
基于海马体的面向未知复杂环境类脑导航技术综述
2022-12-01 12:38陈雨荻熊智刘建业杨闯晁丽君彭杨
兵工学报 2022年11期
陈雨荻, 熊智,2, 刘建业,2, 杨闯, 晁丽君, 彭杨
(1.南京航空航天大学 自动化学院 导航研究中心, 江苏 南京 211106;2.先进飞行器导航、控制与健康管理工信部重点实验室, 江苏 南京 211106;3.上海航天控制技术研究所, 上海 201109)
0 引言
近年来,机器人、无人车、无人机等无人移动平台在民用领域和军事领域被广泛推广[1],在作战领域发挥出巨大的作用。在执行任务时,无人移动平台的自主导航和运动控制能力尤为重要,因此搭载的导航系统占据重要地位,面对未知复杂的环境,现阶段的导航系统面临着诸多挑战,需要进一步提升智能导航能力。
众所周知,自然界中许多动物有较强的导航本领,不需要高精度的传感器就能在未知复杂环境中生存,通过回忆之前的经验可较好地完成导航任务。随着人工智能的发展,智能自主导航技术也有了诸多研究方向。类脑导航作为人工智能(AI)技术的研究方向之一,其内容涉及了脑科学和控制科学等多个交叉学科内容。现阶段类脑自主导航技术的概念还未统一,其基本内涵为通过多种传感器感知环境信息,利用具有与大脑功能相似的智能技术形成仿生的学习、记忆和认知能力,能够融合多种信息构建认知地图并实现智能路径规划和导航[2],与其他导航方式相比,类脑自主导航技术具有不依赖卫星定位、低能耗等特点,为未知复杂环境下自主导航提供了有效的技术途径[3]。本文将介绍类脑自主导航技术的最新研究进展,沿类脑自主导航技术的研究路线,即理论方法、模型构建以及应用实现三方面构成文章组织结构,同时综述与类脑自主导航发展相关的代表研究成果。
1 基于海马体的类脑自主导航技术机制
大脑导航机理提供构建“类脑”模型的方法和结构。由于类脑导航技术源于哺乳动物大脑对环境的认知方式,与仿生导航技术相比,类脑导航技术不止步于模拟生物导航行为,而是深入探索产生导航行为的来源,在模型构建方面会侧重深层次模拟大脑组织结构以及神经细胞活动等。哺乳动物大脑中参与环境认知和导航的组织主要为海马体(HF),其中内嗅皮层- 海马区空间表征机理为类脑导航系统的构建提供理论基础,为了方便读者理解类脑模型构建和应用,本节归纳类脑导航技术研究所需的大脑机理。
1.1 内嗅皮层- 海马区导航信息传递回路
哺乳动物大脑的海马体多个区域参与记忆、认知和导航活动[4],其组织之间的连接与类脑导航系统模块通信方式形成映射。综合Witter和Van的研究内容[5-6],从宏观角度分析,参与导航的海马体可分为齿状回 (DG)、海马区[7-8]、下托综合体[9]和内嗅皮层[10]。其中,下托综合体分为下托(Sub)、前下托(PrS)、傍下托(PaS)和后下托(PoS)。海马区进一步细化为海马角(CA)、CA1~CA4,CA1与CA2通常可定义为海马区。
近年来,随着对海马体研究的逐渐深入,研究学者揭示了海马体整合信息的通信路径。外界环境信息由嗅后皮层(POR)及嗅周皮层(PER)传入内嗅皮层,内嗅皮层分为内侧内嗅皮层(MEC)和外侧内嗅皮层(LEC),内嗅皮层共有Ⅵ层。内嗅皮层的第Ⅱ层将信息传递给齿状回和CA3,CA3有大量的循环连接,产生了自联想记忆的假设。外侧内嗅皮层第Ⅲ层将信息主要传送到Sub,内侧内嗅皮层第Ⅲ层将信息传递给CA1区和Sub,CA1区和下托将信息传回内嗅皮层的深层,由内嗅皮层深层到浅层,再到齿状回构成一个回路[11]。图1为海马体结构示意图,图2为信息传递回路示意图。
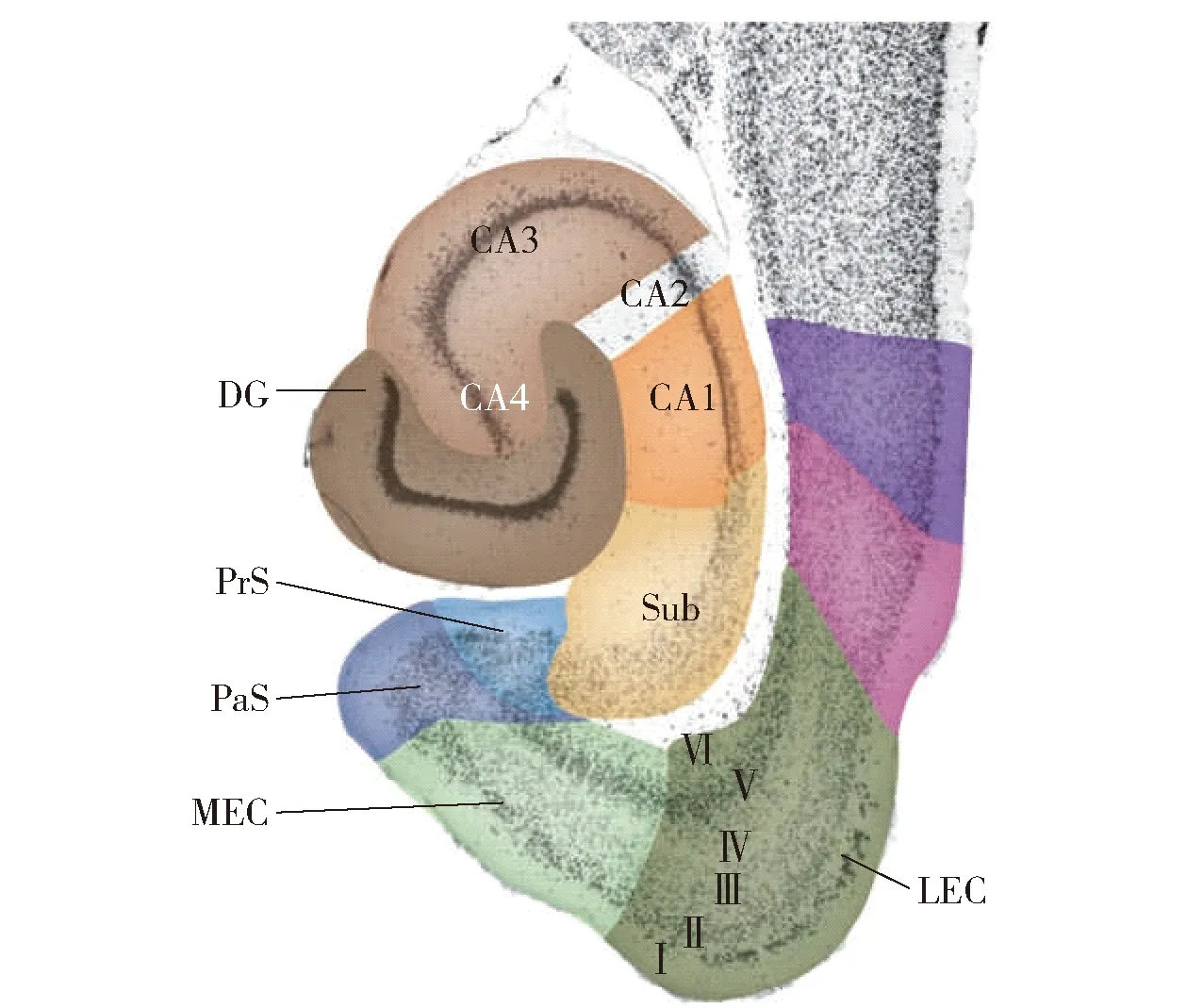
图1 海马体[5-6]
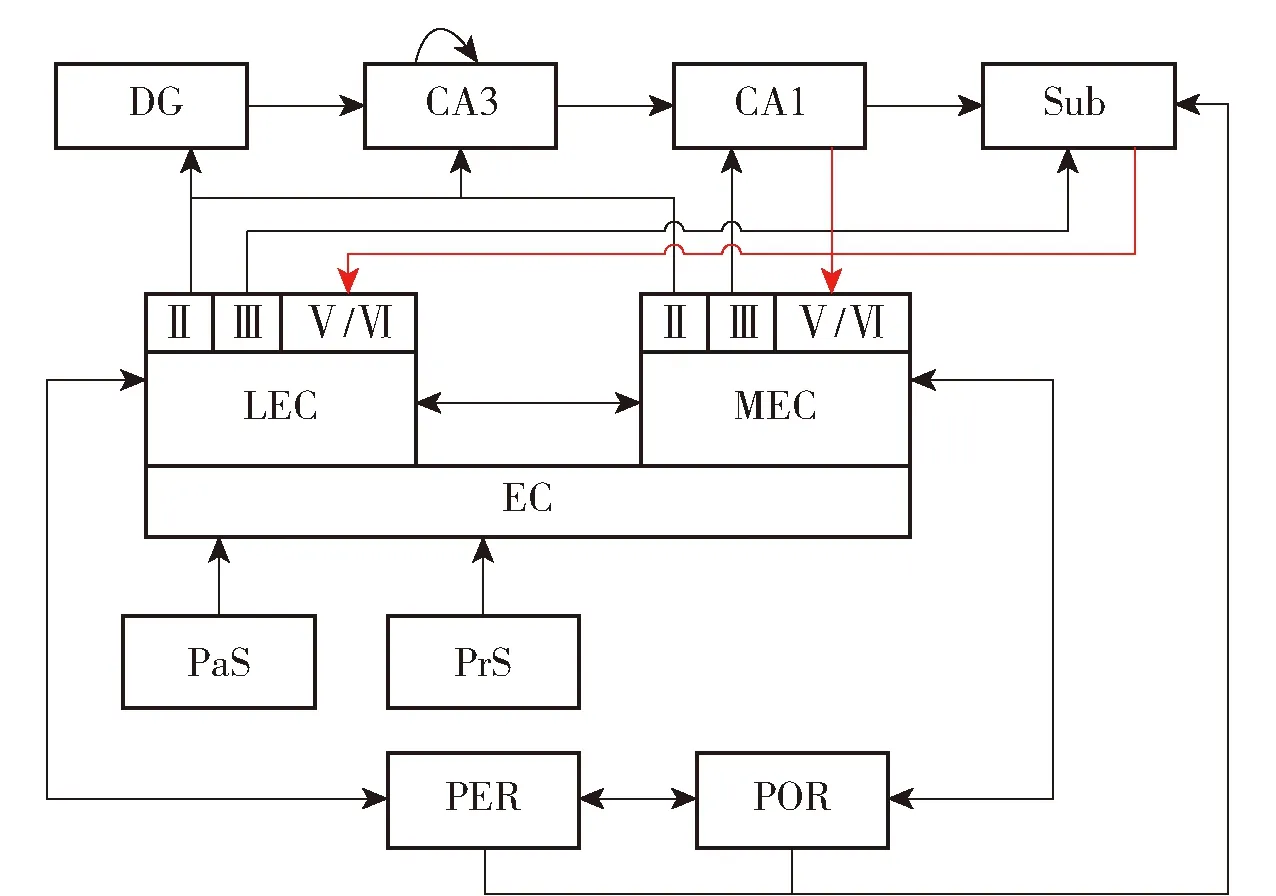
图2 内嗅皮层- 海马区信息传递回路[11]
1.2 空间表征细胞特性
海马体中的空间表征细胞编码输入信息产生放电,其放电形态用于表征环境,由此可以根据放电特性构建模型。现阶段主要研究的空间表征细胞为位置细胞、头朝向细胞以及网格细胞。
1.2.1 位置细胞
O’keefe等在鼠脑CA3和CA1中发现了位置细胞(place cells)[12-13]。位置细胞放电活动与动物所处空间位置有很强的相关性,当大鼠处于特定位置区域时,位置细胞才会产生动作电位[14-15]。位置细胞在环境空间中产生放电的范围称为放电野或位置野[16],根据位置细胞对空间中特定位置产生放电这一特性构建位置细胞模型。其放电形态如图3所示。位置细胞整合内源信息和外源信息进行位置更新[17]。由本体感受的自运动和前庭感觉信息称为内源信息(例如角速度、速度等),基于视觉、听觉、嗅觉等进行外界感知的称为外源信息。
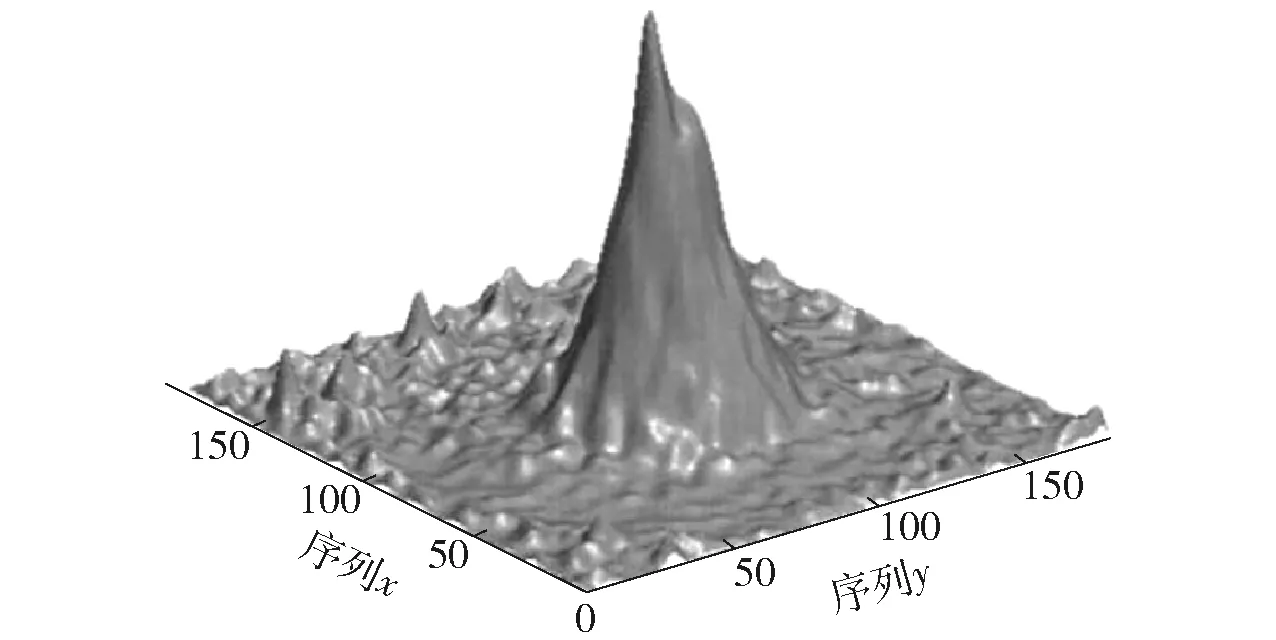
图3 位置细胞放电[14-15]
1.2.2 头朝向细胞
头朝向细胞通过整合内源信息和外源信息中的角速度或者方向角度对当前方位进行估计,起到了“罗盘”的作用。头朝向细胞主要位于鼠脑的PrS、PoS等部位[18],随着Kyriacou等[19]进一步研究发现头朝向细胞具有高斯分布的放电规律,当大鼠头部指向某一特定方向时,相应头朝向细胞会产生最大动作电位,此方向为头朝向细胞放电的最优方向,如图4中头朝向细胞的最优方向为90°。
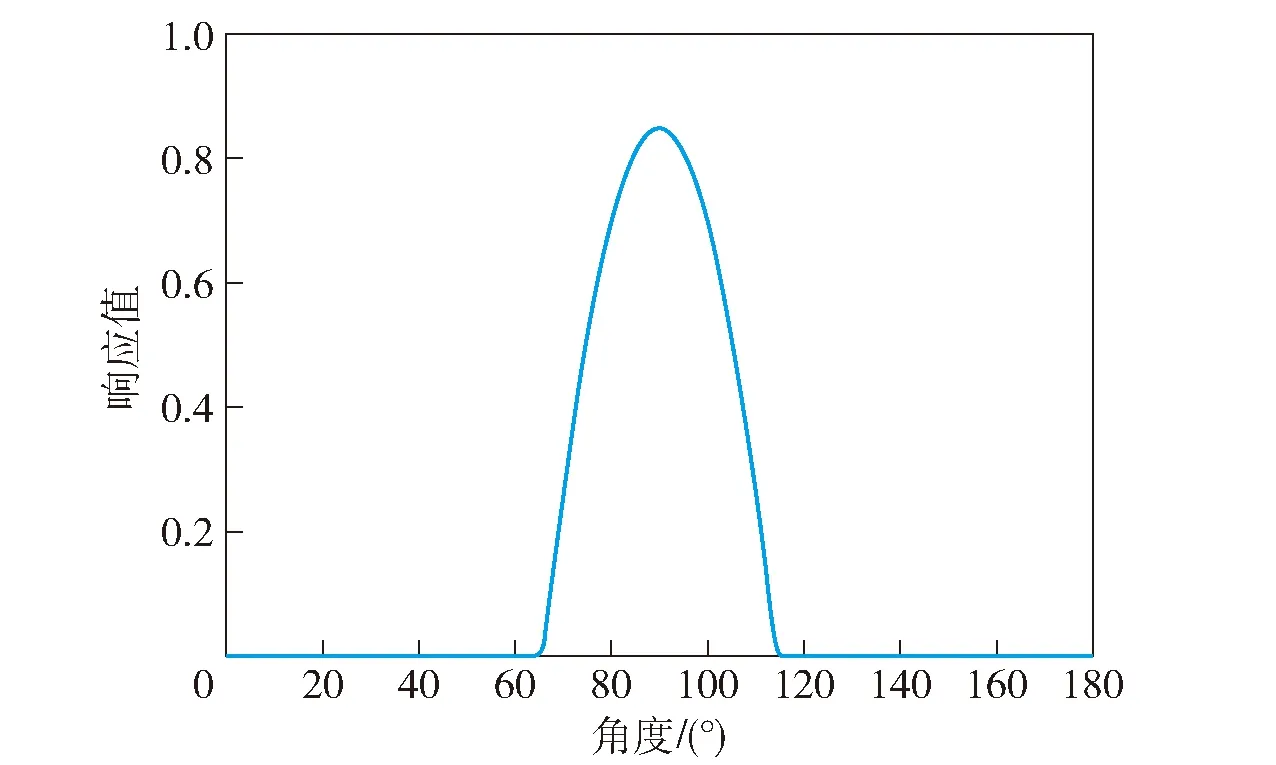
图4 头朝向细胞放电
1.2.3 网格细胞
网格细胞作为“路径积分器”位于大鼠内嗅皮层的第2层中[20]。当大鼠自由活动时,网格细胞在空间位置发生重复性规律放电,放电野在空间环境中形成六边形结构[11-21],如图5所示。在空间环境中形成六边形放电特征成为网格细胞模型模拟的放电特性之一。

图5 网格细胞放电[20]
之后,Fyhn等[22]提出了网格细胞与位置细胞形成的放电野都与空间中的位置相对应的理论,在活动区域中,位置细胞模型需要对应一个特定区域产生放电,网格细胞模型可在多个区域形成放电野。在未知环境下,网格细胞快速形成六边形放电野并保持稳定,同时将外源信息和自运动信息作为信息输入整合运动路径。
另外,网格细胞放电野具有的空间特征在路径综合过程中发挥着重要作用。网格细胞放电野具有3个空间特征,分别为网格尺度、网格方向和网格相位,如图6所示,其中ra和rb表示不同网格尺度的放电野半径,λa和λb表示相同网格尺度放电野之间的距离。网格尺度描述了放电野之间的距离和放电野的大小,网格方向描述放电野连线之间的倾斜程度,网格相位描述相对参考点的位置[23-26]。
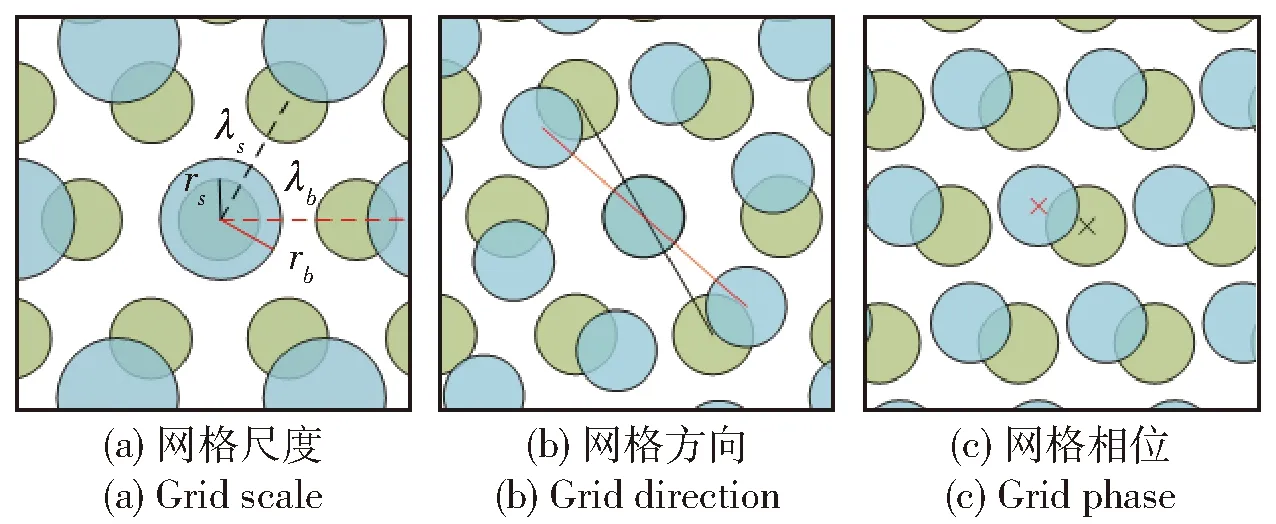
图6 网格细胞空间特征(×表示网格放电野中心点所在相位)
网格细胞的网格尺度大小与表征的空间环境范围相关,多尺度网格细胞模型在小尺度空间(2 m×2 m)范围有较好的表征效果,但在大尺度范围(km及以上)的适用性较差[27],Yang等[28]提出了利用余数系统解码多尺度网格细胞模型的方法,该方法能够使多尺度网格细胞模型适应在较大空间范围中对速度和方位信息的整合。
1.2.4 其他空间表征细胞
随着对内嗅皮层- 海马区空间表征机理的深入研究,除了位置细胞、头朝向细胞和网格细胞,还存在其他功能的空间表征细胞。例如,边界细胞或者称为边界矢量细胞以及条纹细胞和动作细胞等。
边界细胞具有在临近位置细胞群表征空间边界的放电特性[29],之后Lever等[30]发现了边界细胞具有描述空间范围的作用。经研究发现,通过整合具有不同特性的条纹细胞可以形成六边形网格放电野,由此为网格细胞的构建提供理论参考[31]。此外,Dolle等[32]发现大脑中还存在根据环境变化、目标方位以及任务需求产生行为决策响应的动作细胞,该研究为类脑路径规划方法提供了新的技术策略。
2 类脑自主导航系统结构和空间表征细胞模型
类脑自主导航系统模型作为沟通动物大脑导航机制和类脑自主导航系统工程应用的“桥梁”,其系统模型参照大脑导航机制构建,即内嗅皮层- 海马区导航信息传递回路、空间表征细胞之间的信息交互方式以及空间表征细胞放电特性。类脑自主导航系统的构建由单个细胞模型开始,之后以Barrera等[33]的研究为例,开始了网格细胞和头朝向细胞等的集成化模型研究,随后逐渐形成较为完善的类脑自主导航系统模型框架结构。
2.1 类脑自主导航系统整体结构模型
在类脑导航系统结构研究方面,一方面可以从嗅皮层- 海马区整体功能层面建立系统模型,例如Mhatre等[34]提出的自组织内嗅皮层- 海马区计算架构。另一方面,可以将空间表征细胞模型作为模块单元,然后组合模块形成类脑自主导航系统结构,例如Edvardsen等[35]基于网格细胞、位置细胞和边界细胞构建类脑导航系统模型,该系统中结合网格细胞的矢量导航机制以及边界细胞和位置细胞的调节机制实现局部避障功能,并可支持复杂环境中的导航。
图7中展示了较为全面的类脑导航系统结构,该模型由Tejera等[36]构建,在系统中外源信息(例如视觉)和自运动信息作为输入,经过头朝向细胞、网格细胞和位置细胞等空间表征细胞的信息整合和相互作用形成认知地图,进而通过Actor-Critic[37]架构的强化学习奖励机制产生运动决策行为[38]。
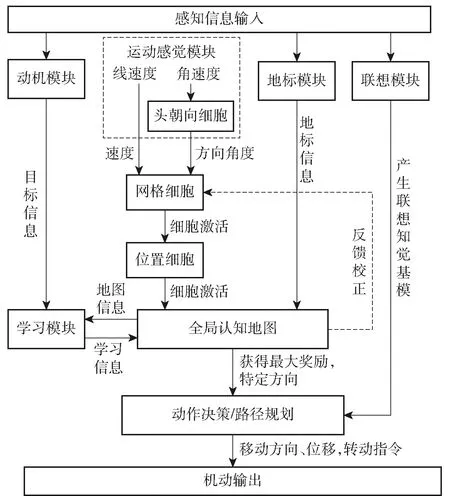
图7 类脑导航系统整体结构模型[36]
2.2 空间表征细胞模型构建方法
空间表征细胞模型作为类脑导航系统的组成单元,其通过编码运动信息模拟神经细胞放电形态,从而表征环境位置变化。现阶段构建空间表征细胞模型可基于吸引子神经网络、振荡干涉原理和脉冲神经网络。
2.2.1 吸引子神经网络模型
吸引子神经网络模型的类脑特性表现为模拟CA3区神经元储存大脑皮层的输入信息和记忆能力。大脑中的神经元之间的突触的连接是可塑的,当最初放电的神经元触发其他神经元时形成记忆的整个形态,此时网络会趋向于一个稳定的状态,具有唯一性、持续性、集成性和漂移校正等特性。
2.2.1.1 吸引子
基于大脑神经特性构建动力学系统,Marr等[39]最早提出了吸引子概念,动力学系统中存在一组稳定的状态,若在输入中施加轻微干扰则会回到这个状态,这些稳定状态即为吸引子,这些吸引子也作为系统中的“记忆”。吸引子被用来描述状态随时间变化的动力学系统的稳定性,其表示一组稳定状态的集合。刘建业等[40]的研究中表明,当系统受到偏移或噪声干扰时,这些代表稳定状态的吸引子会将相邻的状态吸引到该状态,从而保持系统的相对稳定性,这种稳定性体现了吸引子动力学模型与大脑神经网络具有的相关性和一致性。基于上述特性,空间表征细胞通常基于吸引子神经网络进行模型构建。
吸引子按结构分为点吸引子、极限环和连续吸引子等。Wu等[41]的研究证实了连续吸引子神经网络(CANNs)可用于神经系统细胞建模。连续吸引子最简单的结构为一维线性吸引子和环状吸引子。线性吸引子可以表示具有最小值和最大值的变量,例如表示水平方向眼睛位置。环状吸引子适合于表示无边界值的变量,例如头部方向。一维线性和环状吸引子的二维延伸称为平面吸引子,平面吸引子适用于表示二维变量。不同类型吸引子并不相互排斥,可以将多种类型的吸引子嵌入同一网络中。
2.2.1.2 基于吸引子神经网络的空间表征细胞模型
头朝向细胞连接强度的不对称性会使波包沿着圆环移动,波包移动的方向代表头部的朝向,当方向不发生改变时,头朝向细胞连接强度不会发生改变[42]。Stringer等[43]提出了头朝向细胞的自组织连续吸引子神经网络模型如图8所示,每个方向偏好细胞与周围细胞之间存在不同的连接强度,连接强度是它们偏好方向角度差的函数,兴奋性连接的相互作用形成的波包代表当前的头朝向。之后Stringer等[44]提出了基于自组织连续吸引子神经网络的位置细胞模型。
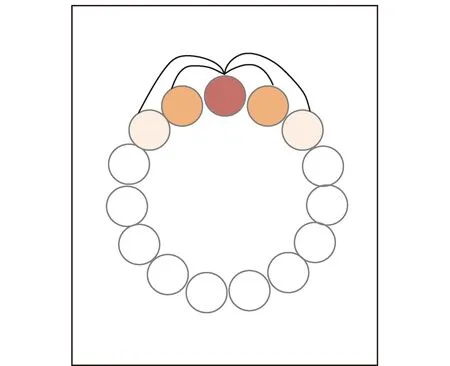
图8 一维环状吸引子[43]
多层连续吸引子的头朝向细胞模型,如图9所示[45],多层连续吸引子模型可扩展空间的表达信息,增加对环境高度的认知,多层头朝向细胞在连续吸引子神经网络中的表达可表示为二维矩阵形式,模型四周为包络连接,实现周期性表达头部朝向和高度信息。

图9 多层连续吸引子[45]
网格细胞模型可建立为二维环状吸引子模型,由于大脑中细胞的活动状态与周围细胞的递归连接强度和输入密切相关,在吸引子网格细胞模型中,每个吸引子代表的是空间环境中的某一位置,神经网络通过吸引子的兴奋和抑制信号来对整个环境进行编码,通过连接神经面板周围的神经元形成环状连续吸引子模型,实现连续的周期性环境信息编码,如图10[46]所示。Burak等[47]提出了速度编码的连续吸引子网络模型,该模型可以在10~100 m,1~10 min 内准确整合速度从而更新位置。为了扩展对三维空间的认知,Yu等[45]将网格细胞模型进行扩展,形成三维连续吸引子模型,该模型可用于三维空间位置积分,如图11所示。
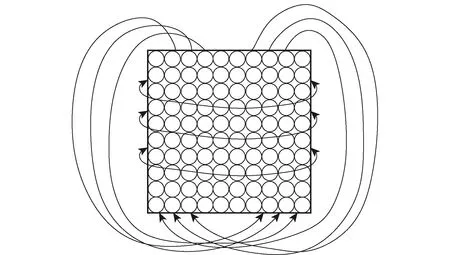
图10 二维环状吸引子[46]
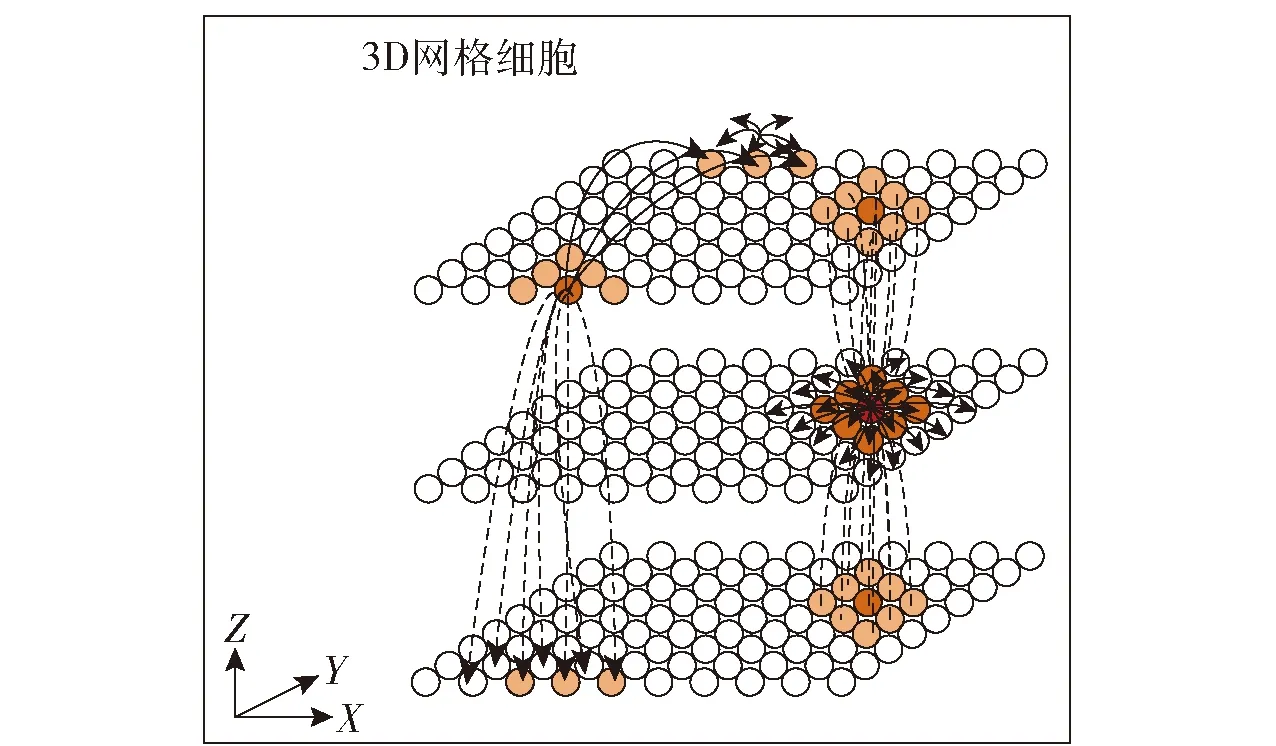
图11 三维连续吸引子[45]
2.2.2 振荡干涉模型
振荡干涉模型主要用于网格细胞模型的构建,模型启发来源于Koenig的研究报告。当大鼠探索环境时,鼠脑内嗅皮层- 海马区结构中脑电图表现 8~12 Hz的θ波振荡,当大鼠运动到位置细胞放电区域时,位置细胞出现θ波振荡,到达网格细胞放电野的位置时也出现θ波振荡,注入利多卡因使θ振荡减少后,内嗅皮层的网格状放电模式消失[48]。
单细胞的振荡干涉模型由Burgess等[49]首次提出,在该模型中,网格细胞的输入来自于头朝向细胞选择的最优方向。Zilli等[50]提出了多种不同网络作用的振荡干涉模型,该模型由可以保持恒定频率的基线振荡器V0和多个有源振荡器组成Vi,i≥1有源振荡器的相位相对于基线振荡器改变从而编码位置信息。不同的速度输入振荡器会改变振荡器的频率,从而产生高于或低于基线振荡器的频率,因此在有源振荡器与基线振荡器具有相同相位的位置集中产生一组平行带。振荡干涉模型如图12所示,模型中的输入为速度信号vi,震荡器Vi通过ωi进行内部耦合连接,网格细胞通过突触ωi,G接受震荡器的所有输出,经过整合后形成网格细胞六边形放电野。基于振荡干涉模型构建的网格细胞模型通过参数调节可形成不同尺度和相位的网格细胞放电野[51]。Yartsev等[52]的最近的研究表明,在果蝙蝠和猕猴的内嗅海马系统中并没有观测到θ波振荡,当对θ波振荡产生进行破坏,网格细胞放电野依然保持不变,振荡干涉模型在细胞建模中不具有广泛通用的生理学机制。
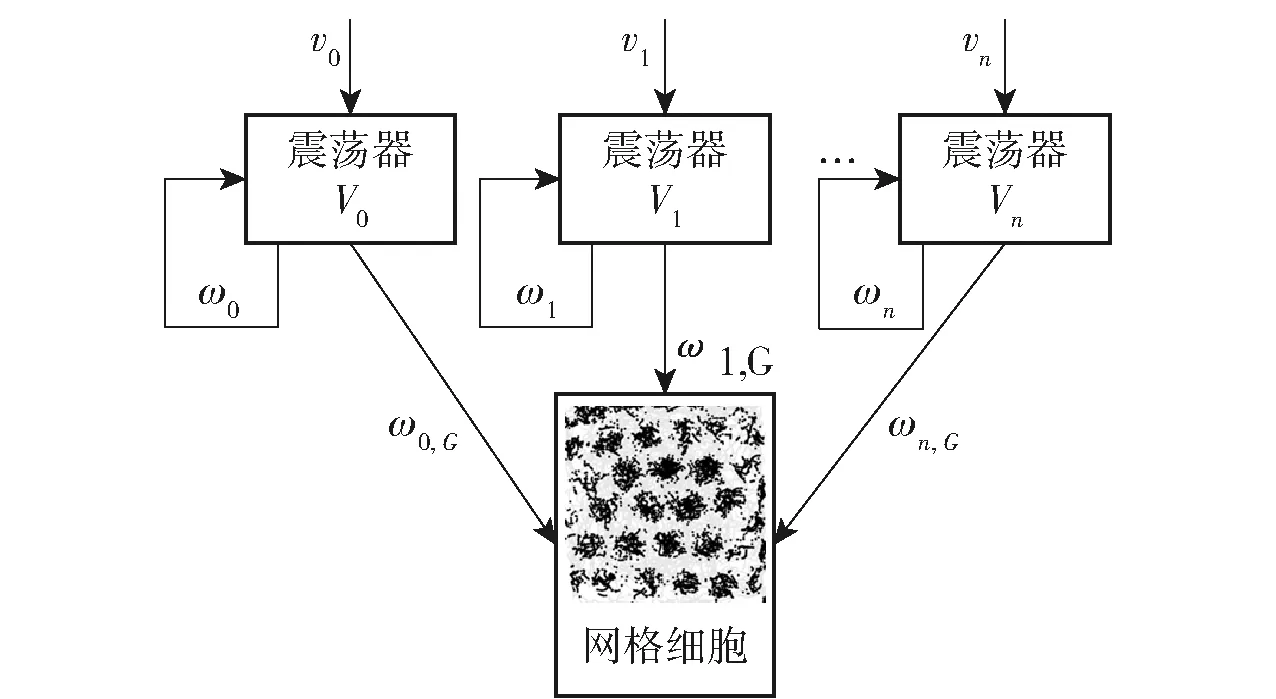
图12 网格细胞振荡干涉模型
2.2.3 脉冲神经网络模型
(1) 脉冲神经网络模型原理
脉冲神经网络(SNN)通过模拟大脑中神经活动进行建模,在生物神经元中,神经元之间的通信是通过传播动作电位序列(也称为传到下游神经元的脉冲序列)来完成的。突触前神经元的活动调节突触后神经元的膜电位,当膜电位超过阈值时产生动作电位或脉冲。脉冲产生的速率和脉冲序列的时间模式携带外部刺激有关信息并持续进行计算。这些单独的脉冲在时间上很稀疏,因此每个脉冲都有很高的信息含量,并且具有均匀的幅值(100 mV,脉冲宽度约为1 ms)[53]。
(2)脉冲神经网络模型类型和应用
脉冲神经网络通过脉冲的精确时间或脉冲序列进行通信[54],使单个脉冲的时间信息能够进行表达,脉冲神经元具有动态特性,即内部状态(定义为每个神经元输入的加权和)随时间变化[55-57],图13所示为前馈脉冲神经网络模型。

图13 前馈脉冲神经网络模型
生物神经元使用离散的脉冲来计算和传输信息,并且除了脉冲发放率外,脉冲时间也含有重要信息,由此脉冲神经网络更加贴近了生物神经元信息传递形式。脉冲神经网络中的信息是通过脉冲时序传达,包括延迟和脉冲速率,也可以通过群体进行信息表达,因此脉冲网络具有对生物神经系统中信息传输时间敏感的优势,这使脉冲神经网络在生物学上更具有生理学特性,对了解大脑的计算方式有很大研究价值。研究表明,每个脉冲的精确时序在神经编码中起着重要的作用[58]。精确的脉冲时序作为感觉信息处理区域[59]和大脑运动控制[60-61]的重要编码信息。
Hodgkin等根据乌贼轴突的细胞膜中离子通道的电压门控特性首次建立了脉冲神经网络模型—HH模型[62],该模型中描述了大量生物细节,计算量较大,随后人们提出了多种神经元模型,例如脉冲响应模型[63]、Izhikevich神经元模型[64]、以及带泄漏的积分触发(LIF)模型[65]。LIF模型直观描述了外部输入信息,具有计算简单等特点[66]。Lorenz等[67]将CA3中的位置细胞建模为LIF模型,该模型能有效对目标位置进行表征并形成放电野。Tang等[68]基于SNN模型构建多种空间表征细胞用于机器人导航控制系统。德国开姆尼茨工业大学Fred Hamker教授团队[67]基于SNN构建位置细胞序列模型建立基于地图的面向目标导航系统。
基于时序的脉冲序列含有大量有效信息可以减少能量消耗,可用于制造低能耗硬件设备。脉冲神经网络的学习通过调整突触权重实现,脉冲神经网络基于神经启发机制的学习规则并不能直接复制非脉冲形式,神经科学家定义这种学习规则及其变化规则为STDP(spike-timing-dependent plasticity)。通常STDP作为无监督学习机制,是一个时序非对称形式的Hebb学习法则,与突触前和突触后神经元峰值之间的时间间隔密切相关。如果突触前神经元在突触后神经元之前不久触发(大约10 ms),则连接它们的权重会增加。如果突触前神经元在突触后神经元后不久触发,则时间事件之间的因果关系是虚假的,权重会减弱。突触前脉冲在突触后脉冲前几毫秒内到达称为LTP (long-term potentiation),反之为LTD (long-term depression)。随着技术发展,神经科学家研究在脉冲神经网络下进行强化学习,特别提出调节奖励STDP (reward-modulated STDP, RM-STDP),由此,SNN可以在设计的全局奖励信息下得到一个最优决策[69],目前SNN模型在准确性方面还有一些不足,但性能随着研究的进步正在逐步提升。
3 面向未知复杂环境的类脑SLAM及路径规划技术应用
在未知复杂环境中实现自主导航需要在没有预设路线的前提下了解实际的环境信息,同步地图构建和定位(SLAM)[70]同时解决环境感知与定位这两个问题,是智能系统在未知复杂环境下进行自主导航的关键技术。传统的视觉SLAM在定位和制图精度上可以满足简单的应用场景需求,其搭载的视觉传感器在获取信息的过程中易受光照变化、环境图像纹理和快速运动等因素影响,以上影响因素对特征点的提取与匹配造成干扰,使其复杂环境的适用性较低。尽管研究人员开发的尺度不变特征变换[71](SIFT)算法使系统适应光照及图形形变带来的影响,提高了在复杂环境中的适用性,但效率较低,难以满足实时定位与建图需求。
通常情况下,环境感知信息存在噪声和误差,因此还需要进行优化处理,传统SLAM技术多数基于概率模型进行优化估计,其主要原理为:将当前采集到的环境数据通过数学模型处理传感器与环境的模糊性,其中较为常用的算法有卡尔曼滤波[72]算法、粒子滤波[73]算法、最大似然估计等。上述模型基于概率模型进行优化和估计,其准确性与可靠性依赖于建立精确的数学模型,而在未知复杂环境中难以构建精确数学模型,这使导航系统的适应性较差。
路径规划同样需要构建环境数学模型,随后利用搜索算法进行最优路径规划。常用环境建模方法主要有:栅格法、可视图法、维诺图法等,路径搜索算法有A*[74-75]算法、Dijkstra算法[76-77]、人工势场法[78-80]等。当面临复杂的大范围任务环境时,传统路径规划算法出现了搜索速度慢、寻优能力差等问题。仿生的群智能路径规划算法克服了传统路径规划算法的部分缺点,具有很好的鲁棒性,其路径规划速度也有所提升。现阶段仿生的群智能算法有蚁群算法[81-83]、遗传算法[84-86]、粒子群优化算法[87-88]等。仿生的群智能路径规划算法主要思想是将路径规划问题转化为最优搜索问题,然而仿生的群智能算法存在实时性较差,计算量大,全局搜索能力较弱等问题,其在路径搜索的快速性、可靠性方面依然需要进一步提升[89]。
经过生物学研究发现,哺乳动物具有构建认知地图和运动决策的能力,因此类脑SLAM和类脑路径规划具有解决实时性差、需要精确建模以及计算量大等问题。
3.1 类脑同步空间地图构建与定位
自然界中的生物不具备高精度传感器也可以在长途复杂环境中寻找食物并在找到食物后以捷径回巢[90],同时在缺乏部分信息源时也能依靠经验信息进行导航。1948年,Tolman等[91]发现在迷宫实验中的老鼠能够迅速地找到通往食物的捷径,并且当熟悉路径被阻挡后能够找到新的路径,由此猜想,哺乳动物通过在大脑中构建地图来进行空间认知与导航。
Tolman[92]提出了认知地图的概念,认知地图是通过各种形式对环境进行的智力描述,动物对外界环境的学习,并不是把起始点到目标点的路径信息进行简单的搜集,而是对外界环境生成一种内部的抽象描述。认知地图是动物对环境信息进行收集、组织、贮存、回忆、并对其空间方位和特征属性加以编码的产物。其建立在直觉和本能的基础上进行编码、解码、学习和理解。
在未知复杂环境中进行导航时,先在认知地图中锁定目标的大致的范围,然后凭借对目标形象的理解锁定小范围内的目标。认知地图的构建主要依靠于海马体各区域内的空间表征细胞,通过对外源信息和内源信息进行整合以及空间表征细胞之间的相互作用最终形成认知地图,实现外部环境在大脑内的神经重现。研究学者受生物导航能力启发,研究基于生物大脑导航机理启发的SLAM系统,进而提升整体地图构建与定位性能,环境地图构建的主要形式有度量地图、拓扑地图及混合地图[93]。
3.1.1 基于单一种类细胞模型的地图构建
如前文所述,位置细胞和网格细胞具有表征位置信息能力,因此构建单一种类空间表征细胞模型可以得到路径位置信息和认知地图,同时也为形成更加完善的类脑自主导航系统提供基础。Jacobson等[94]提出基于位置细胞模型的多尺度建图系统,利用位置细胞模型将相机、WiFi和气压高度计信息进行编码,由不同尺度位置细胞构建路径地图。南洋理工大学Datta等[95]研究了一种在线快速学习的地图构建技术,该技术利用视觉信息作为位置细胞模型输入,通过位置细胞模型解算得出实时位置信息并构建路径地图。Barboni教授研究团队[96]基于网格细胞脉冲放电形式构建模型并用于机器人平台验证,该机器人具有大鼠自由探索环境的行为特征并向中心系统报告位置信息,从而构成生物启发的认知地图,具有一定的智能行为。
3.1.2 基于多种类细胞模型联合的地图构建
随着对单一种类细胞模型的有效研究,逐渐形成基于多种空间表征细胞模型的类脑SLAM系统。通过不同种类空间表征细胞模型之间的信息交互,其系统表现出更加卓越的性能。美国罗格斯大学的Tang等[97]利用大脑导航机理构建一个实时SLAM系统,该系统由位置细胞、网格细胞、头部方向细胞和边界细胞组成。该系统基于SNN网络构建空间表征细胞模型,在SNN中利用胜者为王学习规则构造位置细胞模型,引入异种突触竞争学习塑造网格细胞和位置细胞之间的突触连接。同时将系统集成到Loihi芯片中,该系统在未知环境下构建地图的实时性是传统方法的12倍。
昆士兰科技大学Ball教授团队[98]对基于啮齿动物大脑中导航神经计算机理的RatSLAM系统进行建模,该系统将纯视觉信息作为输入,用连续吸引子网络来建立细胞模型,利用经验来建立未知定位可能假设,使其在感知信息缺失的情况下也能正常工作,通过跑车试验验证了系统的可靠性。在前面的研究基础上,Milford教授团队继续研究,提出基于三维网格细胞和多层头朝向细胞吸引子神经网络模型的四自由度SLAM系统,如图14所示。系统模拟哺乳动物利用视觉对环境的感知,扩展网格细胞与头朝向细胞维度,利用视觉里程计数据激发三维网格细胞和多层头朝向细胞活性,将采集的视觉信息与预先储存的视觉模板进行匹配,当处于熟悉场景时局部场景细胞被激活,将活性输入到头朝向细胞和网格细胞模型中进行校准,通过路径积分形成拓扑地图,实现对三维空间的认知以及通过熟悉场景标定所在位置的功能[45]。
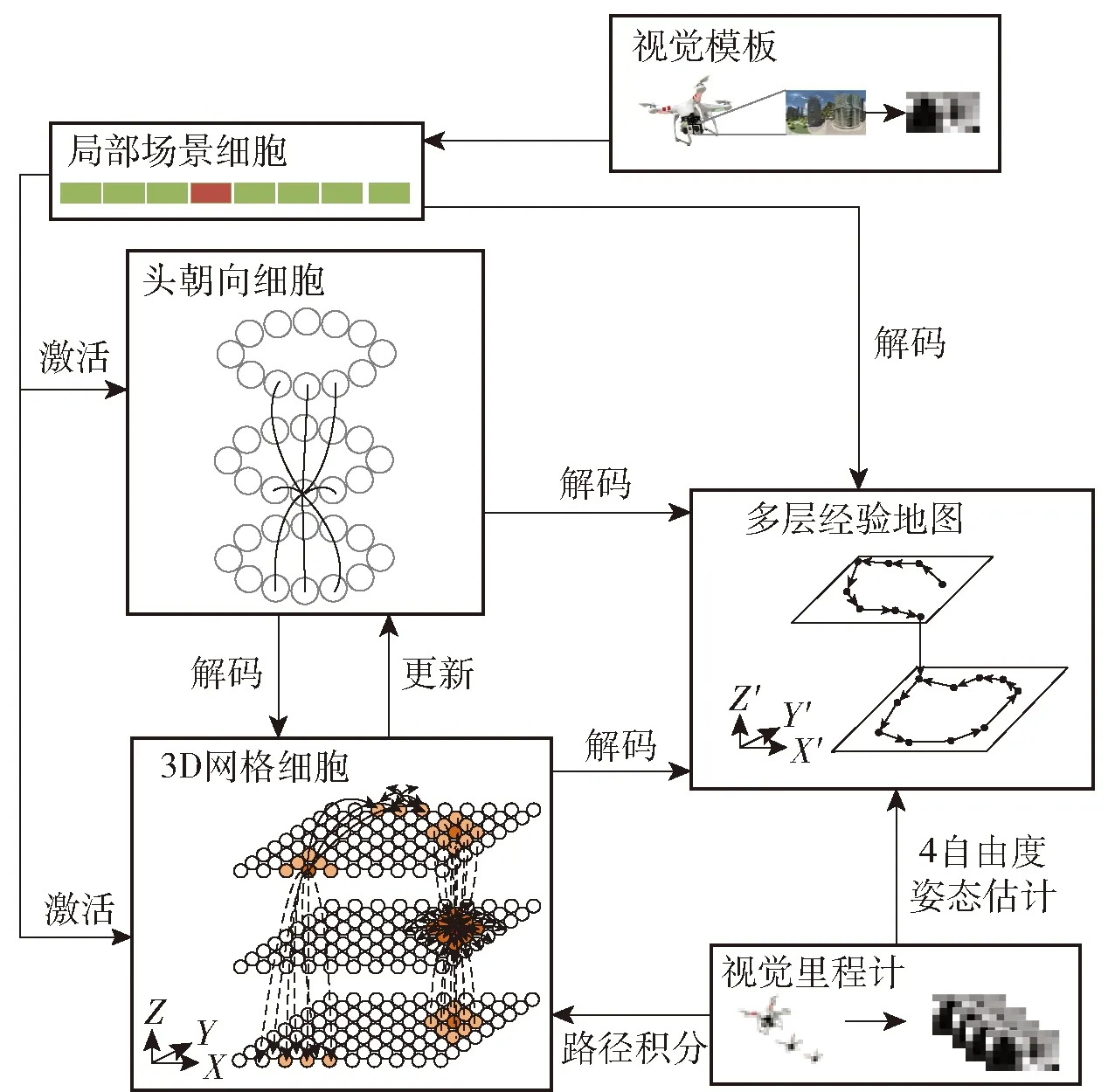
图14 三维四自由度SLAM[45]
复旦大学Zeng等[99]提出基于空间记忆系统的视觉信息和自运动信息融合的贝叶斯吸引子网络模型的SLAM系统,如图15所示。该网络模型由两个子网络模型组成,分别为头朝向细胞网络和网格细胞网络,子网络分别对应哺乳动物大脑头朝向细胞和网格细胞的神经编码形式,其中头朝向细胞用于角度编码,网格细胞用于位置信息编码,分别表示方向和位置信息。将来自前庭的自运动信息输入到每个子网络中,通过细胞群编码进行整合以及视觉信息进行校正,并基于贝叶斯方法进行更新,成功构建半度量半拓扑地图,该系统在不同环境特征条件下均具有自主性。
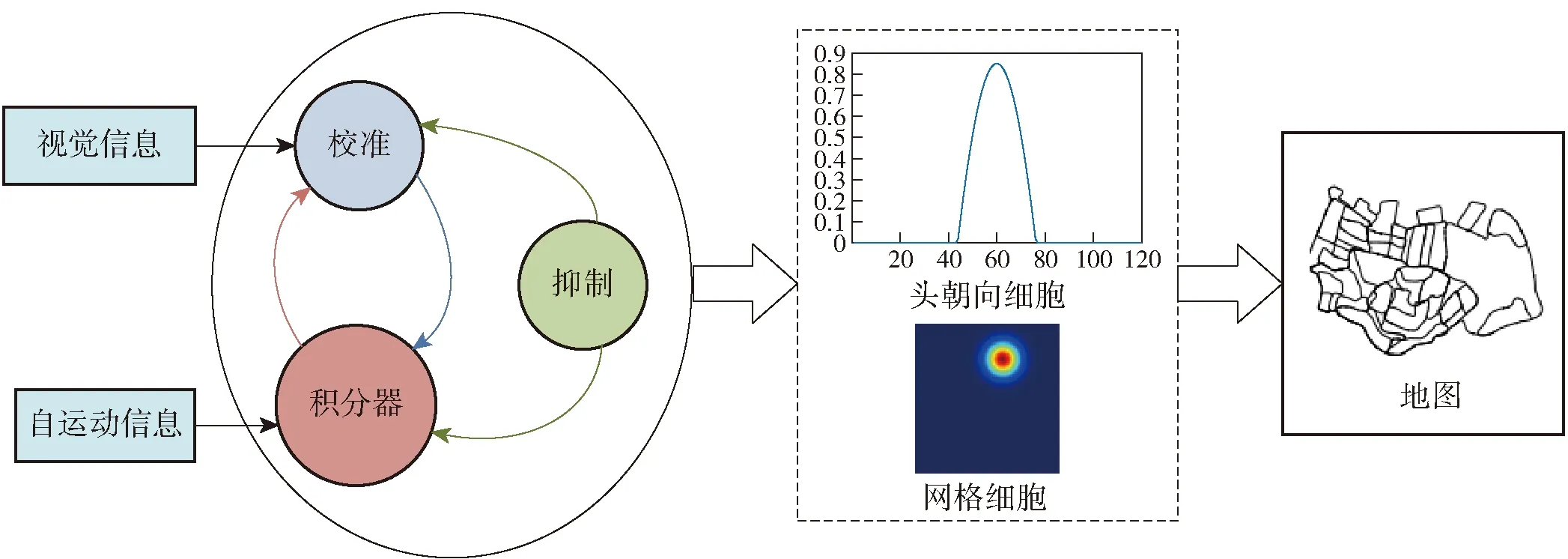
图15 基于贝叶斯吸引子网络SLAM系统[99]
3.2 类脑路径规划
路径规划作为自主导航技术中的另一核心技术,需考虑路径形成过程中的多方面因素,如路径行程距离、避免与障碍物碰撞以及任务执行效率等。随着人工智能技术的飞速发展,路径规划也应用更加智能的技术。无人移动平台通过推断任务目标和位置之间矢量信息产生路径,该路径的产生具有不确定性,随后可由类脑算法选择捷径行走到任务目标区域。
3.2.1 基于单一种类细胞模型的类脑路径规划
由于SNN具有更好的神经放电形态模拟特性,中国、美国和日本等国家的科研单位和高校对基于SNN的空间表征细胞模型在路径规划和导航控制应用方面具有了一些研究进展和成果。日本东京都立大学Naghizadeh等[100]开发基于SNN的动态在线路径规划模型,该模型模拟神经元(某种空间表征细胞)信息传递方式,具有较好的规避障碍能力,并在不可预测的旅行成本下应用。该模型可用于机器人在崎岖地形上的路径规划,系统设计如图16所示[100]。第1步,使用Kinect传感器生成地图的结构,并通过无线通信将其发送给所提出的系统。第2和第3步,执行规划的路径,前向传递构造神经元连接以寻找可能的路径,后向传递用突触修剪模型可减少获得当前位置到目标位置最佳路径的神经网络中的无效神经元。为了应对不可预测的障碍物,执行有效的动态规划,其处理机制为起始点神经元首先向相邻神经元发出信号,计算突触权重值。当信号触发目标神经元后,传输过程停止,然后优化突触数目和处理反向信号。第4步,通过蓝牙通信将最佳路径发送给机器人,进行第5步执行命令产生相应动作,最后,机器人移动并更新当前位置,该方法令机器人在未知环境中更加高效地规划移动路径并灵活躲避障碍物。
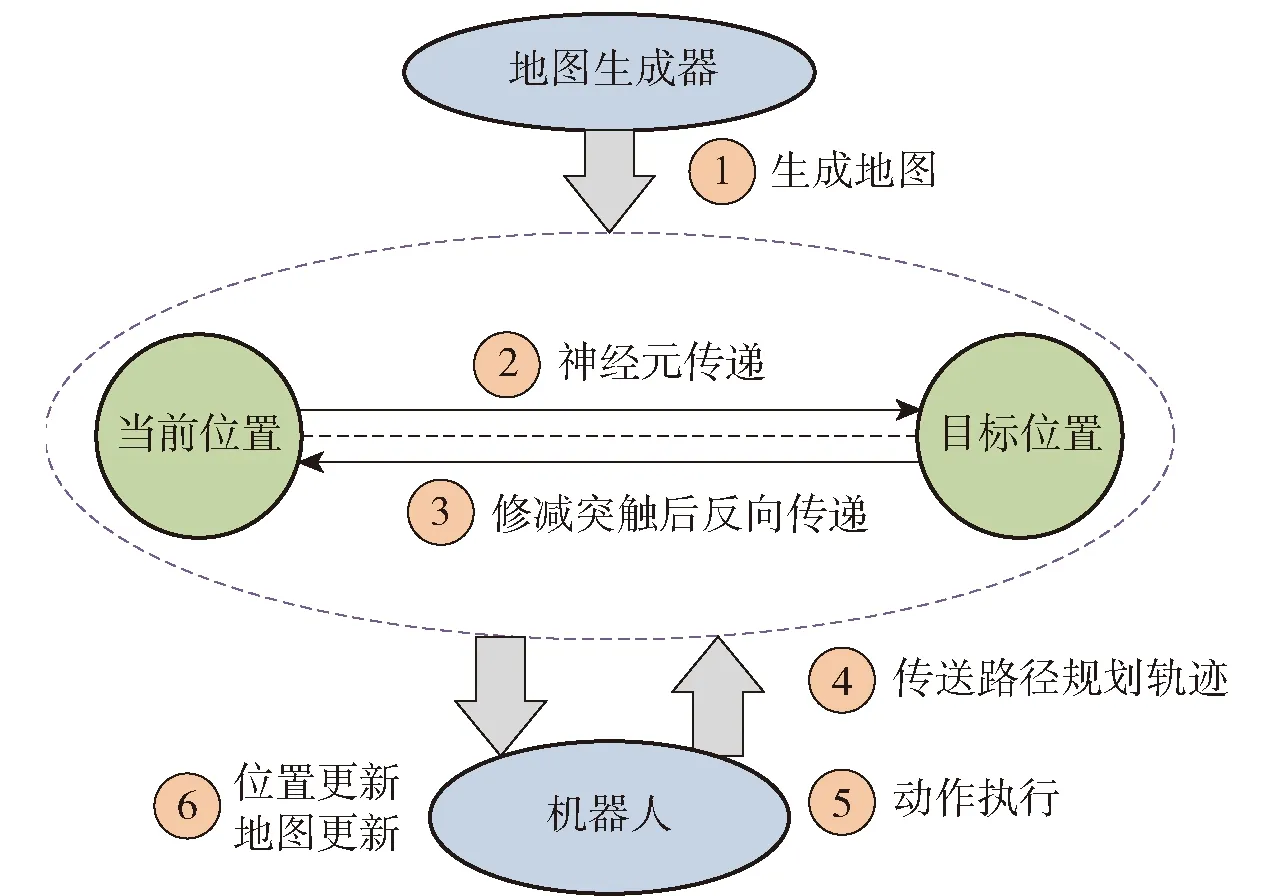
图16 基于神经元活动在线动态路径规划模型[100]
美国Naghizadeh教授研究团队[101]提出一种网格细胞导航模型,该计算模型通过调整细胞之间的权重构建导航决策过程,提供机器人在未知环境以及未知目标位置环境下通用的路径搜索方案,减少了访问先验状态随机游走的可能性,可在合理速度情况下实现相对更小能量消耗的有效路径规划,通过与相近的方法进行比较,该方法具有更高效的速度处理能力,同时运算占据更小的内存。
在已知环境活动的啮齿动物能够在观察到目标位置后,快速规划面向目标导向的轨迹。大脑中CA3区在面向目标导航过程中起到关键作用,其主要特征表现为位置细胞对特定环境的放电行为。由此,如何通过构建位置细胞模型模拟生物在熟悉环境中的面向目标快速导航行为成为研究重点。Dane等[102]将位置细胞建模为吸引子神经网络模型,将障碍物位置、墙体位置等空间环境编码生成序列,之后利用马尔可夫决策过程对位置序列按照离目标距离较近进行较大奖励,进而实现空间任意位置到达目标位置的路线规划。德国Lorenz教授研究团队[67]提出了一个位置细胞序列模型,将大脑DG中的神经细胞和CA3中的位置细胞建模为LIF模型的脉冲神经网络,通过基于奖励机制的目标学习过程生成位置细胞表征的位置序列,并且在形成的位置序列中指导空间活动行为,该模型能够模拟大鼠在决策任务中的行为。上述两种研究中的位置细胞模型虽然不同,但中心思想都是通过构建位置序列并给予目标位置最大奖励产生最佳决策和面向目标的导航路径。
3.2.2 基于多种类细胞模型联合的类脑路径规划
位置细胞模型作为类脑导航系统中的重要模块,能够模拟生物导航过程中的某些方式,但对于类脑导航系统依然存在很多的性能提升空间,由此研究学者开展进一步研究,细化空间表征细胞的功能,增强模型单元之间的信息交互能力,在未知环境中路径规划、避障等展现更优越的性能,同时与构建类脑认知地图相结合,在路径规划的同时对环境形成一定的认知。Dolle等[32]提出由位置细胞、标定细胞、动作细胞等空间表征细胞模型组成的系统结构,该计算模型具有过程线索导航和路径规划转换机制,动作细胞能够在基于程序性或者认知性这两种学习机制下进行导航策略选择,同样可以在环境变化的条件下选择最优行动路径。Cuperlier等[103]依赖海马体解剖结构之间的相互作用机理构建导航和路径规划模型。通过位置细胞和CA之间的联系机理构建过渡细胞模型,过渡细胞模型能够减少路径规划中对位置细胞的依赖,可以通过学习后建立分段认知地图,之后利用Bellman-Ford algorithm算法进行路径选择。美国罗格斯大学Tang等[68]基于SNN构建由网格细胞、头朝向细胞、位置细胞、边界细胞等多种空间表征细胞模型组成的导航控制系统,其构建的系统模型在“Gridbot”机器人中进行验证,实验表明该神经网络能使“Gridbot”机器人在最少的学习条件下就可以自己绘制环境地图并学习不同目标的位置,可以像灵长类动物那样,对环境进行有效的探索和反应。Huang等[104]同样基于SNN构建网格细胞模型,该模型能够在视觉消失的情况下,在短时间内保持对空间的方向定位,同时在模型中加入一个简单的决策网络控制平台使机器人能够执行觅食和规避风险等基本任务。
在认知地图的基础上执行面向目标导航、探索最优路径等任务能更加有效地认知未知环境并提升路径规划效率。Erdem等[51]提出了一种基于头向细胞、网格细胞、位置细胞和额前皮质(PFC)细胞等空间表征细胞模型组成的面向目标导航系统。在未知环境下,系统模拟大鼠随机探索环境过程,逐步创建由位置细胞和PFC细胞组成的地图。在模拟的大鼠探索完成后,系统对位置可形成记忆并快速筛选候选方向进行轨迹的前向线性预先探测,将激活PFC细胞奖励最大的方向作为下一动作的运动方向。日本Chin等[105]基于大脑细胞神经活动提出了动态的路径规划模型,在模型中利用贝叶斯自适应共振联想记忆算法压缩数据提高拓扑地图的生成效率,同时在拓扑地图的生成过程中不需要有关环境的先验知识,此过程可以对未知环境有效进行探索;路径规划器根据拓扑地图信息生成最优路径,此外还可以从最优路径中减除无效节点,进一步提高最优路径生成效率。Shim等[106]提出了方向驱动的类脑导航系统,该系统由认知地图构建、基于网格化的方向规划和多层不对称局部导航3部分组成,通过在认知地图中提取方向,能更快速并有效地从给定目标方向移动到目标所在位置。
我国在类脑路径规划方面同样进行了研究,并在机器人等平台上进行了结果验证。北京科技大学于乃功教授团队[107]基于吸引子神经网络构建类脑SLAM认知地图,在所构建精准的认知地图基础上采用全局导航和局部导航算法实现机器人的导航。全局导航通过构建的认知地图所呈现的最近路线面向目标进行导航,局部导航算法对多组可能路线进行评估之后,选择最优轨迹对应的速度驱动机器人运动。机器人在全局导航形式下向目标点移动,同时当前的最优路径由局部导航算法进行选择,在面向目标进行导航的过程中机器人可以避开障碍物规划路径,并在一定范围内返回到先前的导航路径。
4 展望
随着脑科学的快速发展,导航、制导与控制和神经科学这两个学科之间也在快速地交融发展。虽然在类脑导航模型、类脑SLAM以及类脑路径规划等研究方向有了一定的研究成果,但实现较强的环境认知和适应能力,提高在空间环境中的导航精度还需要开展许多的研究工作。类脑自主导航技术的研究重点和发展方向有以下几个方面:
1)在类脑理论研究层面,探究空间表征细胞在智能导航行为过程中的协作机理。
生物大脑中海马区与内嗅皮层之间构成的信息传递通路为机器人、无人机等无人移动平台提供环境认知、情景学习与自主导航的理论方法。大脑对未知复杂环境进行认知与记忆是一个复杂的细胞放电活动,空间表征细胞放电活动与多种感官信息相关,但目前对神经信息传递机理的理解并不完善,需要随大脑机理的深入研究而升级为更加智能的导航系统。
2)研究空间表征细胞交互连接方法,为类脑导航定位、地图构建、路径规划技术研究提供更加完善的类脑导航系统。
海马体中参与环境认知与导航的空间表征细胞主要为位置细胞、头朝向细胞和网格细胞。在环境的探索过程中,空间表征细胞通过学习改变相互之间的突触连接,从而形成特定的环境认知记忆回路。为了能更好地学习环境,需要寻找最优突触训练方法,增强空间表征细胞的感知和相互作用能力,因此应继续探索与完善空间表征细胞交互连接模型,为后续SLAM、路径规划及运动控制技术研究提供模型基础。
3)研究多源感知信息融合的类脑自主导航技术。
构建认知地图作为动物对未知环境进行认知的重要途径。在构建认知地图的过程中,视觉是重要的信息来源之一,成为人等哺乳动物感知和认知环境的重要方式,在黑暗或弱光环境中,动物通过多种感官信息以及空间表征细胞的相互作用探索环境,具有较好的环境适应能力以及导航能力。因此结合传统导航信息融合方法,开展视觉、惯性导航和卫星导航等多源信息融合的类脑导航系统研究是目前的热点研究方向。
4)研究类脑认知、导航和决策一体化的类脑导航系统。
类脑路径规划/运动决策基于大脑伏隔核中的动作细胞、多巴胺神经元的预测奖励机制,通过已经构建的认知地图确定前进的方向和速度。现阶段通过构建动作细胞模型模拟大脑路径规划方向的研究还需要进一步完善,该方向的研究为未来形成类脑认知、导航和决策一体化的导航系统奠定了基础。
5) 研究面向三维空间环境的六自由度导航信息输出的类脑导航系统。
类脑自主导航的研究还处于发展阶段,基于神经科学的三维空间信息整合机理尚未统一,由此在“类脑”理论层面上缺少经过生物学实验验证的三维模型参考。在类脑自主导航的工程应用方面,目前多以机器人为应用研究对象,已有的研究成果主要为平面空间面向目标的矢量导航研究。无人移动平台在进行环境探索或者面向目标进行导航过程中所构建的地图也多为二维平面地图,从模型中获得的导航参数为二维位置信息和方向信息,缺少俯仰角、滚转角等导航参数,因此在对无人机进行姿态控制的过程中会存在一定的局限,亟需进行三维空间类脑导航技术研究。
5 结论
本文沿着类脑自主导航技术的研究路线进行了总体阐述,归纳了内嗅皮层- 海马区中的传递方式以及空间表征细胞模型的放电特性;分析了基于内嗅皮层- 海马区信息传递通路以及类脑导航系统结构的模块组成和作用,分类总结通过模拟神经元放电形态和空间表征细胞放电特性构建空间表征细胞模型的方法;重点分析了基于空间表征细胞模型在类脑导航系统中的作用以及类脑SLAM和类脑路径规划技术的发展和应用。最后,本文提出类脑自主导航技术未来的研究展望。
通过分析现阶段国内外开展的类脑自主导航研究的现状,类脑自主导航技术有着广阔的发展和应用前景,脑科学的发展与类脑自主导航技术的发展互为辅助,生物大脑作为一个复杂的中枢控制系统,目前对大脑的研究还不够全面,这也是制约类脑自主导航技术理论和模型构建的主要因素之一,与此同时,类脑算法也需要性能提升。另外。类脑芯片的研发与类脑自主导航技术的工程实现息息相关,随着脑科学和类脑芯片的快速发展,类脑自主导航系统相关研究技术有望取得更大的突破。
猜你喜欢
科学24小时(2021年1期)2020-12-24
中国惯性技术学报(2020年2期)2020-07-24
电子制作(2019年19期)2019-11-23
数学年刊A辑(中文版)(2019年3期)2019-10-08
作文新天地(初中版)(2019年6期)2019-08-15
电子制作(2019年24期)2019-02-23
北京航空航天大学学报(2017年6期)2017-11-23
中小学信息技术教育(2017年6期)2017-06-23
浙江大学学报(工学版)(2016年10期)2016-06-05
重型机械(2016年1期)2016-03-01
