A)和黑松(B)土壤有机碳变化
2.2 氮沉降对土壤βG酶活性的影响
麻栎土壤βG酶活性表现出明显的季节性变化(表2),在7月份的达到峰值,麻栎低氮处理除了在9月份的时候对酶活性没有显著影响,在其余月份都显著高于对照处理,麻栎中氮处理在1月和10月酶活性显著高于对照处,麻栎高氮处理除了在3月和9月对酶活性无显著影响,在剩下的采样月份中都显著高于对照处理,各 处理平均酶活性大小排序为LN(10.82 nmol·g-1·h-1)>HN(7.58 nmol·g-1·h-1)>MN(6.32 nmol·g-1·h-1)>CK(4.00 nmol·g-1·h-1)。黑松βG酶活性在4月份的达到峰值21.45 nmol·g-1·h-1。黑松低氮和中氮处理除了3月和4月,在其余月份都显著高于对照处。黑松高氮处理除了在3月对酶活性无显著影响,在剩下的采样月份中都显著高于对照处理。各处理平均酶活性大小排序为HN(18.08 nmol·g-1·h-1)>LN(14.64 nmol·g-1·h-1)>MN(13.40 nmol·g-1·h-1)>CK(10.45 nmol·g-1·h-1)。

表 2 麻栎(A)和黑松(B)土壤βG酶活性变化
2.3 氮沉降对麻栎和黑松土壤碳密度的影响
在模拟氮沉降一年后,各处理土壤有机碳密度差异不显著。氮沉降均会促进麻栎林土壤有机碳分解,降低土壤有机碳密度。具体表现为 MN(0.7823gm-2) 图3 麻栎(A)和黑松(B)土壤有机碳密度 在森林生态系统中,土壤有机碳主要来自于凋落物分解。而土壤有机碳的主要损失途径为微生物利用有机碳过程中呼吸作用释放的CO2(徐小锋等,2007)。因此本研究中氮添加对土壤有机碳密度的影响,主要是通过影响凋落物分解和酶活性实现的。研究表明,凋落叶基质组分和环境条件是调节分解的关键因素,而氮添加通过直接调节微生物活动,改变土壤理化特征,进而导致凋落物的分解和土壤碳循环(邱尔发等, 2005)。在本研究中,模拟氮沉降处理土壤有机碳含量均高于对照处理,说明氮沉降会促进凋落物分解,增加土壤有机碳库的来源(杨玉盛等,2004)。在本研究中,氮沉降一年后会促进麻栎林土壤有机碳分解,导致土壤有机碳密度降低。而氮沉降则会促进黑松林土壤有机碳固存。这可能是由于麻栎的阔叶属于优质碳源,随着优质碳源的输入可能会引起土壤激发效应,导致土壤有机碳以CO2的形式损失,从而降低了麻栎土壤有机碳密度(王棣等,2015)。而黑松由于凋落物木质素含量较高,难以分解并以有机碳的形式固存在土壤中(马志良等,2015)。 氮沉降会促进凋落物分解从而增加土壤有机碳库的来源。但根据凋落物本身性质的差异,氮沉降并不一定会提高有机碳密度,促进土壤碳固存。在本研究中表现为,氮沉降会导致麻栎土壤有机碳密度降低,同时会提高黑松林的土壤有机碳密度。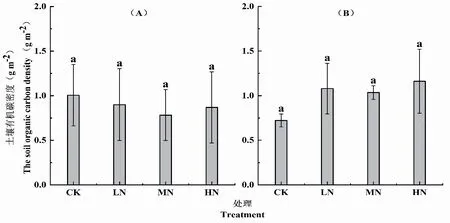
3.讨论
4.结论
猜你喜欢
广东蚕业(2022年4期)2022-11-23 13:31:55
安徽农业科学(2021年9期)2021-06-22 15:02:49
湖北农机化(2020年16期)2020-10-21 01:29:08
江西农业(2020年13期)2020-09-08 07:44:54
环境科技(2016年1期)2016-11-08 12:17:44
防护林科技(2016年1期)2016-02-27 06:09:54
中国铸造装备与技术(2015年5期)2015-12-10 10:23:42
西藏科技(2015年4期)2015-09-26 12:12:59
现代企业文化(2015年1期)2015-02-28 18:58:20
粘接(2014年1期)2014-04-29 00:44:03
