
柚木无性系早期选择年龄的研究
2022-12-03 07:36:20付强黄桂华周强王先棒周再知张绍祥梁坤南
中南林业科技大学学报 2022年10期
付强,黄桂华,周强,王先棒,周再知,张绍祥,梁坤南
(1.云南省普洱市林业和草原种苗工作站,云南 普洱 665000;2.中国林业科学研究院热带林业研究所 热带林业研究国家林业和草原局重点实验室,广东 广州 510520;3.云南省景谷傣族彝族自治县林业技术推广中心,云南 景谷 666400)
林木培育周期长,早期测定对于缩短育种周期和加速世代周转具有重要作用[1]。利用生长性状的早晚相关进行早期选择,可有效加快育种进程。早在20 世纪70 年代,国内学者对林木早期选择就有报道,先后对杉木[2-5]、马尾松[6]、黄山松[7]、火炬松[8]、长白落叶松[9]、杂种落叶松[10]、湿加松[11]、油松[12]、木荷[13]等树种的天然林和人工林进行树干解析或种源/家系试验,通过年-年秩次相关分析等方法确定各个树种、种源和家系早期选择的最佳年龄。在无性系早期年龄选择方面,赵承开[14]研究了107 个杉木无性系认为杉木优良无性系初始年龄为5~6 a,精选年龄8~9 a;马常耕等[15]对17~18 个无性系历年生长分析,认为杉木适宜的早期选择年龄4~6 a;胡德活等[16]研究了22 个和106 个杉木无性系,认为早期选择最佳年龄为3~4 a;吴德军等[17]认为刺槐建筑材轮伐期15 a,无性系早期选择年龄3~4 a,选择有效。对泡桐15 年生无性系来说,3 a 可作为优良无性系早期选择年龄[18]。白榆无性系可根据4 年生的胸径和材积进行早期选择[19]。
柚木Tectona grandis自然分布于缅甸、印度、泰国和老挝[20],被广泛引种到自然分布区外的亚洲、非洲、拉丁美洲和大洋洲等70 多个国家和地区[21]。我国早在1820 年前后引种柚木于云南省边境寺庙等,作为庭院绿化树种[1]。由于其速生、材性优良,现已遍及我国云南、台湾、海南、广东、广西、福建、四川和贵州等地,热带、南亚热带地区或干热河谷热区的80 多个县市均有引种或规模化种植。我国20 世纪70 年代开展了柚木系统的遗传改良,收集和引进种质资源,建立种源/家系试验,选出一批优良种源和家系[22-23]。随着20世纪80 年代组培技术的突破[24-25]和90 年代无性系规模化生产的实现[26-27],2000 年起陆续在广东、广西、福建、海南、云南和贵州等柚木适生和次适生区的20 多个县市建立了无性系区域性试验林。由于柚木培育周期较长,传统方式40~60 a,但随着柚木遗传改良程度和无性繁殖技术的提高、优良无性系的应用和集约栽培,轮伐期已缩短到20~30 a[28]。但为更快地选出优良无性系用于造林,研究柚木无性系早期选择尤为必要,我国开展柚木无性系试验时间不长,尚未见到无性系早晚相关与早期选择年龄研究的报道。本研究以2010 年建于云南景谷的无性系测定林的多年生长指标为对象,研究无性系早期生长、早晚性状相关以及早期选择年龄,以期为无性系早期选择提供科学依据。
1 材料与方法
1.1 试验地概况
试验地位于威远江河谷地带的云南省普洱市景谷傣族彝族自治县益智乡(23°13′51″N,100°35′2″E),属南亚热带季风气候。年平均气温21℃,年降水量1 190 mm,雨季旱季明显,5—10月为雨季,11 月至翌年4 月为旱季。土壤为砖红壤,pH 值5.6。海拔850 m。
1.2 试验材料
参试无性系共23 个,由中国林业科学研究院热带林业研究所从柚木人工林和种源试验林中选优,采集顶芽组培繁殖。以海南尖峰岭无性系种子园采集的1 个家系种子培育的实生苗作为对照,其中,有11 个无性系是来自4 个印度种源,1 个无性系来自尼日利亚种源,2 个无性系是缅甸原生种源,10 个是原产缅甸的我国次生种源(表1)。参试苗木在热带林业研究所苗圃培育成1 年生营养袋苗,运输前除去袋土,截干造林。

表1 柚木无性系试验材料Table 1 The experimental materials of teak clones
1.3 试验设计与造林措施
24 个参试材料采用完全随机区组设计,6 株单行小区,6 次重复(区组),株行距为2.5 m×4.0 m。造林地砍杂清理后,穴状整地,穴规格为60 cm×60 cm×50 cm。每穴施基肥1.0 kg 钙镁磷肥和1.0 kg 石灰,回土搅拌均匀后再回土填满成龟背状。2010 年6 月9—10 日种植。3 年抚育8 次,追肥4 次。
1.4 试验观测与统计分析
对试验林1、3、5、7、9 和11 年生的树木进行每木的树高、胸径生长(1 年生为地径)测定,3 年生以后的单株材积按V=0.4787×H×DBH2[29]计算。采用GENSTAT[30]软件进行方差分析和主成分分析,采用ASReml-R 软件[31]进行遗传分析等。
主成分分析因子得分方程(主分量方程)[32]为:
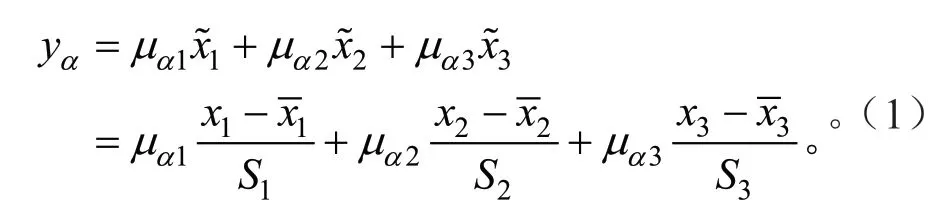
式中:yα为某一无性系在第α主分量得分值(α=1,2,3),μα1、μα2和μα3分别为性状1、2 和3 在第α主分量的特征向量,x1、x2和x3分别为性状1、2 和3 的原数据,为性状1、2 和3的原数据平均值,S1、S2和S3为性状1、2 和3 原数据的标准差。
无性系重复力估算公式[33]为:

式中:F为方差分析的F值。
无性系广义遗传力(H2)的估算公式为:
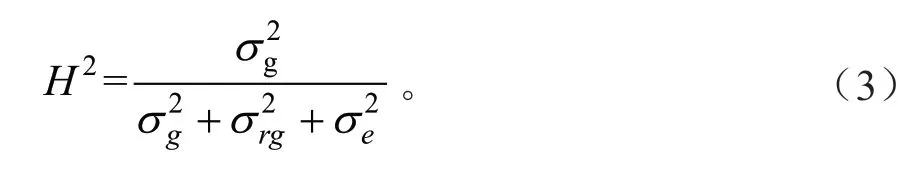
2 结果与分析
2.1 柚木无性系历年生长差异分析
对1、3、5、7、9 和11 年生的树高和胸径进行方差分析,结果(表2)表明,1~11 年生无性系间的树高和胸径生长差异均达极显著(P<0.001),并且F值均随着树龄的增加而增加,7 年生时达到最大,然后下降。1~11 年生区组间的差异从1 年生树高生长显著到3 年生和之后的树高和胸径生长均极显著,区组间差异(F值)有随树龄增加而增大的趋势;树高生长无性系与区组间的交互作用除1 年生树高生长无交互作用外,其余年份交互作用极显著,各无性系在区组中的排序不一。从表2 还可以看出,无性系(试验林)的树高和胸径变异系数分别为4.03%~9.38%和6.90%~9.98%,同一年份胸径变异系数大于树高变异系数,但均随着树龄的增加而减低,于5年生后趋向稳定。
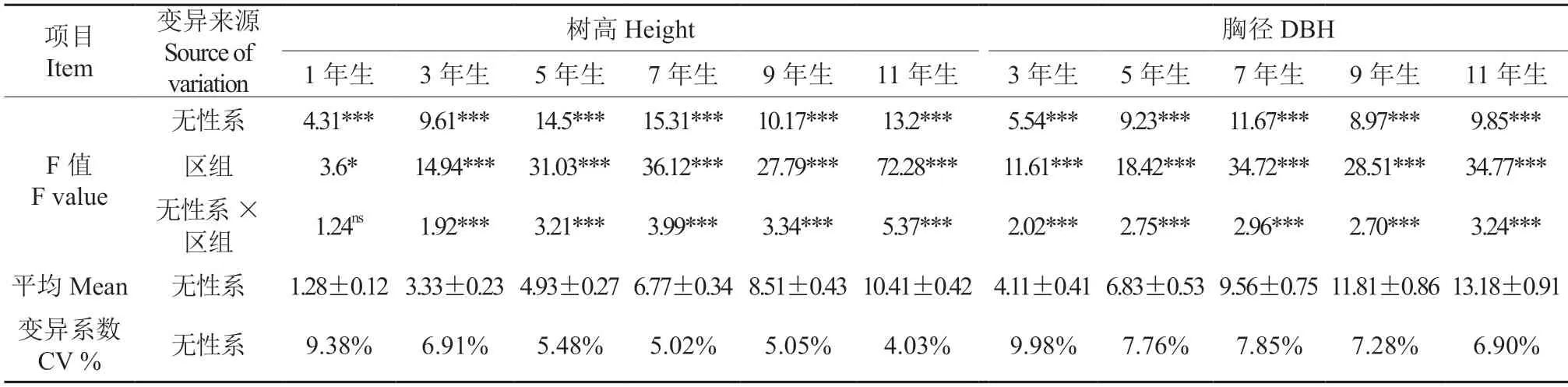
表2 柚木无性系历年树高和胸径生长方差分析†Table 2 Variance analysis of the growth of height and DBH of the teak clones at different ages
2.2 柚木无性系生长性状遗传分析
从表3 可以看出,1~11 年生柚木无性系树高重复力在0.768~0.935 之间和遗传力在0.110~0.310 之间,3~11 年生胸径重复力在0.819~0.914之间和遗传力在0.135~0.252 之间;树高和胸径生长的重复力和遗传力先逐年缓慢增加,7 年生达到最大值,然后缓慢下降并趋于稳定,11 年生的树高和胸径生长的重复力大小与5 年生持平;同样,11 年生树高遗传力与3 年生、11 年生胸径遗传力与5 年生的持平。因此,5 年生以后的树高和胸径重复力和遗传力趋于稳定。
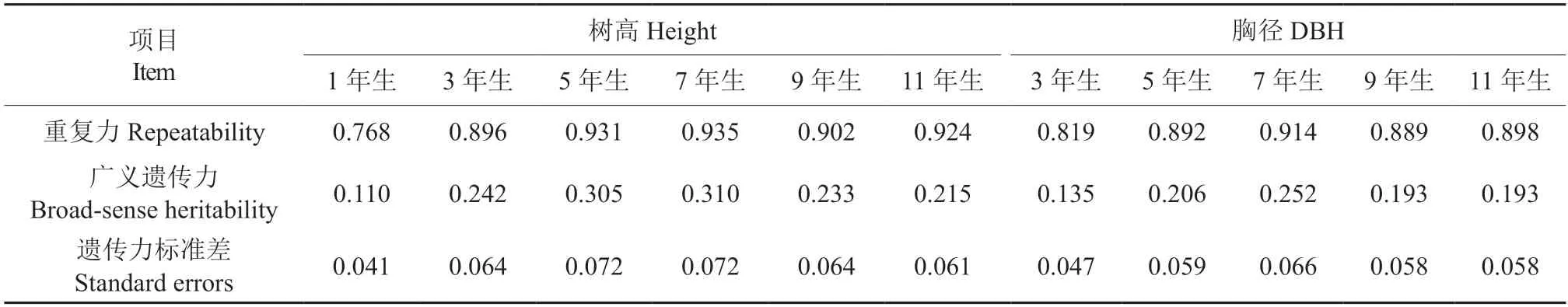
表3 柚木无性系历年树高和胸径生长遗传分析Table 3 Genetic analysis of the growth of height and DBH of teak clones at different ages
2.3 柚木无性系生长性状年-年相关分析
对1~11 年生的树高和胸径进行表型和遗传相关分析,表4 结果表明,遗传相关系数高于表型相关系数,除树高1 年生的外,树高和胸径的其他年份与之后任一年份间的表型和遗传相关关系极显著(P<0.001);树高间、胸径间表型和遗传的相关系数随着两两年份间相距增加而减低;1 年生的树高与其他年份的树高的表型和遗传相关系数最低,分别为0.316~0.488 和0.489~0.675,较其他年份间的相关系数低(表型相关0.701~0.927 和遗传相关0.864~0.996);5 年后的树高和胸径表型和遗传相关系数趋于稳定,11 年生与5 年、7 年和9 年生间的树高表型和遗传相关系数相差不大,分别仅是0.200~0.233和0.027~0.065,但与3 年生树高的表型和遗传相关系数相差分别达到0.299 和0.119;同样,11年生与5 年生、7 年生和9 年生间胸径的表型和遗传相关系数相差不大,分别是0.029~0.143 和0.001~0.058,但与3 年生胸径的表型和遗传相关系数相差较大,分别是0.255 和0.218,因此,说明5 年生可以开始选择无性系。

表4 树高和胸径年-年表型(对角线下)和遗传(对角线上)相关矩阵†Table 4 Correlation matrix of the age-age phenotypic (Lower diagonal) and genetic (Upper diagonal) correlation coefficients of height and DBH
2.4 柚木无性系生长类型划分
以1 年生的树高和地径、3~11 年生的树高、胸径和单株材积进行各年份的主成分分析,1~11年生的第一主成分贡献率为94.50%~98.07%(表5),第一主成分包含了绝大部分信息,以此划分生长类型。本试验第5 年时,林分已郁闭,因此,把郁闭前(1~5 a)划为前期,郁闭后(7~11 a)划为后期,根据表5 对24 个参试无性系(含对照)按25%、50%和25%分别划分为生长快、中和慢3 个生长类型,结果如表6 所示。
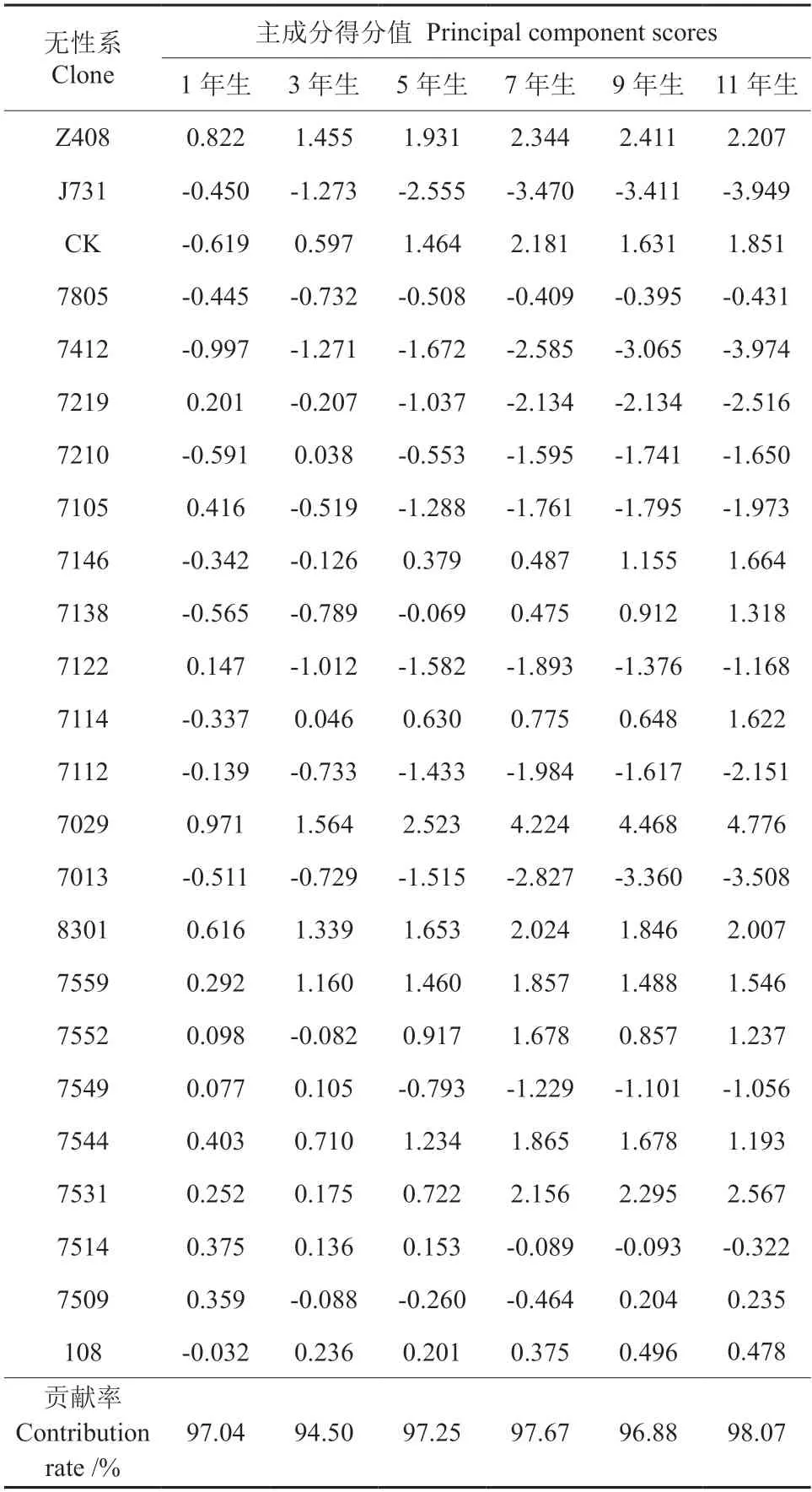
表5 柚木无性系历年生长主成分分析†Table 5 The principal component analysis of the growth of teak clones at different ages

表6 柚木无性系生长类型的划分Table 6 Division of the growth types of teak clones
从表6 可以看出,前期生长快的无性系,后期生长也快,这种生长类型占20.83%;大部分前期生长中等的无性系,后期也保持中等的生长速度,这种类型占37.50%,而仅有1 个无性系前期生长中等,后期生长快,仅占4.17%;前期生长慢的无性系,后期生长也慢,占16.67%,而没有1个是前期生长慢,后来生长变快的无性系。由此可见,前后期生长速率(快、中、慢)保持不变的无性系占75.00%,这结果表明无性系早期选择是可行的。
2.5 柚木无性系早期选择年龄
以表5 第一主成分各个无性系的得分值作为综合评价进行排序,按20%的入选率,选出历年生长前5 位和后5 位的无性系来确定柚木无性系最早选择年龄。以11 年生入选无性系为基准,前3 年入选前20%的无性系有3 个,选准率为60%,5 年以后入选前20%的无性系有4~5 个,选准率保持在80%以上,也就是说,5 年生时可以有80%的准确率预测11 年前20%的无性系生长和排名。其中,有3 个无性系(7029、Z408 和8301)从1年生开始到11 年生,一直保持前5 名的位置,尤其是7029 无性系一直保持第一的位置,而Z408 无性系除在11 年生跌落到第3 位外,其余年份均为第2 位。与前20%入选无性系相比,后20%入选最差无性系,前3 年入选的无性系为2~3 个,选准率为40%~60%,5 a 后的选准率达到80%以上,其中有2 个无性系(J731 和7412)从第3 年开始一直是生长最差的无性系。根据入选前20%或后20%的无性系选准率,5 年生可以作为最早开始选择柚木无性系的年龄,选准率80%,基本上能反映出该试验林11 年生的无性系生长趋势。

表7 柚木无性系不同年龄早期选择结果Table 7 The results of early selection of teak clones at different ages
3 讨论与结论
3.1 讨论
在林木早期选择研究中,生长性状的变异系数被用作确定早期选择年龄的指标之一,把生长变异由剧烈分化趋向稳定的转折年龄作为早期选择年龄[9,34-35]。本研究表明无性系间树高和胸径差异极显著,变异系数随树龄增大而减少,5 a 是变异系数由大到趋向稳定的转折年龄。
广义遗传力随着林木生长发育阶段的年龄变化而变化,并非恒定[36-37],这与本研究是一致的。广义遗传力是随林分年龄有规律的变化,先略升后缓降,并趋向稳定。11 年生树高和胸径的遗传力大小与5 年生的持平,同样,重复力与广义遗传力有相同变化。根据相关分析,11 年生与5 年生间树高和胸径的表型相关和遗传相关极显著,且相关系数与11 年生和3 年生的数值有明显差异。而从生长类型划分来说,前后期生长速率保持不变的无性系占75%,以11 年生选择为基准,3 年、5 年、7 年和9 年生优良无性系选准率分别为60%、80%、100%和80%。由此,可确定柚木无性系早期选择的适宜年龄为5 a,这与杉木无性系的3~4 a[14]和5~6 a[15]的结果一致,但与柚木种源试验[38]结果不一致。后者结果表明4.5 年生与30.5 年生的种源生长性状相关不显著,主要原因可能是试验材料不一致,无性系内个体间不存在遗传差异,仅存在小环境造成的差异,而种源试验既存在种源内个体的遗传差异,又存在小环境间造成的差异,因而可能导致年份间性状生长不一致,相关性也就不显著,4.5 年生柚木优良种源选准率仅33.33%。江西大岗山杉木种源试验也说明了这一点,31 年生种源如在5 和10 a 的选准率18.18%和33.33%[39]。因此,随着早期选择年龄延后,选准率也在提高[40]。
无性系早期选择的目的是尽早选出优良无性系应用于生产造林,缩短选育周期。尽管不同无性系遗传特性不一,随着树龄增长,生长趋势有可能不同,只要早期选出的优良无性系与后期生长选出的无性系基本一致,就可达到本研究的目的。从本研究结果对11 年生优良无性系来说,5年生优良无性系选准率达到80%,说明5 年生作为柚木无性系早期选择年龄是可行的。本研究结果可为柚木优良无性系早期选择提供依据,5 年生结果选出排列前4~5 的优良无性系可用于混系造林,不必等待半个轮伐期或1 个轮伐期的选择结果。同时混系造林也规避了单一无性系选择的误选或单一无性系难以抵抗病虫大面积危害的风险。本研究基于目前的材料,连续观测11 年生柚木无性系的生长,之后的无性系生长与5 年生的相关性有待今后进一步的观测与验证,如因所用的材料或环境不一,结果或许不同。
3.2 结论
柚木无性系前期生长与后期生长保持不变的无性系达到75%,无性系早期选择是可行的。无性系树高和胸径生长变异系数5 年生趋于稳定;以5 年生无性系20%选择强度选择柚木优良无性系,对11 年生相应选择强度的优良无性系选准率达到80%,可初步确定5 年生作为柚木无性系早期选择的年龄。
猜你喜欢
林业科学研究(2023年1期)2023-04-29 15:10:21
花火B(2022年6期)2022-08-23 04:16:39
林业科技情报(2022年4期)2022-04-07 13:39:35
湖南林业科技(2020年2期)2020-09-18 05:36:44
国际木业(2019年2期)2019-09-10 21:52:23
现代农业科技(2017年4期)2017-04-10 00:20:43
北广人物(2017年2期)2017-03-25 04:52:40
作文周刊·小学六年级版(2016年26期)2017-03-06 10:08:56
绿色科技(2017年1期)2017-03-01 10:17:01
广西林业科学(2016年4期)2016-03-16 05:44:55
