
亚热带稻田土壤持续固碳机制研究进展
2022-12-03 06:26:54李宝珍周萍李宇虹李颜魏晓梦陈香碧高威吴金水
华中农业大学学报(自然科学版) 2022年6期
李宝珍,周萍,李宇虹,李颜,魏晓梦,陈香碧,高威,吴金水
中国科学院亚热带农业生态研究所/农业生态系统过程重点实验室/长沙农业环境观测研究站,长沙 410125
土壤碳库是陆地生态系统碳库中最大的储库,全球约1.5×1012~2.5×1012t碳以有机质形态储存于土壤中,是陆地植被碳库储量的2~3 倍,是大气碳储量的2 倍[1],其储量的微小变化,会强烈影响大气CO2浓度从而影响全球气候系统[2]。土壤固碳被认为是目前经济可行和环境友好、能够减缓大气CO2浓度的有效措施[3]。据估计,从1982 年到2006 年的25年来我国农田土壤增加的碳汇相当于2006年全国总CO2排放量的40%,农田土壤固碳为我国贡献了20%的减排份额[4]。
我国水稻种植历史悠久,是世界上稻田面积最大的国家,总面积在3 000万hm2左右,约占全国耕地总面积的27%,其中90%分布在南方亚热带双季稻区[5],在我国的粮食和油料等大宗农产品生产中占据举足轻重的地位。水稻土作为一种独特的人工湿地生态系统,是在长期植稻下人为培育的特殊耕作土壤,其除了具备一般土壤所具有的物理、化学和生物过程之外,因其湿地属性,还具有氧化-还原交替过程及由此诱发的特殊化学与生物化学过程(如铁、硫等元素的氧化-还原、有机质的嫌气分解和CH4的产生等),这决定了其碳与养分循环过程区别于其他土壤。而且,稻田具有固碳的生态功能,是重要的碳汇,且是全球N2O 和CH4的主要排放源之一。因此,水稻土壤是研究碳固定和温室气体减排的关键对象之一[6-7],土壤固碳对于加快我国农业双碳目标和农业绿色发展具有重要的意义,也一直是反映我国土壤学的国际地位的重要标志。
近30 年来,我国亚热带水稻土壤有机碳含量持续增加,显著高于旱地和林地等其他类型土壤,表明亚热带稻田生态系统具有重要固碳作用[8-9],相关学者在亚热带水稻土壤的碳循环过程以及固碳减排潜力和机制方面进行了大量的研究,推动了全球碳循环的生物地球化学理论发展。基于此,本文就亚热带水稻土壤有机碳储量的时空分布与演变规律及碳循环生物地球化学过程等前沿性的重要基础科学问题进行了探讨,旨在为稻田土壤培肥和地力提升,以及我国农业双碳目标和农业绿色发展提供科学依据。
1 亚热带稻田土壤有机碳储量的时空分布与演变规律
我国亚热带区稻田土壤有机碳储量与固碳效应明显高于旱地土壤,具备较强的固碳潜力[9-11]。研究表明,1980—2002年的20多年时间里,我国农田土壤增加的有机碳中有74%来源于水稻土耕层有机碳的积累[12],且以双季稻区>水旱轮作区>单季稻区[13]。我国南方亚热带区域的稻田土壤约60%是以双季稻种植为主,故而我国农田土壤固碳效率以南方较高[13-14]。水稻土壤碳氮储量增加主要有3 个原因:(1)水稻秸秆和根系分泌物等对土壤有机碳的输入量大于大多数旱地谷物;(2)植物残体和有机质在稻田缺氧条件下的分解较慢;(3)铁氧化物在有机质的稳定中更为重要(图1)[11]。因此,亚热带稻田土壤有机碳的储量动态对于准确评估我国区域碳“源汇”功能具有重要意义。

图1 水田和旱地土壤碳储量差异的影响因素概念框架图[11]Fig.1 Mechanisms leading to greater C and N stocks in paddy soils compared with those in upland soils[11]
土壤碳库中的有机碳库集中分布于土层1 m 左右深度,而长期水稻种植条件下,1 m 深土壤剖面中约有50%以上的有机碳储存在20 cm 以下的底土层,极易受到人为干扰和环境变化的影响[15-16]。Pan等[8]利用第二次全国土壤普查,估算了土壤有机碳密度在12~226 t/hm2,1 m 深土壤剖面中有机碳库为13 亿t,其中耕层为8.5 亿t,犁底层为4.5 亿t。稻田表土每年有机碳的积累速度为170~178 kg/hm2,而底土每年有机碳则以29~84 kg/hm2的速度损失[17]。但是,施肥条件下稻田土壤有机碳积累不仅局限于0~20 cm 表层,更有向犁底层以下迁移的趋势,并将其原因推测为是溶解性有机碳(DOC)的向下迁移并与有机矿物的相互作用而稳定的结果[18-19]。因此,为充分了解区域稻田土壤有机碳库对气候变化和人类活动的响应,有必要对涵盖底土的土壤剖面有机碳库的过去和未来的演变规律进行有效评估。
土壤有机碳储量受气候、水文和地理环境、土壤条件、植被与土地利用等要素的影响[20-21]。最佳的氮肥施用可能显著影响着土壤的固碳效率,化学氮肥在农田土壤表层的每年平均固碳潜力为(−20±210)kg/hm2[22-23];施用富含有机质的农家肥可提高土壤有机碳储量,每年平均固碳潜力为(292±132)kg/hm2[22];在表层土壤(0~20/30 cm),免耕或少耕措施分别使表层土壤每年固定(343±167)、(324 ±138)kg/hm2[24-25]。国际上也建立了许多土壤有机碳过程模拟模型,可用于评估和预测区域土壤有机碳的长期变化[26-27]。土壤有机碳库分为活性碳库、慢性碳库和惰性碳库,其中活性炭库平均滞留时间为2~4 a;慢性碳库平均滞留时间为20~50 a;惰性碳库平均滞留时间可高达400~2 000 a[28]。土壤活性碳库具有稳定性差、易矿化、性高的特点,对环境的反应敏感,成为反映土壤有机碳库的重要指标[29]。
2 稻田土壤有机碳的输入与作用机制
稻田土壤突出的碳固持能力主要是由于水稻光合碳的输入、土壤自养微生物固碳及淹水限制了微生物活性、抑制植物残体微生物分解过程,促进以植物残体直接积累[30-32](图2)。稻田土壤固持的有机碳中植物残体占33%~54%(是旱地土壤的3.3 倍)、微生物占28%~36%[33]。13C 自然丰度能够用于揭示团聚体与土壤有机质(SOM)密度组分之间的碳稳定途径[34-35],稻田土壤团聚体大小类别之间的碳流动路径遵循从宏观到微观团聚体的趋势,光合碳13C主要分布在最小的粉砂黏土矿物颗粒上[36];与轻组分相比,重组分的13C 消耗更多,这表明分子质量低的有机物在黏土表面上快速稳定,并且浸入铁(氢)氧化物沉淀[37]。
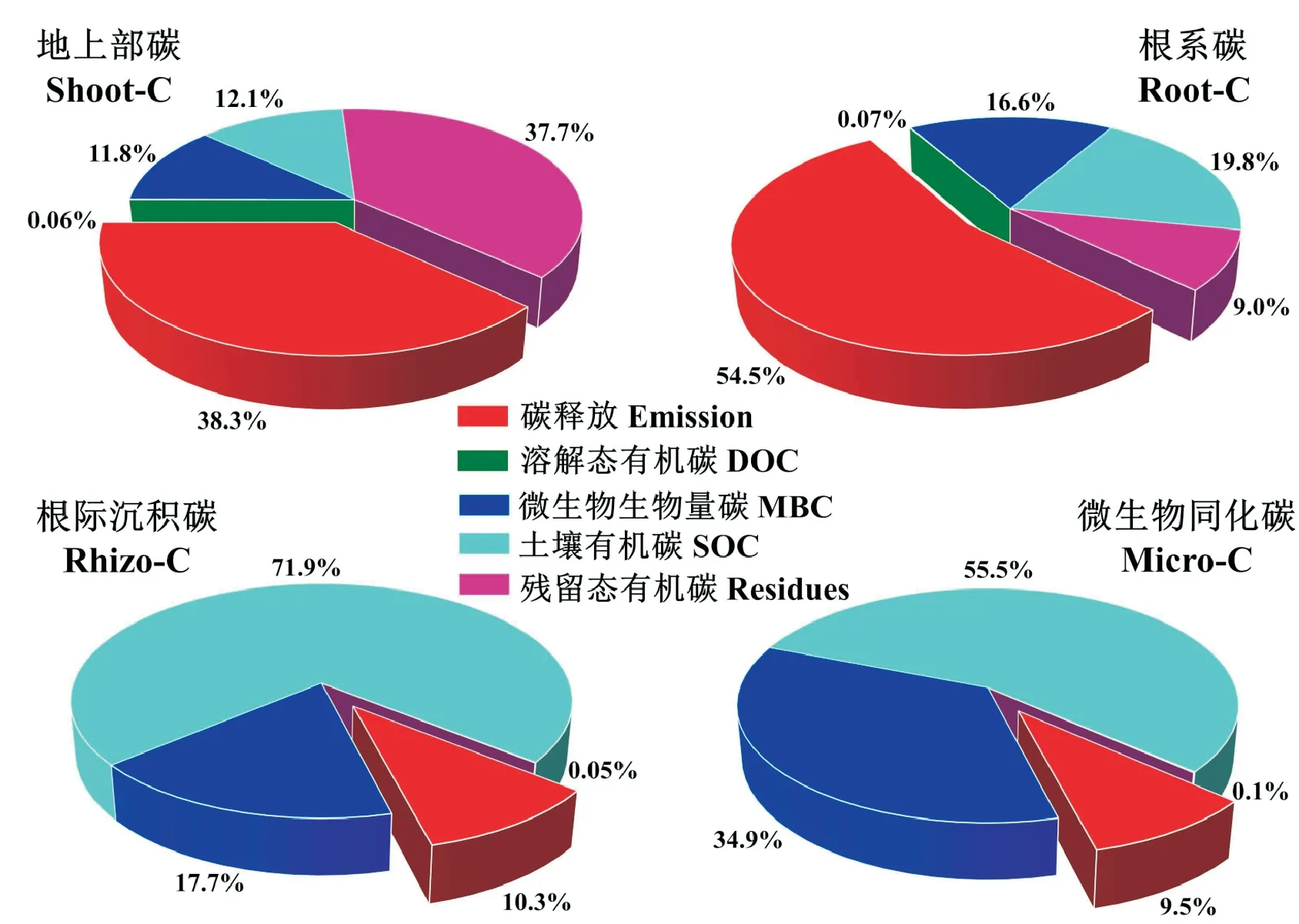
图2 水稻秸秆、根系、根际沉积碳和微生物同化碳在不同碳库中的分配[32]Fig.2 Distribution of different C substrates(shoot-C,root-C,rhizosphere-C,microbial-C)in different C pools[32]
水稻光合碳是作物光合作用固定的大气二氧化碳经过凋落物、根系和根系分泌物等根际沉积形式进入土壤的碳,在农田碳循环中起着重要的作用,是水稻土壤高碳库的重要碳汇来源之一[38-39]。研究报道,水稻光合同化碳向土壤的输入量约占根系碳量的34%,据估算一季水稻的地下碳输入量为630~1 080 kg/hm2,其中包括160~330 kg/hm2的水稻光合碳[38]。根系沉积的碳表现出较强的根际效应[40],水稻根际DOC 随着根系分泌物的增加而升高,且根际微生物量碳也与DOC 显著相关[41-42]。水稻光合碳在有机质含量低的土壤中60%以呼吸形式消耗掉,而在有机质含量高的土壤中则具有较低的微生物碳周转率,对土壤有机碳库贡献较高[43]。根际沉积碳稳定性受作物生长时期和水肥管理的影响。施氮增加了根际沉积碳量以及光合新碳(13C)向根际土的传输量[44-45],使水稻光合碳在不同时期的输入提高34%~381%,其中进入土壤中的光合碳提高0.9~1.9 倍[42,46]。而且,施氮与干湿交替促进光合碳向土壤的输入及其稳定性,从而提高土壤的固碳能力[45]。但是,施氮也提高了根系的分泌物,根系分泌物输入量的增加也加速了根际微生物对新鲜碳源周转速率[42,46]。
3 稻田土壤微生物固碳功能与机制
农田土壤中自养细菌分布广泛,能够通过多种生物固碳途径固定大气中的CO2,其具有巨大的固碳潜力[47-49]。据估算,水稻土的微生物年碳同化量达100~450 kg/hm2,占土壤有机碳含量的比例达0.9%~4.1%,即对土壤有机碳年输入量相当于土壤有机碳总量的0.9%~4.1%,与一般生态系统中植物对土壤的年输入量几乎相当[50]。另有研究报道,稻田14C-CO2的固定量可达0.074~0.175 g/kg[51]。水稻土固碳细菌、蓝细菌和藻类的CO2光合细菌发挥同化CO2的主要作用,揭示了土壤微生物固碳速率与土壤固碳细菌cbbL基因丰度及RubisCO 固碳酶活性显著相关,固碳酶RubisCO 活性可作为土壤微生物碳同化速率的估算指标[49-50]。
土壤固碳微生物和固碳酶的活性受长期施肥的影响,从而显著影响稻田土的固碳速率[50,52]。长期单施化肥或秸秆还田配施化肥可以提高RubisCO 酶活性,且以秸秆还田配施化肥对RubisCO 酶活性的影响最大,从而可以提高稻田土壤的固碳效率[50]。但亦有不同的发现,长期单施化肥使稻田RubisCO酶活性降低[53]。这是因为pH 可以直接影响Rubis⁃CO 酶活性,长期施用化肥导致土壤酸化,pH 下降导致土壤固碳效率下降[50,53-54]。光照亦是土壤的固碳能力的一个重要因素,研究发现土壤表层1 cm 中RubisCO 活性更高,其固碳量也显著提高(高达(1.180±0.105)g/kg)[49,54-55]。土壤质地也会影响土壤固碳能力,黏土具有更高的固碳能力[56]。施磷处理显著提高了根际微团聚体和粉砂黏土颗粒上13C的分布,这有利于水稻土对碳的固定和储存[36]。真菌和革兰氏阴性菌是重要的光合碳的根际同化者,负责将根际碳周转到水稻土壤中的其他的微生物群落中利用[57-58]。添加可溶性有机物料与氮肥联合施用,导致了革兰氏阳性菌和真菌丰度的降低,降低了光合碳在有机质和微生物量中的分配[59]。
4 稻田土壤有机碳矿化过程与调控机制
土壤碳矿化是有机碳分解转化成CO2的过程,与土壤活性有机碳库关系密切,因此,土壤有机碳矿化释放CO2的数量与强度是评价环境因素对土壤有机碳分解转化的重要依据[60]。土壤有机碳矿化与土壤结构、土壤团聚体以及与之相关的物理化学性状密切相关,从而调控着土壤的碳源利用和微生物激发效应,并进一步影响土壤有机碳的积累[61-62]。稻田土壤干湿交替的水分管理方式导致了其特殊的物理、化学、生物环境条件,影响着土壤有机碳矿化过程。
稻田土壤微生物生物量通常数倍于旱地[63-64],然而水稻土有机碳矿化率普遍低于旱地土壤,而且新鲜有机碳对土壤本身有机碳的矿化无激发效应或存在负激发效应,即对有机碳矿化存在“阻滞效应”[63,65]。旱地土壤中,氧气的可利用率高,好氧微生物活性较强,促进细菌量的增加,导致土壤有机碳矿化速率较快;与旱地相比,水稻土氧气可利用率较低,有机碳的氧化热动力学上受限[66],好氧微生物活性较弱,因此,微生物生物量周转比旱地慢很多[64],生物酶(如多酚氧化酶、水解酶)的分泌也受限[67],淹水稻田土壤中真菌/细菌比值的增加也会抑制土壤有机碳的矿化速率[68-69]。土壤中微生物量碳(MBC)氮磷计量比及土壤酶活性计量比通过直接或间接地影响土壤有机碳的矿化[40]。在低、中等碳(50%~300%MBC)输入下,水稻土有机碳矿化的激发效应产生是由于微生物需要通过加速本身SOM 的分解获取足够N 素;但是高碳输入下(300%~500%MBC),土壤微生物无法通过单纯加快SOM(较难利用)分解而迅速获得足够的N 素,转而通过死亡微生物量的再利用而解决严重的N 匮乏[70]。另外,在中低等碳输入下,氮硫磷(NPS)养分输入使资源获取策略微生物(A-策略)占据主导地位,增加了SOC 的矿化,且正激发效应的强度随着NPS 养分输入的增加而增加;在高量活性碳输入下,NPS 养分输入有利于高产策略微生物(Y-策略)生长,负激发效应的强度增加(图3)[71]。
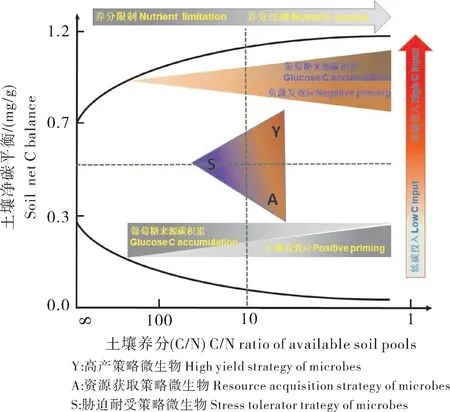
图3 微生物生活策略对激发效应和土壤碳平衡的化学计量调节机制的概念图[71]Fig.3 Adjustment of microbial life strategies to the amount and stoichiometry of labile resources,and outcomes in terms of soil C processing and net C balance[71]
厌氧条件下有机碳分解不彻底,会伴随有机酸、乙醇等中间产物的产生,有机碳矿化的最终产物包括CH4和CO2。厌氧条件下,水稻土中有机碳矿化和甲烷排放在低温下(<15 ℃)对温度变化更敏感。CO2排放的Q10值(温度敏感性指标)在5~15 ℃时比在25~35 ℃时高1.1~3.4 倍;CH4排放的Q10值在5~15 ℃时比在25~35 ℃时高2.8~13.5 倍[72]。氧气充足时,土壤有机碳可利用性是有机碳矿化的限速因子;氧气受限或缺乏时,微生物生物量、有机碳形态组成和可利用电子受体共同限制有机碳矿化[73]。亚热带水稻土通常含有丰富的铁铝氧化物,可以对有机碳形成物理保护,干湿交替土壤具有交替的好氧-厌氧循环,铁(羟基)氧化物可以与有机碳发生吸附或共沉淀,降低其与微生物的可接触性,从而限制微生物对有机碳的分解与矿化[63,74]。另外,铁氧化物还可以作为电子受体,促进厌氧条件下水稻土有机碳矿化为CO2,抑制有机碳矿化为甲烷;对于乙酸的矿化,针铁矿的电子受体作用大于吸附作用,水铁矿的吸附作用大于电子受体作用[75]。
5 “双碳”背景下稻田土壤固碳研究展望
尽管过去几十年间针对稻田土壤碳循环的关键生物地球化学过程已进行了广泛研究,但相关过程的内在作用机制及生态环境效应认识仍存在许多不足。面对全球气候变暖、温度逐渐升高的巨大挑战以及国家粮食安全和生态环境建设的双重要求,作为世界第二大CO2排放国,进一步增强稻田土壤的碳汇功能,是推进水稻绿色低碳高质量发展的重要内涵,也是助力国家碳达峰碳中和的“双碳”战略的重要手段。现提出如下展望:
系统揭示稻田生态系统中土壤机碳累积和转化的作用机制,助力提升农田土壤肥力。针对亚热带稻作系统生产力提升、减投增效和农产品安全的国家需求,研究亚热带稻作系统水稻土有机碳积累、持续生产力和微生物过程的内在机制,突破水稻土肥力提升的科技瓶颈;借助生态化学计量学新工具,深入研究“水稻-土壤-微生物”系统的根际沉积碳周转与稳定过程,揭示驱动稻田生态系统中土壤有机碳持续累积的内在调控机制,为稻田土壤培肥和地力提升提供科学依据。
构建高精度的区域土壤有机碳库模拟模型及储量估算方法。由于时空不平衡、监测方法不一致等因素的制约,我国稻田碳汇功能的评估还存在较大不确定性。依据已有研究数据和成果支撑,结合野外调查与遥感同步反演,获取系统全面的土壤监测数据,通过分析土壤有机碳密度随深度变化的普遍性规律,剖析土壤剖面有机碳密度与表土碳密度和土壤深度的函数关系,构建基于表土碳密度估算土壤剖面有机碳库的模拟模型,优化土壤剖面碳库估算方法,从而提高全球或区域土壤有机碳储量的估算精度。同时,随着人工智能模拟技术越来越成熟,神经网络模拟方法需要逐步引入到生态学和表层地球科学等领域,将在区域土壤有机碳模拟与储量估算方面展现很好的前景。
猜你喜欢
中国土壤与肥料(2021年5期)2021-12-02 01:06:26
绿色科技(2020年20期)2020-11-20 01:56:34
天然产物研究与开发(2018年3期)2018-05-07 06:38:35
江苏农业科学(2017年6期)2017-05-11 14:35:37
林业勘查设计(2017年1期)2017-03-11 16:49:57
中国蔬菜(2016年8期)2017-01-15 14:23:38
浙江农业学报(2016年7期)2016-06-15 20:30:08
中国农业文摘-农业工程(2016年5期)2016-04-12 05:38:16
现代农业(2016年4期)2016-02-28 18:42:07
环境科技(2015年2期)2015-11-08 12:11:30
