
重金属胁迫下四种红树植物幼苗生理响应特征
2022-12-05 12:53:48谢勇王友绍
热带海洋学报 2022年6期
谢勇 , 王友绍
1. 热带海洋环境国家重点实验室(中国科学院南海海洋研究所), 广东 广州 510301;
2. 中国科学院大亚湾海洋生物综合实验站, 广东 深圳 518121;
3. 中国科学院南海生态与环境工程创新院, 广东 广州 510301;
4. 中国科学院大学, 北京 100049;
5. 南方海洋科学与工程广东实验室(广州), 广东 广州 511458
随着工农业的发展,经济效益提升的同时带来了较大的污染问题,其中重金属污染是一种常见的污染。由于重金属污染物不能被生物降解,存在时间长,可通过食物链富集,危害人类健康,因而受到广泛的关注(王宏镔 等, 2005,Sodango et al, 2018)。特别是沿海湿地的重金属污染问题较为突出,与其他湿地生态系统一样,红树林生态系统也经历了陆源污染物输入,而城市化的快速发展也加剧了红树林区污染,使其成为重金属污染物的重要储存库 (Usman et al, 2013; Bourgeois et al, 2019)。邓利 等(2014)发现深圳福田红树林自然保护区沉积物中重金属 Cu、Pb、Zn 等全部超过《海洋沉积物质量》(GB 18668-2002)中I 类标准。这些研究表明红树植物正遭受着不同程度的重金属污染胁迫,高浓度的重金属胁迫能够干扰植物一些保护酶的活性,导致活性氧(reactive oxygen species, ROS)大量累积,ROS 可以攻击生物膜上的不饱和脂肪酸,造成氧化损伤(Hegedüs et al, 2004),进而对植物的光合作用和呼吸作用产生影响,限制植物生长发育,严重时导致植物死亡。在重金属污染胁迫下红树植物的生长与生理生化响应已经有较多研究(Zhang et al, 2007; 王友绍, 2019, 2021; 王芳洲 等, 2020; Wang et al,2021),但是关于不同红树植物抗重金属胁迫能力研究相对较少(王友绍 等, 2019)。
本实验选取 4 种常见的红树科植物, 红海榄(Rhizophora stylosa)、秋茄(Kandelia obovate)、木榄(Bruguiera gymnorhiza)和角果木(Ceriops tagal)为研究对象,分析在不同浓度重金属水溶液胁迫下四种红树植物幼苗叶片的生理生化变化,以筛选出抗重金属能力更强的红树物种,为受损湿地人工种植选种提供参考,对红树林的生态恢复、保护与持续发展具有重要意义。
1 材料与方法
1.1 实验材料
实验采用的红海榄、秋茄、木榄、角果木幼苗均采自广东省湛江市红树林育苗基地。所有红树幼苗均挑选六叶期的幼苗进行培养,用自来水冲洗干净之后分别栽培于装有沙子的塑料盆中,每三天用1/2 Hoagland’s 营养液浇灌一次,以补充水分和营养物质。Hoagland’s 营养液是植物培养中的常用营养液,主要成分包括铁盐、硝酸盐、磷酸盐、硫酸镁和微量元素等。幼苗培养两周后, 挑选无机械损伤、无虫害的长势良好的幼苗移至人工气候培养箱进行培养,培养条件设置如下:温度26℃、相对湿度75%、光照强度20000lux,光照周期12h·d–1,在人工气候箱中进行培养驯化。
1.2 实验方法
将培养驯化好的红树幼苗各自分为5 组,用1/2 Hoagland’s 营养液配置成4 个浓度梯度的复合重金属水溶液分别进行胁迫处理,每3 天浇灌一次,每次浇灌600mL 复合重金属水溶液。依据预实验结果设置了如下复合重金属水溶液浓度,具体含量见表1。复合重金属水溶液胁迫处理28d 后剪取四种植物的叶片进行各生理生化指标的测定(Zhang et al,2007)。
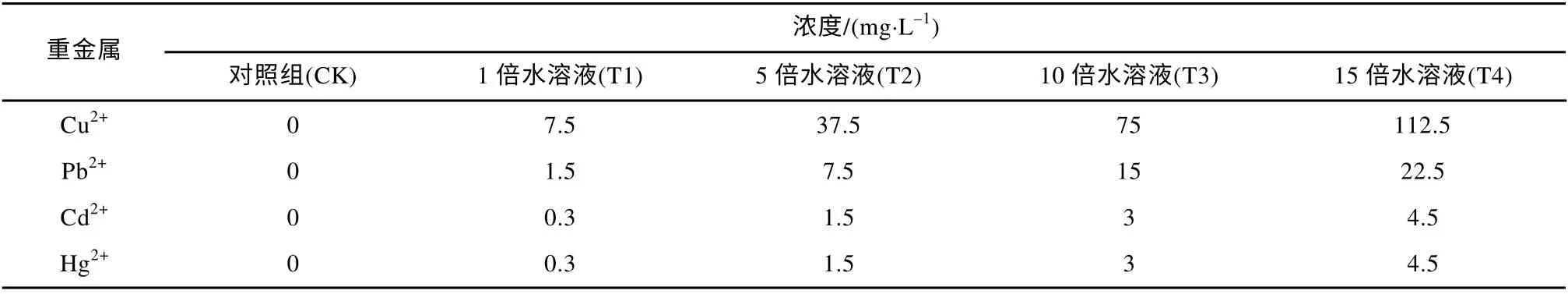
表1 不同处理重金属浓度Tab. 1 Concentrations of heavy metals in the different treatments
1.3 植物叶片可溶性蛋白的测定
可溶性蛋白的测定参照考马斯亮蓝测蛋白法(Bradford, 1976),标准曲线的绘制及注意事项参考考马斯亮蓝法测定蛋白质含量中细节问题的描述(蒋大程 等, 2018)。
1.4 植物叶片SOD、POD、MDA 的测定
氯化硝基四氮唑蓝(NBT)光还原法测植物超氧化物歧化酶(superoxide dismutase, SOD)活性、愈创木酚氧化法测植物过氧化物酶(peroxidase, POD)活性、 硫代巴比妥酸比色法测植物丙二醛(malondialdehyde, MDA)含量,所用试剂盒均由南京建成生物工程研究所提供。
1.5 数据处理
本实验所有指标的测定均作三次生物学重复,最后的结果用平均值±标准差表示,用 Graphpad prism 7 进行作图分析。使用SPSS(Version 21)进行单因素方差分析、Pearson 相关性分析和主成分分析(principal component analysis, PCA),直观地反映各生理生化指标之间的关系,标注*的数据代表与对照组具有显著性差异(P<0.05)。
2 实验结果
2.1 复合重金属水溶液胁迫对四种红树植物叶片可溶性蛋白含量的影响
由图1 可知,重金属胁迫下红海榄叶片可溶性蛋白的含量除T2 组略高于对照组外,其余各组均低于对照组,但差异不显著(P>0.05)。秋茄叶片的可溶性蛋白含量在各处理组均略低于对照组,差异不显著(P>0.05),木榄叶片可溶性蛋白含量除T2 组显著低于对照组外,其余各组与对照组相比差异不显著(P>0.05)。角果木叶片可溶性蛋白含量除T1 组外均显著低于对照组(P<0.05)。
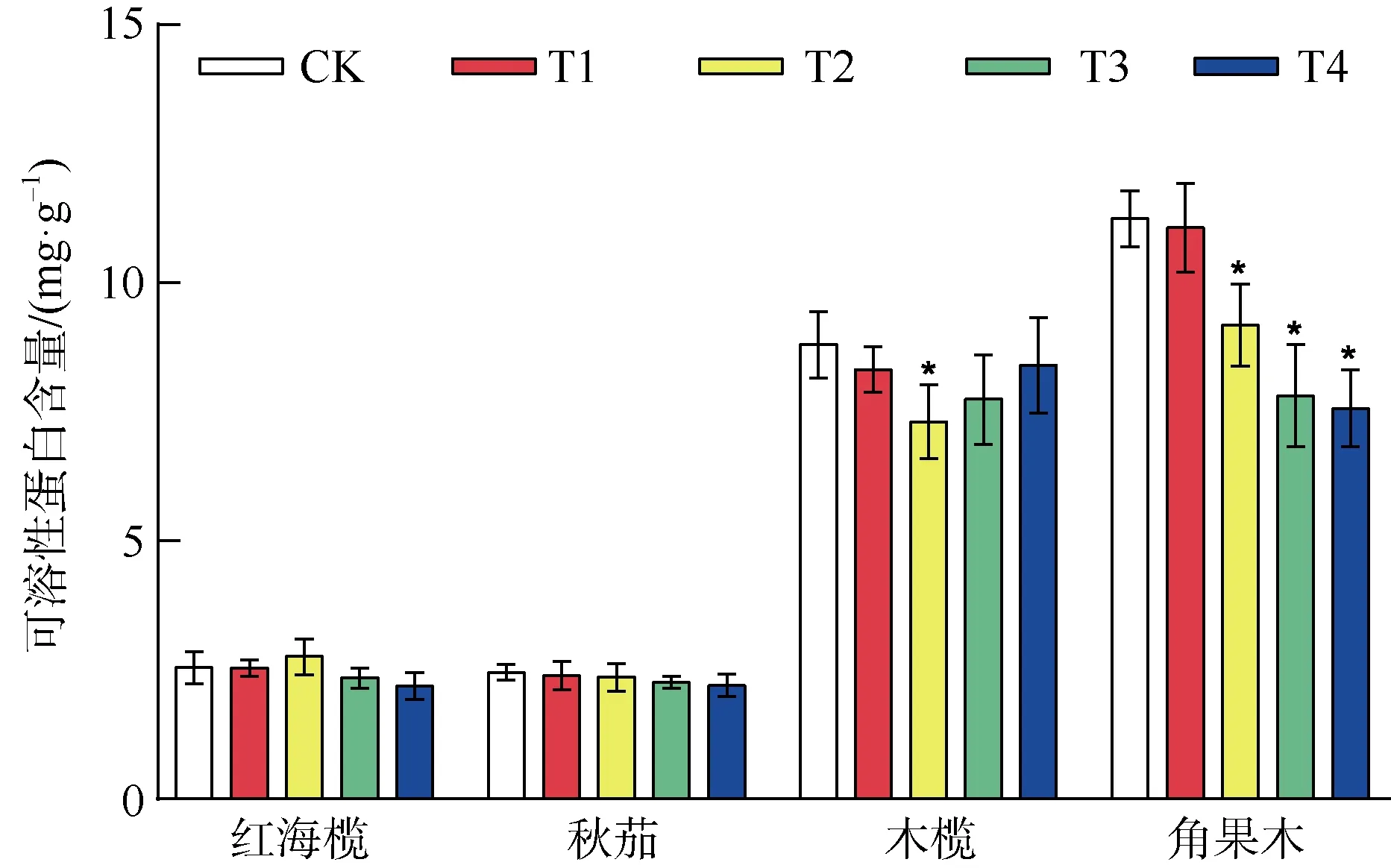
图1 重金属胁迫对四种红树植物可溶性蛋白含量的影响Fig. 1 The effect of heavy metal stress on the soluble protein content of four mangrove plants
2.2 复合重金属水溶液胁迫对四种红树植物叶片SOD 活性的影响
由图2 可知,复合重金属水溶液胁迫可以增加四种植物幼苗叶片的SOD 活性,随重金属水溶液处理浓度的提高,四种红树植物叶片中SOD 活性均表现出先升高再降低的趋势,红海榄叶片SOD 活性在各处理组均显著高于对照组(P<0.05),在T2 处理组达最大值,其中T2 组与T3 组SOD 活性很接近,分别为对照组的1.55 倍、1.52 倍,且均显著高于对照组。秋茄和木榄叶片SOD 活性均在T2 处理组达到最大值,分别为对照组的1.39 倍和1.56 倍。角果木叶片在T1 处理组达到最大值,为对照组的1.36倍,当重金属水溶液浓度超过T1 组浓度后,随重金属浓度的进一步提高SOD 活性开始下降,在T4 组时甚至低于对照组的活性。

图2 重金属胁迫对四种红树植物SOD 活性的影响Fig. 2 The effect of heavy metal stress on SOD activity of four mangrove plants
2.3 复合重金属水溶液胁迫对四种红树植物叶片POD 活性的影响
由图3 可知重金属水溶液胁迫处理后四种红树植物叶片内POD 活性除木榄的T4 处理组、角果木的T3、T4 处理组外,叶片内POD 活性均显著高于对照组(P<0.05),四种红树植物叶片中POD 活性均表现出随重金属水溶液处理浓度的增加先升高再下降的趋势,红海榄叶片POD 活性在T3 处理组达最大值,为对照组的1.77 倍; 秋茄、木榄叶片中POD活性在T2 处理组达最大值,分别为对照组的1.58倍、1.67 倍,角果木在T1 处理组达最大值,是对照组的1.59 倍,当重金属水溶液浓度超过T1 后,表现出下降的趋势。
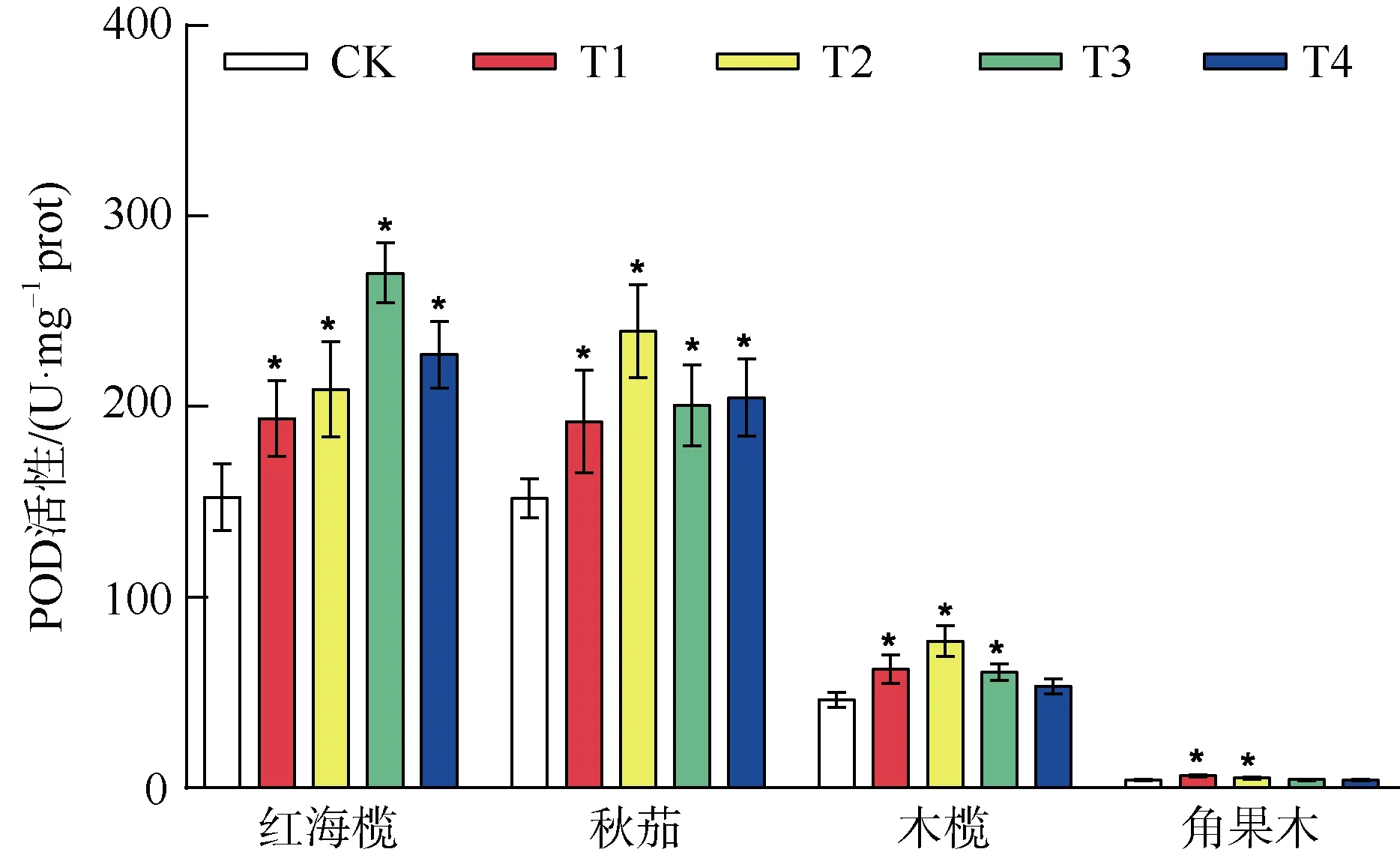
图3 重金属胁迫对四种红树植物POD 活性的影响Fig. 3 The effect of heavy metal stress on POD activity of four mangrove plants
2.4 复合重金属水溶液胁迫对四种红树植物叶片MDA 含量的影响
如图4 所示,复合重金属水溶液处理下四种红树植物幼苗中的MDA 含量基本呈现逐步升高的趋势,其中红海榄在较低浓度处理组T1、T2 组的MDA含量高于对照组,但并不显著(P>0.05),秋茄在T1处理组略低于对照组,木榄、角果木在T1 处理组高于对照组但并不显著(P>0.05),其他各组 MDA含量均显著高于对照组(P<0.05)。在T4 处理组时,四种红树植物叶片内MDA 含量达最大值,红海榄叶片内MDA 含量最高为对照组的1.82 倍,秋茄是对照组的2.23 倍,木榄是对照组的2.25 倍,角果木是对照组的2.73 倍。
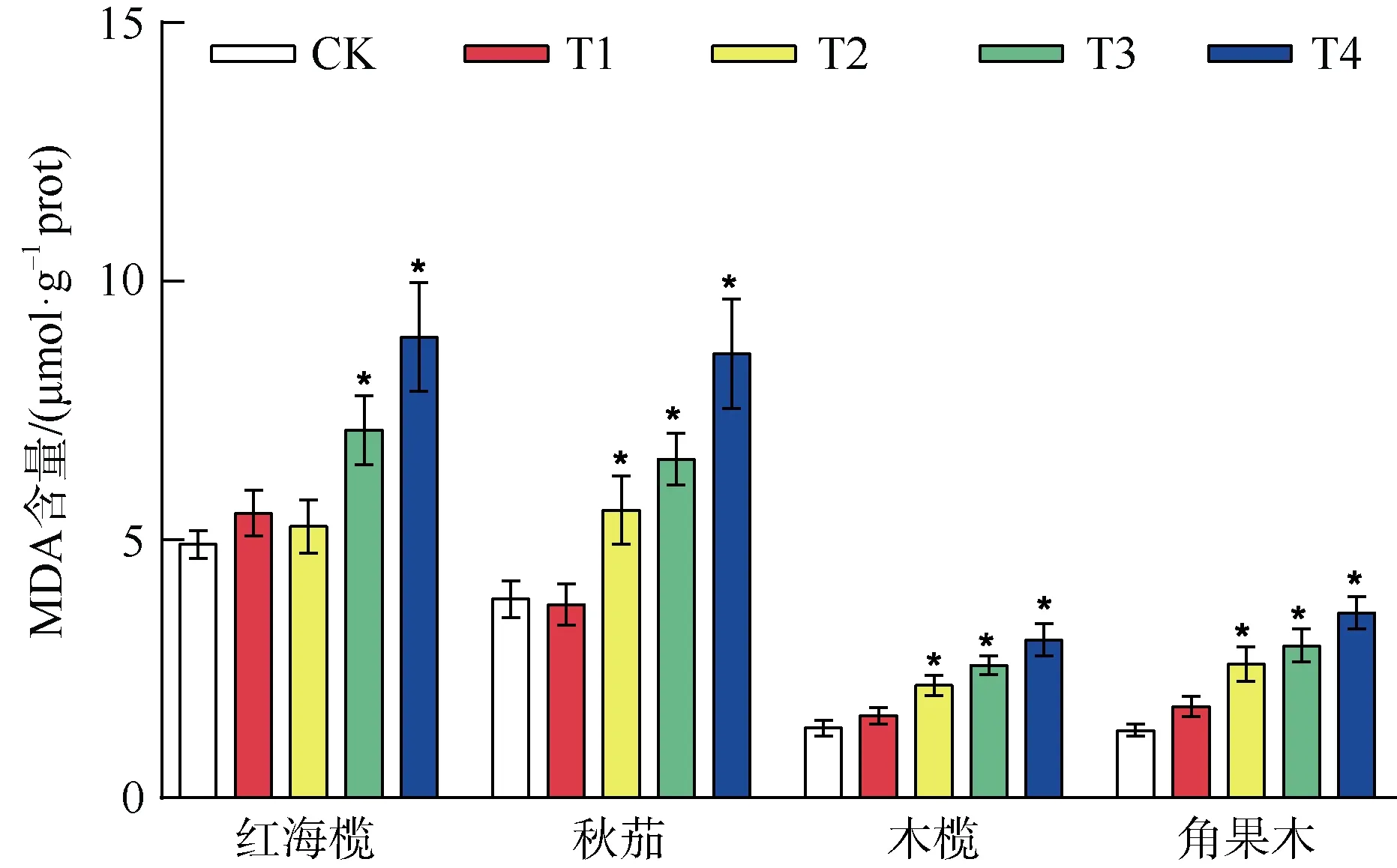
图4 重金属胁迫对四种红树植物MDA 含量的影响Fig. 4 The effect of heavy metal stress on MDA content of four mangrove plants
2.5 各指标间的相关性分析
Pearson 相关性分析结果表明(表2),红树植物叶片内可溶性蛋白含量与POD、MDA 呈极显著负相关关系(P<0.01),MDA 含量与POD 活性成极显著正相关关系(P<0.01),SOD 活性与其他几项指标相关性不显著(P>0.05)。
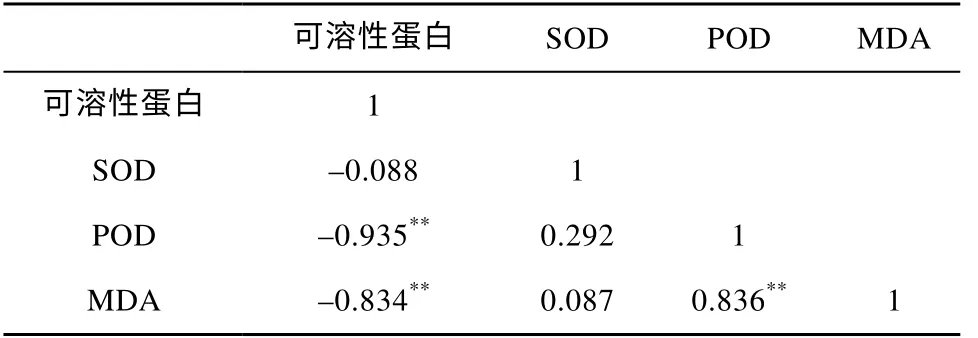
表2 重金属胁迫下各生理生化指标间的相关性Tab. 2 Correlation between various physiological and biochemical indexes under heavy metal stress
利用SPSS 做主成分分析,对生理生化指标进行降维处理,可以直观体现重金属水溶液胁迫下生理生化指标间的变化关系。结果如图5 所示,生理生化指标分为2 个主成分,PC1 描述了总方差的73.030%,PC2 描述了总方差的25.926%,二个主成分合计描述了总方差的98.956%。其中MDA 和POD距离较近,说明二者关系密切。
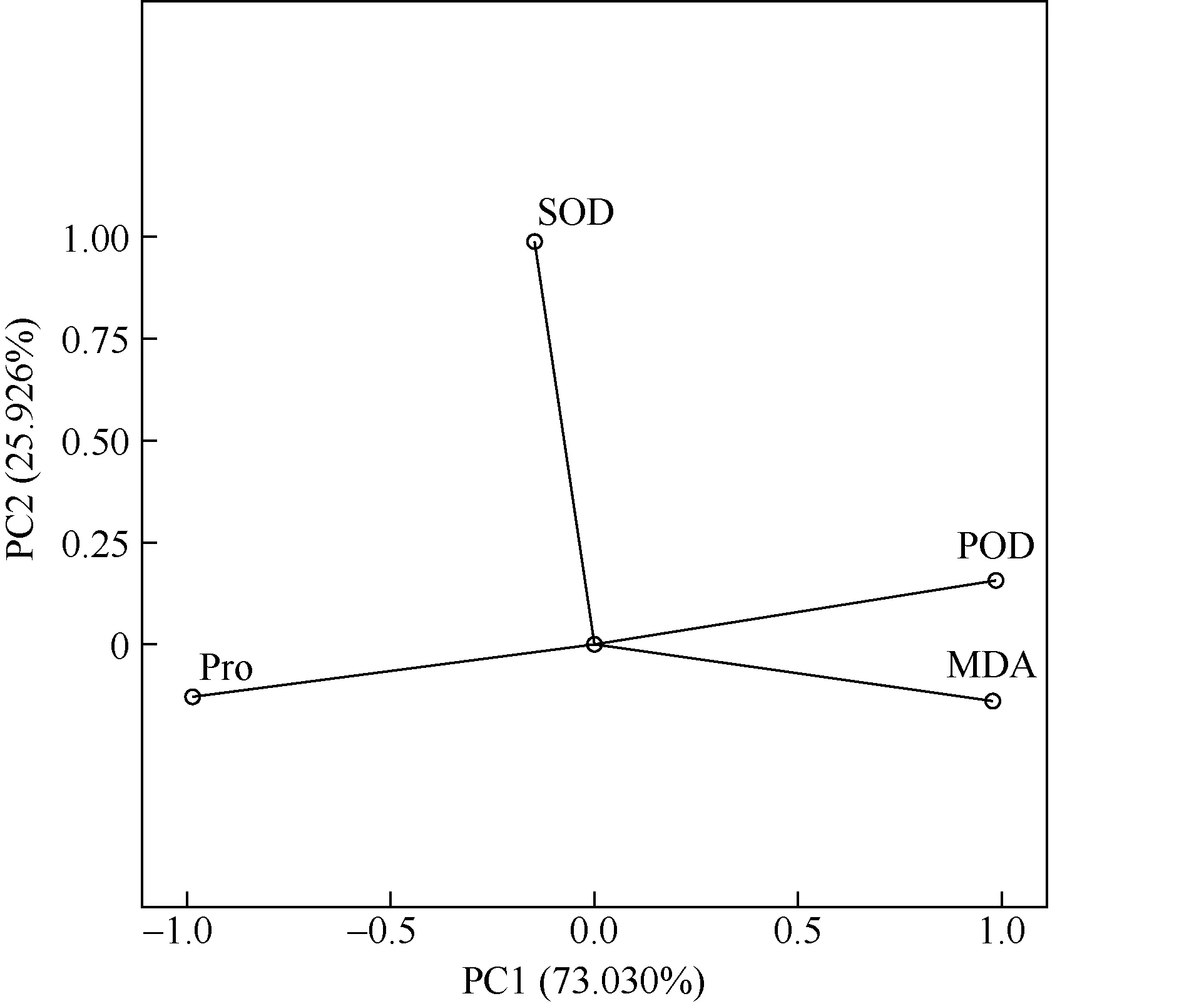
图5 重金属胁迫下各生理生化指标的PCA 分析Fig. 5 PCA analysis of various physiological and biochemical indexes under heavy metal stress
3 讨论
植物体内的可溶性蛋白多是参与各种代谢活动的酶类,其含量越多代表代谢活动越强,因此可溶性蛋白含量的多少可以反映植物体代谢活动的强弱(孙天国 等, 2010)。另外由于重金属离子的存在,外界渗透势会降低,细胞内的水会向低渗透势的外界流失,导致植物缺水,对植物造成损伤。可溶性蛋白含量增加可以降低细胞内的渗透势,提高植物的保水能力, 从而避免缺水导致的损伤。可溶性蛋白含量的多少可以代表植物调节渗透压能力的强弱,因此可作为关键的生理生化指标,指示植物的抗逆性强弱和代谢情况(赵胡 等, 2016)。本实验中,红海榄、秋茄叶片内可溶性蛋白含量略小于对照组,但差异不显著(P>0.05),木榄叶片除T2 组可溶性蛋白含量显著低于对照组外,其余各组与对照组相差不大,表明红海榄、秋茄、木榄对重金属胁迫有比较好的抗性,可溶性蛋白的含量能保持在一个较为正常的水平,维持渗透平衡,保证细胞代谢活动的正常进行,而角果木叶片可溶性蛋白含量除T1 组外均显著低于对照组(P<0.05),说明角果木在遭受超过1 倍重金属水溶液浓度胁迫时,叶片代谢活动减弱,调节渗透压的能力减弱,重金属胁迫对角果木叶片的蛋白质合成造成了影响。
氧化损伤是植物受重金属胁迫伤害的主要机理之一,重金属胁迫会导致ROS 的过度产生和积累(Sch ü tzendü bel et al, 2002),高水平的 ROS 通常会破坏细胞成分,如细胞膜、核酸和叶绿素等(Tewari et al, 2002),造成氧化损伤,影响植物发生长发育。低浓度的重金属胁迫会激活植物的抗氧化系统,诱导SOD、POD 等抗氧化酶活性增加,清除植物体内活性氧 (Srivastava et al, 2004),抗氧化酶的诱导是减少氧化损伤的重要保护机制,其中SOD 在诱导下活性增加,催化超氧阴离子转化为过氧化氢和氧气,过氧化氢也是一种自由基,也能对细胞膜造成氧化损伤,POD 则可以催化过氧化氢生成无毒害的水和氧气,使植物体内的自由基保持在较低水平,从而避免自由基对植物造成氧化伤害,是逆境下植物体内重要的保护酶(Blokhina et al, 2003)。本研究中,随着重金属水溶液处理浓度的增加,四种红树植物幼苗叶片内的SOD、POD 活性均呈现出先升高再降低的变化趋势,这与Cu2+处理下秋茄幼苗叶片的抗氧化酶变化情况一致(赵胡 等, 2016),低浓度的重金属胁迫诱导了抗氧化酶活性的提高,当重金属浓度超过承受阈值后,植物体内的活性氧含量超过了正常的歧化能力,而对组织和细胞多种功能膜及酶系统造成破坏,反过来抑制了植物叶片内SOD、POD等抗氧化酶的活性。红海榄叶片中POD 活性在T3处理组达到最大值; SOD 活性在T2 处理组时达到最大值,其中T3 组与T2 组SOD 活性非常接近,无明显差距且均显著高于对照组,分别是对照组活性的1.52 倍和1.55 倍。秋茄、木榄叶片中POD、SOD活性均在 T2 处理组达最大值,而角果木叶片中POD、SOD 活性均在T1 处理组达到最大值,随后再增加重金属水溶液处理浓度,SOD、POD 活性开始受到抑制,活性下降。说明红海榄在T3 处理组,秋茄、木榄在T2 处理组,角果木在T1 处理组的重金属胁迫时具有最积极的抗氧化酶活性,此时清除细胞内的ROS 能力最强,当重金属处理浓度持续增加,超过了植物承受阈值时,抗氧化酶活性受到抑制,清除ROS 能力减弱,导致植物过氧化水平加剧,进而破坏膜结构,影响蛋白质合成等各种代谢活动,对植物产生损害。当逆境发生时,植物会采取各种措施提高自身的抵抗力以适应逆境压力,但当逆境压力超过植物的耐受极限时,其自身防御系统也会遭到破坏(王友绍,2019)。
植物在遭受重金属胁迫时,体内会积累大量ROS,这些活性氧会攻击生物膜上的不饱和脂肪酸,发生脂质过氧化反应,产生MDA,MDA 的产生能加剧膜的损伤,因此可以通过MDA 含量间接反映膜的受损程度(Knight et al, 1990)。本研究中,重金属水溶液胁迫增加了四种红树植物叶片中的MDA含量,当重金属水溶液浓度增大时,叶片中的MDA含量持续增加,膜系统损伤加剧,植物受损程度加大。其中,红海榄在T1、T2 浓度时MDA 含量虽然增加,但与对照相比,MDA 增加并不显著(P>0.05),表明T2 浓度的重金属水溶液处理对红海缆的损伤并不严重,同样的,T1 浓度的重金属污染对秋茄、木榄、角果木的损伤也不严重。红海榄、秋茄、木榄、角果木四种植物在最高浓度处理组时, MDA含量分别为对照组的1.82、2.23、2.25、2.73 倍,表明在高浓度重金属水溶液胁迫下,红海榄受氧化损伤程度较低,秋茄、木榄受损程度相当,角果木受损程度最重。
Pearson 相关性分析结果显示红树植物叶片内可溶性蛋白含量与POD、MDA 呈极显著负相关关系(P<0.01),MDA 含量与POD 活性成极显著正相关关系(P<0.01),SOD 活性与其他几项指标相关性不显著(P>0.05)。MDA 含量的增加表明植物遭受重金属胁迫后发生氧化损伤,植物体内的氧自由基、过氧化物增多,刺激植物合成SOD、POD 等抗氧化酶,以清除体内的过氧化物,减少氧化损伤,所以MDA与POD 呈正相关关系。而重金属离子进入细胞后会与细胞内的化合物结合,影响植物代谢活动,特别是影响蛋白质的合成,所以MDA 含量与可溶性蛋白含量呈现极显著负相关关系。从PCA 分析图中可知,将四个生理生化指标分解到2 个主成分中,可以解释总方差的98.956%,其中POD 和MDA 距离最近,表明二者相关性很强,当重金属胁迫产生氧化损伤时,MDA 含量增加,POD 活性也会提高,以清除氧自由基,减少氧化损伤。
4 结论
本实验结果显示,四种红树植物幼苗在遭受重金属水溶液胁迫时,均表现出一定的抵抗重金属胁迫能力,SOD、POD 两种抗氧化酶能在一定程度上能够清除重金属胁迫产生的氧自由基,保护膜结构,避免氧化损伤。但是这种抵抗重金属胁迫的能力有一定的限度,当重金属浓度超过承受极限时,抗氧化酶的活性将会被抑制,活性氧大量积累,氧化损伤加剧。其中,红海榄在10 倍重金属水溶液处理时仍具有较为积极的抗氧化酶活性, 秋茄、木榄在5 倍重金属水溶液处理时具有较为积极的抗氧化酶活性, 角果木在1 倍重金属水溶液处理时抗氧化酶活性最强,表明红海榄抗氧化酶系统承受重金属污染能力强于秋茄、木榄,角果木抗氧化酶系统承受重金属污染能力最弱。高浓度重金属水溶液处理下角果木受氧化损伤程度最高,木榄与秋茄相当,红海榄受氧化损伤程度最低。红海榄的抗重金属胁迫能力较强,秋茄与木榄的抗重金属胁迫能力相当,角果木抗重金属胁迫能力最弱。四种红树科植物中,红海榄的重金属抗性最强,角果木的重金属抗性最弱,在受重金属污染严重的海岸带更适合种植红海榄进行植物修复,恢复生态。本实验筛选出了抗重金属能力较强的红树种类红海榄,其相关抗性基因有待进一步研究。
猜你喜欢
热带林业(2023年3期)2023-10-30 10:38:34
小猕猴学习画刊(2022年4期)2022-05-05 01:52:59
广东第二课堂·小学(2021年10期)2021-12-03 04:35:50
世界科学技术-中医药现代化(2021年12期)2021-04-19 12:32:10
海洋通报(2020年5期)2021-01-14 09:27:04
天然产物研究与开发(2016年6期)2016-06-05 10:29:31
浙江大学学报(工学版)(2016年9期)2016-06-05 09:20:57
艺术评鉴(2016年13期)2016-05-30 16:19:05
中学生理科应试(2016年2期)2016-05-30 14:23:28
广东石油化工学院学报(2016年3期)2016-05-17 05:16:21
